牦牛小腦不同區域神經營養素-4及其受體的表達特征與定位研究
2024-04-17 01:00:38劉珊珊杜曉華劉霞吳亞娟鄭麗平
核農學報 2024年5期
關鍵詞:區域
劉珊珊 杜曉華,* 劉霞 吳亞娟 鄭麗平
(1甘肅農業大學動物醫學院,甘肅 蘭州 730070;2甘肅農業大學生命科學技術學院,甘肅 蘭州 730070)
牦牛是青藏高原特有畜種和主要的畜禽遺傳資源,是世界上唯一能夠在高寒低氧、強紫外線、牧草營養短缺等極端環境中生存的大型反芻動物[1-2]。由于牦牛具有與低氧適應相關的解剖結構和生理特征,如較大的心和肺、較厚的體表覆蓋物和發達的皮膚腺,使其能夠適應高寒低氧環境[1-3]。腦作為耗氧量較大的組織器官,在高原地區對氧分壓的變化十分敏感[4-5]。小腦是中樞神經系統運動協調的主要中心,與大腦運動皮層保持著廣泛的神經連接,盡管其體積僅為大腦總體積的10%,但其中包含了超過50%的神經元[6-8]。缺氧可導致神經元供能不足,引發多種異常生理活動,包括酶活性降低、細胞膜通透性改變、膜電位異常和神經沖動傳導障礙,最終引起一系列高原病[9-11]。
神經營養素-4(neurotrophin-4,NT-4)是神經營養因子家族重要的成員,也是中樞神經系統神經元的存活因子[12-13],在促進神經元生長、發育、分化與成熟,維持神經元存活以及促進神經元損傷后修復與再生及神經-免疫-內分泌等方面均發揮著重要作用[14-15]。神經營養性酪氨酸激酶受體2(neurotrophic tyrosine kinase receptor type 2,NTRK2)是一種酪氨酸激酶受體,在神經系統中發揮重要的生物學功能[16-17]。研究發現,NT-4可促進腦缺血缺氧后血管的增殖來抵抗缺血缺氧性腦損傷,當NT-4與反應神經元表面受體NTRK2結合后,觸發受體激活,從而啟動復雜的細胞通路,并促進神經元的生存和發育,抑制細胞凋亡[18-19]。同時,NT-4通過表達上調激活受體NTRK2,組裝為NT-4-NTRK2復合物,能夠啟動細胞內一系列反應來調節缺氧誘導因子-1α(hypoxia-inducibe factor-1α,HIF-1α)的表達,進一步調節參與糖代謝和血管生成的下游蛋白的表達,進而提高動物機體對低氧環境的耐受性[20-21]。此外,機體長期處于高原低氧環境下會導致活性氧(reactive oxygen species,ROS)蓄積,腦組織因ROS過量堆積而引起炎癥反應[22-23],星形膠質細胞和小膠質細胞在腦內炎癥反應中明顯激活,釋放多種炎癥因子,并損傷腦組織及血管內皮功能[24]。NT-4也可通過降低促炎因子的表達、減輕神經炎癥以及改善神經功能對缺血缺氧性腦損傷產生保護作用[25]。目前對NT-4及其受體NTRK2在神經系統方面的研究主要集中在小鼠、大鼠、猴等物種的神經損傷修復等領域,關于牦牛的研究尚鮮見報道。
鑒于此,本研究以牦牛和黃牛小腦組織為研究對象,通過蘇木精-伊紅染色(hematoxylin-eosin,HE)、免疫組織化學(immunohistochemistry,IHC)、實時熒光定量PCR(quantitative real-time PCR,qRT-PCR)、蛋白免疫印跡(Western blot,WB)對NT-4和NTRK2的分布及表達特征進行研究,旨在闡明NT-4及其受體NTRK2在牦牛與黃牛小腦各區域中的表達規律,分析與牦牛腦組織適應低氧環境的關系,以期為研究牦牛腦組織低氧適應性機制提供參考。
1 材料與方法
1.1 樣品采集
甘南牦牛(母,6歲)樣品在甘肅省甘南藏族自治州合作市采集,南陽黃牛(母,6歲)在河南省鄭州市采集,各3頭。分別采集不同區域小腦組織,置于液氮和4%多聚甲醛溶液中保存備用。
1.2 主要試劑及儀器
TriQuick RNA提取試劑盒、放射免疫沉淀試驗(radioimmunoprecipitation assay,RIPA)組織快速裂解液,科寶(蘭州)生物科技有限公司;Evo M-MLV反轉錄試劑盒,(長沙)艾科瑞生物工程有限公司;NT-4多克隆抗體(rabbit anti-NT-4 polyclonal antibody,DF6226),艾菲(美國)生物科技有限公司;神經營養性酪氨酸激酶受體2(rabbit anti-NTRK2 monoclonal antibody,4603),上海優寧維生物公司;兔抗β-actin多克隆抗體(rabbit anti-β-actin polyclonal antibody,bs-0061R)、山羊抗兔IgG/HRP (goat anti-rabbit IgG/HRP,bs-0295GHRP),博奧森(北京)生物技術有限公司;鏈霉卵白素-生物素法檢測(streptavidin peroxidase,SP)試劑盒,中杉金橋(北京)生物技術有限公司;增強型HRP-DAB底物顯色試劑盒,賽維爾(武漢)科技有限公司;LightCycler96熒光定量PCR儀,瑞士Roche公司;冷凍型高通量組織研磨儀,寧波新芝生物公司;臺式高速冷凍離心機,長沙湘儀離心機儀器有限公司;TGrade加熱型程控五段金屬浴,北京天根生化公司;超微量分光光度計,北京凱奧科技發展有限公司。
1.3 實時熒光定量PCR(qRT-PCR)
1.3.1 引物設計 根據NCBI網站GenBank數據庫(https://www.ncbi.nlm.nih.gov/genbank)公布的牦牛NT-4(NM-001304828)與NTRK2(NM-010808129.3)基因序列,同時以β-actin(NM-173979.3)為內參基因,使用Primer Premier 6.0軟件設計引物,并送至上海生物工程股份有限公司進行合成,引物信息見表1。
1.3.2 總RNA提取和反轉錄 用研磨儀充分研磨牦牛與黃牛小腦各區域組織樣品,使用TriQuick RNA提取試劑盒提取組織總RNA,并用分光光度計測定RNA濃度,按照Evo M-MLV反轉錄試劑盒說明書將RNA反轉錄為cDNA,置于-20 ℃冰箱保存備用。
1.3.3 牦牛與黃牛小腦不同區域NT-4及NTRK2基因擴增 采用反轉錄的cDNA作為模板,對NT-4、NTRK2、β-actin進行PCR擴增,并使用10 g·L-1瓊脂糖凝膠對擴增產物進行分析,以確定目標片段的大小和純度。
1.3.4 qRT-PCR檢測NT-4及NTRK2在牦牛與黃牛小腦中的表達 以β-actin為內參基因,使用熒光定量PCR儀分別將牦牛與黃牛小腦不同區域NT-4及NTRK2進行擴增反應,每組3個重復。反應體系為20 μL:SYBR High-Sensitivy q-PCR SuperMix 10 μL,上下游引物各0.8 μL,cDNA模板 1 μL,dd H2O 7.4 μL。反應條件:50 ℃預熱2 min,95 ℃預變性40 s;95 ℃變性30 s,60 ℃退火30 s,共40個循環;每個樣品設4次重復。反應結束后,使用Light Cycler?96軟件采集Ct值,利用2-ΔΔct法計算目的基因的相對表達量。
1.4 蛋白免疫印跡(WB)
1.4.1 蛋白樣品制備 稱取牦牛與黃牛小腦不同組織各0.2 g于無酶離心管中,分別加入2 mL RIPA裂解液和20 μL蛋白酶抑制劑,置于預冷的-10 ℃研磨儀中研磨,然后搖床冰浴2 h,離心后收集上清,蛋白濃度測定后分裝于-80 ℃冰箱保存備用。
1.4.2 WB檢測 將提取的蛋白與4×蛋白上樣緩沖液按3∶1的比例混勻后,98 ℃恒溫金屬浴進行蛋白變性10 min,冷卻至室溫,存于-20 ℃備用。變性蛋白經10%聚丙烯酰胺凝膠電泳后,濕轉法轉印至0.22 μm聚偏二氟乙烯膜(polyvinylidenefluoride,PVDF)上,用50 g·L-1脫脂奶粉室溫封閉4 h后,孵育一抗(NT-4:1∶500,NTRK2:1∶1 000,β-actin:1∶2 000稀釋)4 ℃過夜,磷酸鹽緩沖液+Tween(phosphate buffer solution with Tween,PBST)洗滌4次,每次15 min,室溫搖床孵育二抗(1∶3 000)2 h,PBST洗滌4次,每次15 min,滴加化學發光液(enhanced-chemiluminescence,ECL)置于化學發光儀進行顯影。
1.5 免疫組織化學(IHC)檢測
石蠟切片經二甲苯、苯酒(無水酒精與二甲苯1∶1配制)和梯度酒精脫蠟脫水,然后用檸檬酸鹽緩沖液進行抗原熱修復,冷卻至室溫后,按SP試劑盒說明書依次滴加內源性過氧化物阻斷劑和山羊血清工作液進行阻斷封閉,隨后滴加一抗,其中NT-4和NTRK2稀釋比例均為1∶100,陰性對照用磷酸鹽緩沖液(phosphate buffer solution,PBS)代替,4 ℃過夜孵育,次日用PBS洗滌15 min后,滴加生物素標記山羊抗兔IgG于37 ℃溫箱孵育15 min,PBS洗滌后滴加辣根酶標記鏈酶卵白素工作液于37 ℃溫箱中孵育15 min,PBS洗滌15 min后滴加現配的增強型二氨基聯苯胺(diaminobenzidine,DAB)進行顯色,在光學顯微鏡下控制染色并用自來水終止顯色,依次用蘇木精復染、鹽酸酒精分化、自來水返藍,脫水透明后用中性樹膠封片,在光學顯微鏡下觀察并拍照。
1.6 數據統計分析
使用SPSS 23.0軟件進行單因素方差分析(analysis of variance,ANOVA),所有數據均以平均數±標準差(Mean±SD)表示,并采用Graphpad Prism 8.0進行繪圖。
2 結果與分析
2.1 qRT-PCR檢測結果
PCR擴增產物通過10 g·L-1瓊脂糖凝膠電泳后,片段大小與預期相符。NT-4(圖1-A)、NTRK2(圖1-B)和β-actin(圖1-C)分別為160、105和158 bp,擴增產物條帶單一,以上結果說明cDNA模板可用于qRTPCR檢測。
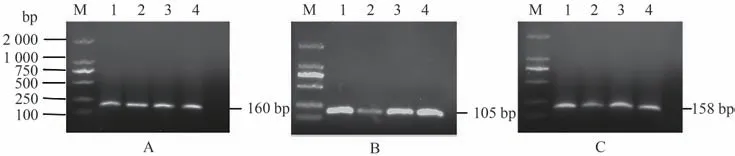
圖1 NT-4 (A)、NTRK2 (B)和β-actin (C)基因擴增Fig.1 NT-4 (A),NTRK2 (B) and β-actin (C) gene amplification
qRT-PCR檢測結果表明,牦牛與黃牛小腦各區域中均有NT-4基因的表達,其在牦牛小腦半球皮質中表達量最高,且顯著高于其他區域(P<0.05,圖2-A),蚓部髓質中表達量最低。另外,黃牛NT-4基因的相對表達量在蚓部髓質中最高,與其余各區域相比,具有顯著差異(P<0.05,圖2-B),小腦半球髓質表達量最低。
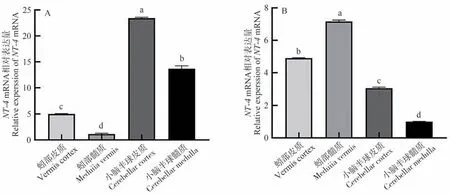
圖2 NT-4 基因在牦牛(A)與黃牛(B)小腦不同區域的表達結果Fig.2 The expression of NT-4 gene in different regions of the yaks (A) and cattles (B) cerebellum
NTRK2基因在牦牛與黃牛小腦各區均有不同程度的表達。其中,NTRK2基因在牦牛蚓部皮質中表達量最高,且顯著高于其他組織(P<0.05,圖3-A),小腦半球皮質次之,小腦半球髓質表達量最低。而在黃牛小腦半球髓質表達量最高,且顯著高于其他組織(P<0.05,圖3-B),蚓部皮質次之,小腦半球皮質表達量最低。
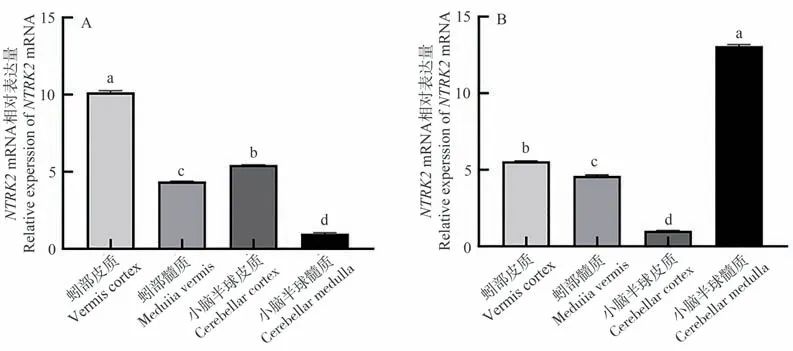
圖3 NTRK2基因在牦牛(A)與黃牛(B)小腦不同區域的表達結果Fig.3 The expression of NTRK2 mRNA in different regions of the yaks (A) and cattles (B) cerebellum
與黃牛相比,NT-4基因在小腦半球皮質和小腦半球髓質中的表達量顯著高于黃牛,而在蚓部髓質中低于黃牛(圖4-A);NTRK2基因在牦牛小腦髓質中的表達量顯著低于黃牛,而在蚓部皮質和小腦半球皮質的表達量顯著高于黃牛,蚓部髓質的表達量與黃牛無明顯差異(圖4-B)。
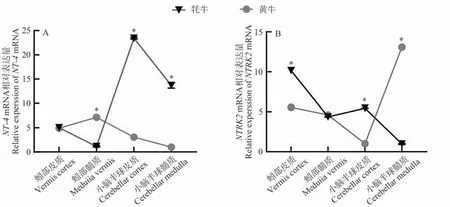
圖4 牦牛與黃牛小腦不同區域NT-4及NTRK2基因表達比較Fig.4 Comparative expression of NT-4 and NTRK2 in different regions of the yaks and cattles cerebellum
2.2 WB檢測
WB結果表明,NT-4蛋白在牦牛與黃牛小腦各區域均有表達。牦牛小腦半球皮質表達量最高,且顯著高于其他組織(P<0.05,圖5-A),小腦半球髓質次之,蚓部皮質表達量最低。黃牛蚓部髓質表達量最高,顯著高于其他組織(P<0.05,圖5-B),蚓部皮質次之,小腦半球髓質表達量最低。
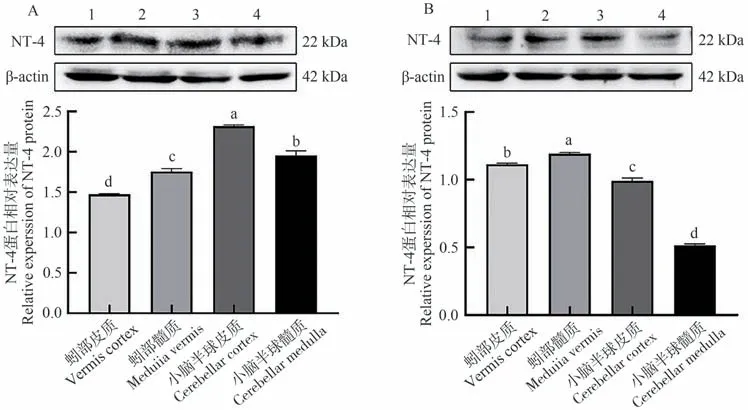
圖5 牦牛(A)與黃牛(B)小腦不同區域中NT-4蛋白表達情況Fig.5 Expression of NT-4 protein in different regions of the yaks (A) and cattles (B) cerebellum
NTRK2蛋白在牦牛與黃牛小腦各區域均有表達,其中,NTRK2在牦牛蚓部皮質中表達最高,且顯著高于其他組織(P<0.05,圖6-A),小腦半球髓質次之,蚓部髓質表達量最低。NTRK2在黃牛小腦半球髓質中表達量最高,顯著高于其他組織(P<0.05,圖6-B),蚓部皮質次之,小腦半球皮質表達量最低。
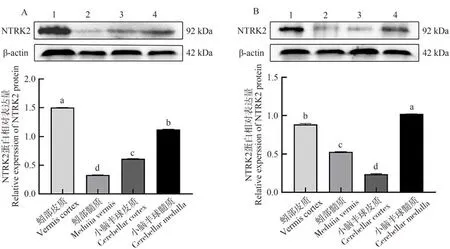
圖6 牦牛(A)與黃牛(B)小腦不同區域中NTRK2蛋白表達情況Fig.6 Expression of NTRK2 protein in different regions of the yaks (A) and cattles (B) cerebellum
與黃牛相比較,NT-4蛋白在牦牛小腦各區域中的表達量均高于黃牛(圖7-A);NTRK2蛋白的表達量在蚓部髓質和小腦半球髓質中低于黃牛或無差異,其余區域均高于黃牛(圖7-B)。
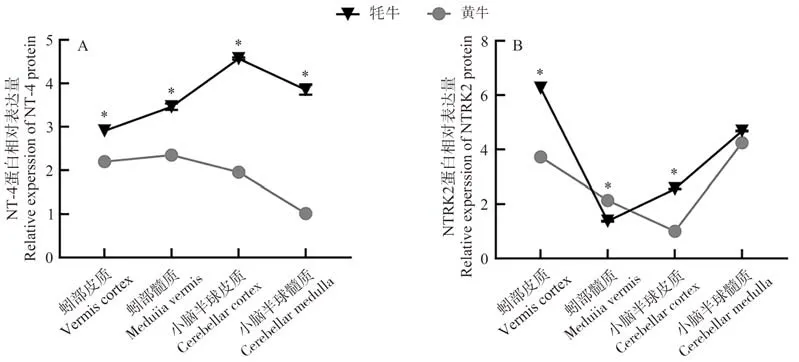
圖7 牦牛與黃牛小腦不同區域NT-4 (A)及NTRK2 (B)蛋白表達比較Fig.7 Comparative expression of NT-4 (A) and NTRK2 (B) in different regions of the yaks and cattles cerebellum
2.3 IHC檢測結果
通過HE染色觀察牦牛和黃牛小腦組織結構,結果如圖8、9所示,牦牛和黃牛小腦各區域組織結構相似。蚓部皮質和小腦半球皮質從淺到深均可分為分子層、浦肯野細胞層和顆粒層。分子層主要包括無髓神經細胞纖維和少量神經元,如胞體較大的籃狀細胞;浦肯野細胞層由排列整齊的浦肯野細胞胞體組成,胞體呈梨形;顆粒層則由密集的顆粒細胞和一些高爾基細胞組成。蚓部髓質和小腦半球髓質形態結構相似,主要有神經膠質細胞和神經纖維組成,并伴有少量血管。

圖8 牦牛小腦不同區域中NT-4和NTRK2蛋白分布Fig.8 Distribution of NT-4 and NTRK2 proteins in different regions of the yaks cerebellum
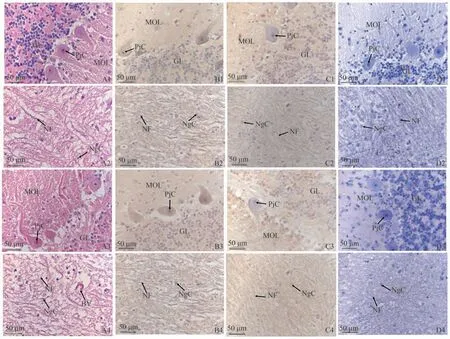
圖9 黃牛小腦不同區域中NT-4和NTRK2蛋白分布Fig.9 Distribution of NT-4 and NTRK2 proteins in different regions of the cattles cerebellum
通過免疫組織化學染色法檢測牦牛與黃牛小腦各區域NT-4及NTRK2的分布及定位。結果如圖8、9所示,黃牛小腦NT-4和NTRK2陽性細胞的分布和定位趨勢與牦牛一致,陽性產物主要分布于細胞質。在蚓部皮質和小腦半球皮質的分子層中,可見散亂分布的NT-4和NTRK2陽性反應,同時籃狀細胞的胞質中也有少量陽性表達。浦肯野細胞的胞質中亦有陽性產物分布,而顆粒細胞的胞質中也呈現大量細胞表達陽性產物。此外,在蚓部髓質和小腦半球髓質中,陽性著色多集中于神經元胞質中,在神經膠質細胞中也有少量陽性產物分布。
3 討論
NT-4作為神經營養因子家族中的重要成員之一,可控制脊椎動物神經元的存活和分化,同時在腦缺血缺氧后可通過NT-4-NTRK2信號傳導途徑發揮內源性保護作用[26]。本研究結果顯示,NT-4基因和蛋白在牦牛與黃牛小腦不同區域均有表達,牦牛NT-4蛋白在小腦各區域中表達水平顯著高于黃牛,該蛋白在小腦半球皮質中表達量最高,小腦半球髓質中表達量次之,在牦牛蚓部皮質中表達量最低。然而,NT-4基因在牦牛蚓部髓質中表達量最低。有研究報道,基因的轉錄和蛋白的翻譯存在時空差異[27],這可能是造成NT-4基因及其蛋白存在表達差異的主要原因。前人研究表明,機體長期處于高原低氧環境下,會使小腦半球浦肯野細胞中酶活性降低、供能不足、機體免疫能力減弱,進而引起炎癥反應[28-29]。此外,還有研究報道NT-4可通過降低促炎因子的表達水平、減輕神經炎癥以及改善神經功能對缺血缺氧性腦損傷產生保護作用[25,30]。髓質主要由密集的纖維束和神經網絡組成,其中神經膠質細胞是神經營養因子的主要來源,已被證明在控制神經發生、細胞遷移、軸突生長、維持血腦屏障以及控制體內平衡和血管張力等方面發揮重要作用[31-32]。由此推測NT-4可能在牦牛與黃牛小腦神經元的存活、生長以及功能發揮方面具有調節作用,牦牛小腦半球中NT-4的高表達可能也會參與神經-免疫-內分泌調控網絡,進而使腦組織更好的適應低氧環境。
NTRK2是原肌球蛋白受體激酶家族中的重要成員,也是NT-4的高親和力受體,NT-4對腦組織的神經保護作用需NTRK2的激活[33]。相關研究發現,當機體或細胞受到缺血缺氧刺激時,會誘導NT-4和NTRK2的表達增加,NT-4隨后誘導NTRK2形成二聚體,并自動發生磷酸化,磷酸化受體再通過相關途徑調節缺氧誘導因子-1α(HIF-1α)的表達[26]。本研究結果與上述研究結果相一致,即NT-4及NTRK2在牦牛蚓部皮質和小腦半球皮質中的表達量均顯著高于黃牛,說明二者協同對神經元發揮了內源性保護作用,但兩種因子在牦牛蚓部髓質的表達趨勢不一致,提示NT-4與NTRK2并非一一對應關系,可能與低親和力受體競爭NT-4并與之結合有關。
已有研究表明,在中樞神經和周圍神經系統中的大多數神經元中均可觀察到NT-4和NTRK2陽性表達[34],這與本研究中NT-4和NTRK2蛋白在牦牛和黃牛小腦各區域均有陽性表達結果一致,即在小腦各區域的皮質中主要定位于分子層、浦肯野細胞層和顆粒層的神經元胞質中,在髓質各區域中主要分布于神經纖維以及神經膠質細胞中,說明NT-4和NTRK2對小腦的神經保護作用主要依賴于以上神經元的存活來維持中樞神經系統內環境的穩定。但是,在成年恒河猴腦組織中NT-4僅存在于浦肯野細胞中,而在顆粒層未見陽性表達[35],這與本研究結果不一致,可能與物種差異以及機體耗氧量有關。此外,有研究顯示,NT-4在不同神經元中的表達水平會隨著缺血缺氧性損傷增強而升高[12],本研究發現,牦牛小腦各區域中NT-4和NTRK2蛋白免疫陽性反應強度整體強于黃牛,推測可能與牦牛長期處于低氧環境有關。腦組織受到低氧刺激時,上述細胞被激活并誘導NT-4和NTRK2表達,再通過HIF-1α調節參與糖代謝和血管生成的下游蛋白的表達,有助于牦牛適應高原低氧環境。
4 結論
NT-4和NTRK2在成年牦牛與黃牛小腦各區域中均有表達且存在顯著差異,與黃牛相比,兩種因子在牦牛小腦中的表達水平更高,提示NT-4和NTRK2除了共同參與調控機體正常的生理功能之外,也可能在牦牛小腦神經元及神經膠質細胞低氧適應過程中發揮內源性神經保護作用,進而使腦組織免受低氧損傷。
猜你喜歡
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
科學(2020年5期)2020-11-26 08:19:22
軟件(2020年3期)2020-04-20 01:45:18
商周刊(2018年15期)2018-07-27 01:41:20
敦煌學輯刊(2018年1期)2018-07-09 05:46:42
北京教育·普教版(2017年1期)2017-02-05 13:26:23
新疆農墾科技(2016年2期)2016-08-21 13:50:16
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
新疆財經大學學報(2015年3期)2015-12-10 03:49:15
