微管結合蛋白tau互作蛋白的篩選
2024-03-07 01:33:46王雄雄張圣權孫靜毛傳樨
湖北大學學報(自然科學版) 2024年2期
關鍵詞:實驗
王雄雄,張圣權,孫靜,毛傳樨
(湖北大學生命科學學院, 湖北 武漢 430062)
0 引言
微管是一種具有極性的細胞骨架,由α,β蛋白亞基組成的微管蛋白二聚體,微管蛋白二聚體頭尾相接形成微管原纖維,由13根這樣的原纖維縱向排列形成微管的壁。并且微管參與細胞分裂、細胞運動、胞內運輸、分泌、形態構成、組織器官發育等多種重要的生物學過程[1]。在微管的動態調節過程中,有兩類蛋白參與其調節過程[2]。一類是微管結合蛋白(microtubule associated protein, MAP)(如tau、MAP1和MAP4)和微管末端協助微管聚合的因子(如EB1和CLIP170)等;另一類是參與微管剪切的蛋白,包括Spastin、Katanin、Fidgetin等。
Tau作為重要的微管結合蛋白,促進微管的穩定,主要在中樞和周圍神經系統中表達,并富集于神經元軸突中[3]。Tau蛋白在結構上可以分為4個區域:N端投射區、脯氨酸富集區、微管結合結構域、C端域。Tau對于微管的穩定性就是通過微管結合結構域來實現的,對于神經元的發育和軸突的正常運輸發揮著重要作用[4-5]。
早在20世紀80年代就有研究報道,不可溶和過度磷酸化的tau是神經纖維纏結的主要成分,是阿爾茲海默癥的病理特征之一[6-9]。體外生化研究表明,過度磷酸化的tau不與微管結合,導致微管穩定性降低[10-11]。此外,tau突變導致17號染色體相關的帕金森綜合征的額顳葉癡呆中,其中一些突變已經被報道降低微管和tau結合親和力[12-13],強調了tau的正常微管穩定性動能喪失在神經退行性tau蛋白病的發病機制中的重要性。除了阿爾茲海默癥外,還有一些退行性疾病有tau的包涵體,這類tau相關的疾病統稱為Tauopathies[14], 如額顳葉變性癥、嗜銀顆粒病、肌萎縮性脊髓側索硬化癥和慢性創傷性腦病等[15-17]。由于tau與很多神經退行性疾病相關,研究tau的致病機制為治療相關的神經退行性疾病提供重要依據。
之前的報道顯示,在GMR-Gal4驅動下人的野生型tau(UAS-tau0N4R)表達可導致粗糙眼(rough eye)表型[18-20]。本研究通過該模型篩選微管異常的RNAi表達品系[21],并進一步通過成蟲盤凋亡細胞和行為學實驗驗證,尋找與tau共同調控視神經發育的基因,進而找到直接或間接與tau互作的蛋白。從而能夠為治療阿爾茲海默癥等和tau相關的神經退行性疾病提供潛在的藥物靶點。
1 材料和方法
1.1 果蠅品系
見表1。

表1 實驗所用果蠅品系
1.2 實驗所用儀器

表2 實驗所用儀器
1.3 實驗所用抗體
見表3。

表3 實驗所用抗體
1.4 實驗所用試劑
實驗所用試劑見表4。試劑配方分別見表5~7。

表4 實驗所用試劑

表5 解剖液HL 3.1

表6 4%多聚甲醛固定液(PFA)

表7 1×PBS緩沖液
1.5 實驗方法
1.5.1 果蠅粗糙眼觀察
將羽化后的果蠅成蟲在25 ℃,12 h光照:12 h黑暗的培養箱中飼養3 d,之后在-20 ℃冰箱分別將雌雄果蠅麻醉10 min,使用LEICA M205C顯微鏡觀察并拍攝果蠅眼睛。
1.5.2 果蠅幼蟲解剖
1)用實驗室公用鑷子從果蠅管中取出幼蟲(長鑷子方便取出幼蟲,解剖的短鑷子難以伸到管底),用解剖液洗去附著在幼蟲身體上的培養基,并將幼蟲放置在紙巾上蘸干。
2)將幼蟲放到樹脂解剖盤上,用解剖針分別固定幼蟲的頭部和尾部,然后調整針的位置,避免幼蟲拉太長,后續伸展不開。
1.4 統計學方法 采用SPSS 13.0統計學軟件進行數據分析,計量資料以表示,治療前后自身對照均數比較采用配對t檢驗,2組間均數比較采用獨立樣本t檢驗,檢驗水平α=0.05。
3)用移液槍吸取解剖液,加入到幼蟲表面,使其完全浸沒。用解剖剪刀在幼蟲尾部剪開一個口子,然后用解剖剪刀沿著中線剪至頭部。
4)用鑷子輕輕挑去幼蟲的內臟,注意不要碰到eye disc,以免破壞組織完整性。
5)用4根解剖針分別固定頭尾兩邊的位置,使幼蟲平鋪開,呈現出片狀。
1.5.3 eye disc免疫染色
1)在解剖好的幼蟲(保存完整的eye disc)中加入4% PFA固定液覆蓋表面,室溫固定40 min。
2)固定完后,吸去固定液,取下幼蟲放入2 mL EP管中,加入0.2%的PBST溶液(1×PBS+0.2% TritonX-100),然后將樣品放在側擺洗脫固定液1 h,每10~15 min換一次EP中PBST溶液。
3)洗完固定液后,吸去EP管中的PBST溶液,加入200 μL 0.2% PBST+5% 山羊血清,在側擺搖床上室溫封閉40 min;封閉完后加入1∶200 Dcp1和1∶100 elav抗體(用0.2% PBST溶液配制 抗體),將樣品放入4 ℃冰箱的側擺中,對樣品進行低溫孵育過夜。
4)待一抗孵育完成后,吸去抗體,將樣品放入室溫搖床下,用0.2% PBST溶液洗脫,在此期間洗脫1 h,每10~15 min換一次EP管中PBST溶液;洗完抗體后,吸去抗體,加入1∶800 Goat anti rabbit 555和1∶800 Goat anti mouse 488二抗(用0.2% PBST溶液配制 二抗),將樣品放入4 ℃冰箱的側擺中,對樣品進行低溫孵育過夜;并做避光處理。
5)待二抗孵育完成后,吸去二抗,將樣品放入室溫搖床下,用0.2% PBST溶液洗脫二抗,在此期間洗脫1 h,每10~15 min換一次EP管中PBST溶液。
6)二抗洗脫完后,取出幼蟲,用鑷子小心地將解剖好幼蟲上的eye disc取出,然后放在載玻片,擺放完成后,用紙巾吸去殘留在組織的PBST,小心在組織旁邊滴一滴封片油,輕輕地蓋上蓋玻片,避免有氣泡影響后續的觀察,并在蓋玻片周圍涂上指甲油,避免蓋玻片滑動破壞組織,封片完后寫上對應的基因型,放入4 ℃冰箱保存,后續用共聚焦顯微鏡拍攝。
1.5.4 果蠅的睡眠監測
本實驗所用的睡眠監測采用的是pySolo v2.0系統[22]。pySolo系統的工作原理是采用高分辨攝像頭來記錄每分鐘果蠅的移動距離,以此來判定果蠅是否處于睡眠狀態。系統中,我們定義果蠅在靜止超過5 min時判定為睡眠狀態,系統每次采樣16只雄果蠅,pySolo系統的優勢就在于其高效、準確、靈敏,能準確反映果蠅每分鐘的活動狀態。
1.5.5 果蠅eye disc凋亡細胞平均熒光面積比的統計
本實驗利用ImageJ進行熒光定量分析,統計elav抗體標記區域內的凋亡信號(death caspase-1,Dcp1)分布面積,并用對照組對所有組別進行標準化。同時,通過t檢驗比較實驗組與對照組之間的差異。
2 結果
2.1 以rough eye系統為模型進行微管互作蛋白的篩選
我們在早期的實驗中使用C57-Gal4(在肌肉中特異性表達的Gal4)與清華大學果蠅庫中的541株RNAi基因進行雜交,最終篩選出了45株有表型的RNAi[21]。在果蠅視網膜中表達人的野生型tau會導致中度的粗糙眼表型,其特征是果蠅單眼陣列的紊亂和眼睛大小的減小[23]。Tau誘導的粗糙眼表型是篩選可能增強或抑制tau蛋白的相關互作蛋白和和藥理學試劑的優良系統[24]。有研究利用該模型篩選出了可能與tau互作的候選基因,其中與細胞骨架相關的包括微絲結合蛋白和沿微管運輸的馬達蛋白[24-26]。我們在GMR-Gal4驅動下表達人的野生型tau(UAS-tau0N4R)可導致rough eye表型,與之前的報道一致[18-20]。為了篩選出與tau蛋白相互作用的蛋白,我們在高表達tau0N4R的背景下表達之前篩選出的45株具有表型的RNAi。在進一步的篩選中,我們篩選出了6株RNAi,這些RNAi能夠在雌性和雄性果蠅的粗糙眼表型中增強,相關基因分別是CG32068、CG9886、CG5588、CG3298、CG7834、CG18001,如圖1所示。篩選出雌性和雄性都粗糙眼表型減弱的5株RNAi,相關基因分別是CG33102、CG16827、CG8171、CG3697、CG3325,如圖2所示。說明CG32068、CG9886、CG5588、CG3298、CG7834、CG18001這些RNAi可能誘導tau毒性,導致感光神經元喪失增多,粗糙眼表型增強,而CG33102、CG16827、CG8171、CG3697、CG3325則相反。

圖1 6個基因在tau高表達背景下敲降后果蠅粗糙眼表型增強敲降后果蠅粗糙眼增強。從圖中可以看出,敲降CG32068、CG9886、CG5588、CG3298、CG7834、CG18001后粗糙眼表型增強。圖中對照組基因型為GMR-Gal4/+;UAS-tau0N4R/+,實驗組為GMR-Gal4/CyO;UAS-tau0N4R背景下敲降基因
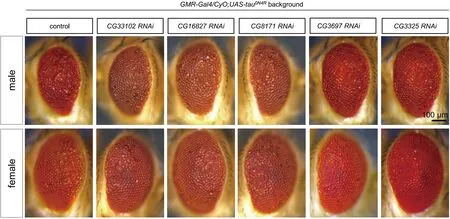
圖2 5個基因在tau高表達背景下敲降后果蠅粗糙眼表型減弱敲降后果蠅粗糙眼減弱。從圖中可以看出,敲降CG33102、CG16827、CG8171、CG3697、CG3325后粗糙眼表型減弱。圖中對照組基因型為GMR-Gal4/+;UAS-tau0N4R/+,實驗組為GMR-Gal4/CyO;UAS-tau0N4R背景下敲降基因
2.2 敲降后影響eye disc的凋亡
發育中的果蠅眼組織,即果蠅眼睛成蟲盤,是研究不同細胞凋亡反應的理想系統,因為每種細胞類型的命運是已知的,并且已有豐富的細胞標記來跟蹤它們[27]。在早期幼蟲階段,眼睛成蟲盤中的細胞不斷增殖,以產生眼睛所需的細胞團[27]。為了驗證粗糙眼的表型是否會體現在eye disc凋亡,我們在UAS-tau0N4R背景下使用GMR-Gal4和rough eye表型增強的RNAi果蠅雜交,通過果蠅凋亡信號通路中的效應caspase(Dcp1)的免疫熒光染色來檢測神經元凋亡的程度。以elav標記的神經元區域為范圍(圖3A),統計Dcp1信號覆蓋面積(圖3B和3C)。結果顯示,敲降CG32068、CG3298、CG7834,凋亡細胞數量增加,而敲降CG9886、CG5588、CG18001凋亡細胞數量無變化。而在另外rough eye表型減弱的RNAi中,發現敲降CG16827、CG8171凋亡細胞數量增加,敲降CG3697凋亡細胞數量減少,而敲降CG33102、CG3325凋亡細胞數量無變化。以上結果說明CG32068、CG3298、CG7834、CG16827、CG8171和CG3697可能與tau有互作。
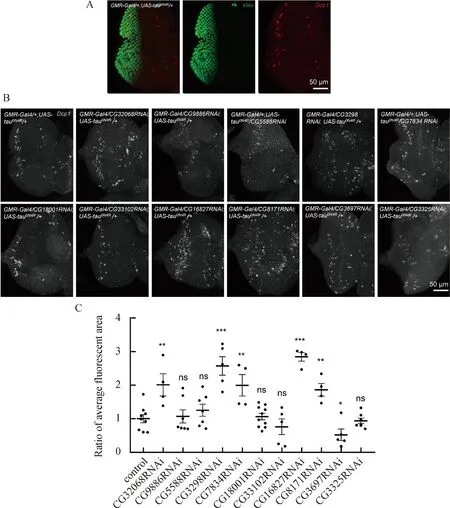
圖3 敲降后果蠅eye disc凋亡細胞染色和平均熒光面積比A)對照組果蠅的eye disc免疫染色。elav標記所有神經元,用紅色表示;Dcp1標記凋亡細胞,用綠色表示。B)敲降后果蠅eye disc凋亡細胞數量呈現增加或無變化的免疫染色圖。由圖可以看出,敲降CG32068、CG3298、CG7834 、CG16827、CG8171凋亡細胞數量增加,敲降CG9886、CG5588、CG18001、CG33102、CG3325無變化,而敲降CG3697凋亡細胞數量減少。圖中凋亡細胞是用抗體Dcp1標記。C)敲降后果蠅eye disc凋亡細胞平均熒光面積比的統計,以elav為范圍統計該區域的平均熒光面積,并利用t檢驗比較實驗組與對照組之間的差異。圖中對照組基因型為GMR-Gal4/+;UAS-tau0N4R/+,實驗組為GMR-Gal4/CyO;UAS-tau0N4R背景下敲降基因
2.3 敲降后對晝夜睡眠節律的影響
晝夜振蕩在細胞水平上由生物鐘表示,生物鐘是由環境光形成的分子程序-關鍵時鐘蛋白對光敏感,因此在光振蕩條件下,它們的水平有節奏地上升和下降[28]。由于具有粗糙眼表型增強的RNAi果蠅可能會導致感光神經元喪失增加,因此它們的光敏感性可能會受到影響,從而進一步影響其晝夜節律。本實驗在25 ℃ 12 h光照:12 h黑暗條件下,利用睡眠監測系統pySolo v2.0系統,對敲降CG32068、CG9886、CG5588、CG3298、CG7834、CG18001成蟲果蠅進行了晝夜睡眠監測,在監測4日后,將監測數據導入高通量數據分析軟件RStudio對果蠅的活動及睡眠進行分析,分析結果如圖4所示。從果蠅睡眠曲線圖可以看出,與對照相比,在關燈后,敲降CG32068,果蠅對于光線改變不敏感,而敲降CG9886、CG5588、CG3298、CG7834、CG18001無明顯變化。綜合以上結果,說明CG32068RNAi可能與tau互作,導致粗糙眼表型增強和eye disc凋亡細胞增加,從而導致感光神經元喪失和眼睛發育缺陷,進而導致對光線改變不敏感,影響果蠅的晝夜節律。
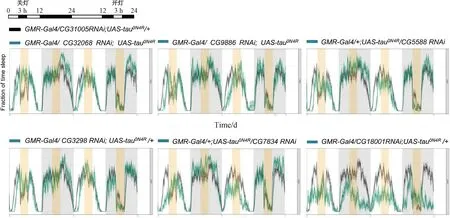
圖4 睡眠監測模式和敲降后睡眠監測圖在25 ℃ LD條件下,果蠅適應2天,在第3天12 L關燈3 h, 以及在第4天的12 D開燈3 h, 觀察突然關燈或開燈,果蠅對光是否敏感。與對照相比,在關燈后,敲降CG32068后,果蠅對于光線改變不敏感,果蠅沒有被喚醒,而敲降CG9886、CG5588、CG3298、CG7834、CG18001明顯變化。圖中對照組基因型為GMR-Gal4/ CG31005RNAi;UAS-tau0N4R/+,實驗組為GMR-Gal4/CyO;UAS-tau0N4R背景下敲降基因
3 討論
據研究顯示,粗糙眼表型增強會引發感光神經元的喪失[29]。在本次實驗中,篩選出的11株具有粗糙眼表型增強或減弱的RNAi果蠅,可能是由于對這些基因進行敲降導致感光神經元數量的增加或減少引起的。
在果蠅的早期幼蟲階段,眼睛成蟲盤中的細胞持續增殖,以形成眼睛所需的細胞集合[27]。在本實驗中,篩選出5株 RNAi果蠅表現出凋亡細胞數量的增加,1株RNAi凋亡細胞數量減少,這可能是由于對這些基因進行敲降導致細胞增殖受限或促進所引起。
通過本實驗的篩選,發現候選基因CG32068在果蠅的成蟲盤細胞凋亡、粗糙眼表型及行為學方面表現出結構和功能的一致性。進一步查閱相關文獻發現,該基因參與了果蠅甲硫氨酸挽救途徑[30]。接下來我們計劃進行以下實驗工作:首先,通過對果蠅喂食甲硫氨酸的實驗來驗證其是否可以挽救粗糙眼和凋亡等表型,并購買其他的工具果蠅來驗證前期得出的實驗結果。其次,在C57-Gal4/CyO;UAS-tauV337 M背景下敲降該基因,觀察微管表型和進行蛋白印跡實驗,以檢測該基因與微管結合蛋白tau之間的互作情況。后續的計劃正在進行中,需要進一步深入研究該候選基因在果蠅發育過程中的其他作用及其調節機制。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
