兩種養殖蛙類肌肉品質、腸道形態學及微生物構成比較分析
2024-03-06 01:45:38王志龍王廣軍郁二蒙雷小婷龔望寶
南方水產科學 2024年1期
王志龍 ,夏 耘,謝 駿,舒 銳,王廣軍,郁二蒙,雷小婷,龔望寶
1. 水產科學國家級實驗教學示范中心 (上海海洋大學),上海 201306
2. 中國水產科學研究院珠江水產研究所/農業農村部熱帶亞熱帶水產資源利用與養殖重點實驗室,廣東 廣州 510380
3. 廣東省星蛙農業科技有限公司,廣東 肇慶 526070
牛蛙 (Lithobatescatesbeiana) 原產于北美洲,具有繁殖快、適應性強、生長迅速等優點,已成為我國主要養殖蛙類[1];泰國虎紋蛙 (Hoplobatrachus tigerinus) 是廣東省近年來引進的優良水產養殖蛙類,其養殖投入低、養殖周期短且生長快,現已初步形成產業規模[2]。由于近幾年養殖蛙市場需求量大,養殖戶大多采用小池高密度養殖模式,且不同養殖品種均使用同一種 (同品牌) 商業人工配合飼料。同種人工飼料是否適宜不同養殖品種的生長和消化,相應的蛙產品肌肉品質是否能滿足消費者需求以及如何科學開發飼料配方是養殖蛙產業研究的熱點問題。肌肉品質是進入消費市場的重要評價指標,而腸道菌群一定程度上可反映養殖蛙的消化特性,這些基礎數據對蛙類養殖和配合飼料的可持續開發至關重要。
肌肉品質包括營養價值和質構特性等[3],肌肉營養價值通常由蛋白質和氨基酸、脂肪和多不飽和脂肪酸的組成來作為評估指標[4],肌肉質構特性則由肌肉的硬度、黏性、彈性、咀嚼性、膠黏性和內聚性等質構參數來體現[3]。有關養殖蛙的肌肉營養價值,有學者針對黑斑蛙 (Pelophylaxnigromaculatus)[5]、棘胸蛙 (Quasipaaspinosa)[6]、泰國虎紋蛙[7]、東北林蛙 (Ranadybowskii) 和牛蛙[8]肌肉營養組成開展了部分研究。然而,國內外有關養殖蛙類肌肉質構特性的研究報道較少,而肌肉質構是產品進入消費市場的重要評價指標。
腸道菌群與宿主的消化、營養吸收、代謝和生理功能密切相關[9]。目前,已報道腸道菌群的養殖蛙類有棘胸蛙[10]、黑斑蛙[11]、虎紋蛙 (H.chinensis)[12]等,但對泰國虎紋蛙和牛蛙腸道菌群的研究較少。開展人工養殖條件下泰國虎紋蛙和牛蛙腸道顯微結構和微生物群落結構研究,有助于分析其對飼料的消化吸收及生長特性。本研究通過測定泰國虎紋蛙和牛蛙的肌肉營養組成和肌肉質構特性對其肌肉品質進行全面評價,通過高通量測序技術和組織切片技術分析兩種養殖蛙的腸道微生物群落構成和顯微結構,旨在全面了解泰國虎紋蛙和牛蛙的肌肉品質和腸道菌群差異,為養殖蛙類的開發利用及配合飼料的研制提供基礎資料。
1 材料與方法
1.1 樣品采集
鮮活的成年雄性牛蛙和泰國虎紋蛙 (投喂同種膨化配合飼料) 取自廣東省星蛙農業科技有限公司花都養殖基地。采用小池高密度養殖模式,養殖密度為140 只·m?2,養殖水位為5~10 cm,水溫為 (28±2) ℃。配合飼料營養成分組成 (w):水分9.34%、粗蛋白質41.90%、粗脂肪5.78%、粗灰分9.48%。同一種蛙隨機選擇大小均一、形態相近的個體各6 只,泰國虎紋蛙體質量為 (125.28±2.56) g,牛蛙體質量為 (151.87±3.15) g。毀髓處死后,每只蛙取1 cm 大小的腿部肌肉組織,并于前腸相同腸道部位切取腸道組織各1 cm,用4% (φ) 多聚甲醛溶液固定,用于組織切片制作;將其余含有內容物的腸道組織迅速取出,液氮速凍后于?8 0 ℃保存,用于腸道菌群分析;沿著肌纖維剪取腿部肌肉組織,切成1 cm×1 cm×2 cm 的長方體組織塊,用于質構特性檢測;腿部其余肌肉用于營養成分測定。每組做6 個重復 (n=6)。
1.2 肌肉營養成分測定
按照鮮質量計算各營養成分的含量。參照GB 5009.5—2016 (凱氏定氮法)、GB 5009.6—2016 (索氏抽提法)、GB 5009.3—2016 (直接干燥法) 和GB 5009.4—2016 (第一法) 分別檢測兩種蛙肌肉的粗蛋白、粗脂肪、水分和灰分含量。使用全自動氨基酸分析儀參照GB 5009.124—2016 測定氨基酸組成;由于采用酸水解處理樣品,色氨酸被酸解,未測定。使用氣相色譜儀參照GB 5009.168—2016 分析脂肪酸種類和含量。根據聯合國糧農組織/世界衛生組織 (FAO/WHO) 提出和1991 年中國預防醫學科學院提出的氨基酸評分模式,計算氨基酸評分(Amino acid score, AAS)、化學評分 (Chemical score,CS) 和必需氨基酸指數 (Essential amino acid index,EAAI)[13]。
1.3 肌肉質構測定
將肌肉組織塊放在CT3 質構儀 (Brookfield 公司,美國) 上,探頭為P35,采用TPA 模式進行測定,測前速度為2 mm·s?1,測試速度為1 mm·s?1,測后速度為5 mm·s?1,2 次壓縮的時間間隔為2 s,壓縮比為25%,對肌肉質構特性進行測定。
1.4 組織切片觀察
組織切片:對肌肉和腸道組織按照固定、脫水、透蠟、包埋、切片和染色的順序進行操作,最終制作成組織切片。按照S=πr2計算肌纖維直徑(S為肌纖維橫切面面積;r為肌纖維橫切面半徑;π 為圓周率) ;并測量統計不同區域內肌纖維的數量和面積,計算肌纖維密度。每個樣本對300 條肌纖維進行統計。
1.5 腸道菌群高通量測序
按照EZNA?Soil DNA Kit 試劑盒 (Omega Biotek) 說明書提取腸道菌群總DNA。DNA 濃度和純度使用NanoDrop2000 檢測。以正向引物338F (5'-ACTCCTACGGGAGGCAGCA-3') 和反向引物806R(5'-GGACTACHVGGGTWTCTAAT-3') 擴增16S rRNA基因V3—V4 區。20 μL PCR 反應體系為:10 μL 2×Pro Taq,正向引物和反向引物 (5 mmol·L?1) 各0.8 μL,10 ng DNA 模板,ddH2O 補足至20 μL。PCR 過程:95 ℃ 3 min;95 ℃ 30 s,60 ℃ 30 s,72 ℃ 45 s,29 個循環;最后72 ℃延伸10 min。利用AxyPrep DNA Gel Extraction Kit 進行PCR 產物純化,使用Illumina 測序專用的TruSeqTMDNA Sample Prep Kit 試劑盒建立基因文庫。構建好的文庫通過Illumina Miseq PE300 平臺進行測序,由上海美吉生物醫藥科技有限公司完成。對獲得的原始測序序列進行過濾、雙端拼接,得到優化序列(Tags);將優化序列進行聚類,劃分操作分類單元(Operational taxonomic units, OTU),并根據OTU 的序列組成得到其物種分類。基于OTU 分析結果,在美吉生物云平臺 (https://cloud.majorbio.com) 上利用R 語言 (version 3.3.1) 軟件進行Alpha 多樣性指數分析、顯著物種差異分析及功能差異預測分析。具體測定和分析方法參照Xia 等[14]。
1.6 統計學分析
2 結果
2.1 常規營養成分
如表1 所示,牛蛙的粗脂肪含量顯著高于泰國虎紋蛙,而泰國虎紋蛙的灰分含量顯著高于牛蛙(P<0.05)。泰國虎紋蛙和牛蛙的水分含量和粗蛋白含量均無顯著性差異 (P>0.05)。

表1 泰國虎紋蛙和牛蛙每100 g 肌肉常規營養成分比較Table 1 Comparison of conventional nutrients per 100 g of muscle between H. tigerinus and L. catesbeian g
2.2 氨基酸含量
在兩種養殖蛙肌肉中均檢測到16 種氨基酸,其中必需氨基酸7 種,非必需氨基酸9 種 (表2)。兩種蛙的甘氨酸含量均最高,其次是天門冬氨酸和異亮氨酸。除泰國虎紋蛙的蘇氨酸含量顯著高于牛蛙外 (P<0.05),其余氨基酸之間無顯著性差異(P>0.05)。兩種蛙的必需氨基酸 (Essential amino acids, EAA)、非必需氨基酸 (Non-essential amino acids, NEAA)、呈味氨基酸 (Flavor amino acids,FAA)、支鏈氨基酸 (Branched chain amino acids,BCAA)、芳香氨基酸 (Aromatic amino acids, AAA)與氨基酸總量 (Total amino acids, TAA) 均無顯著性差異 (P>0.05)。
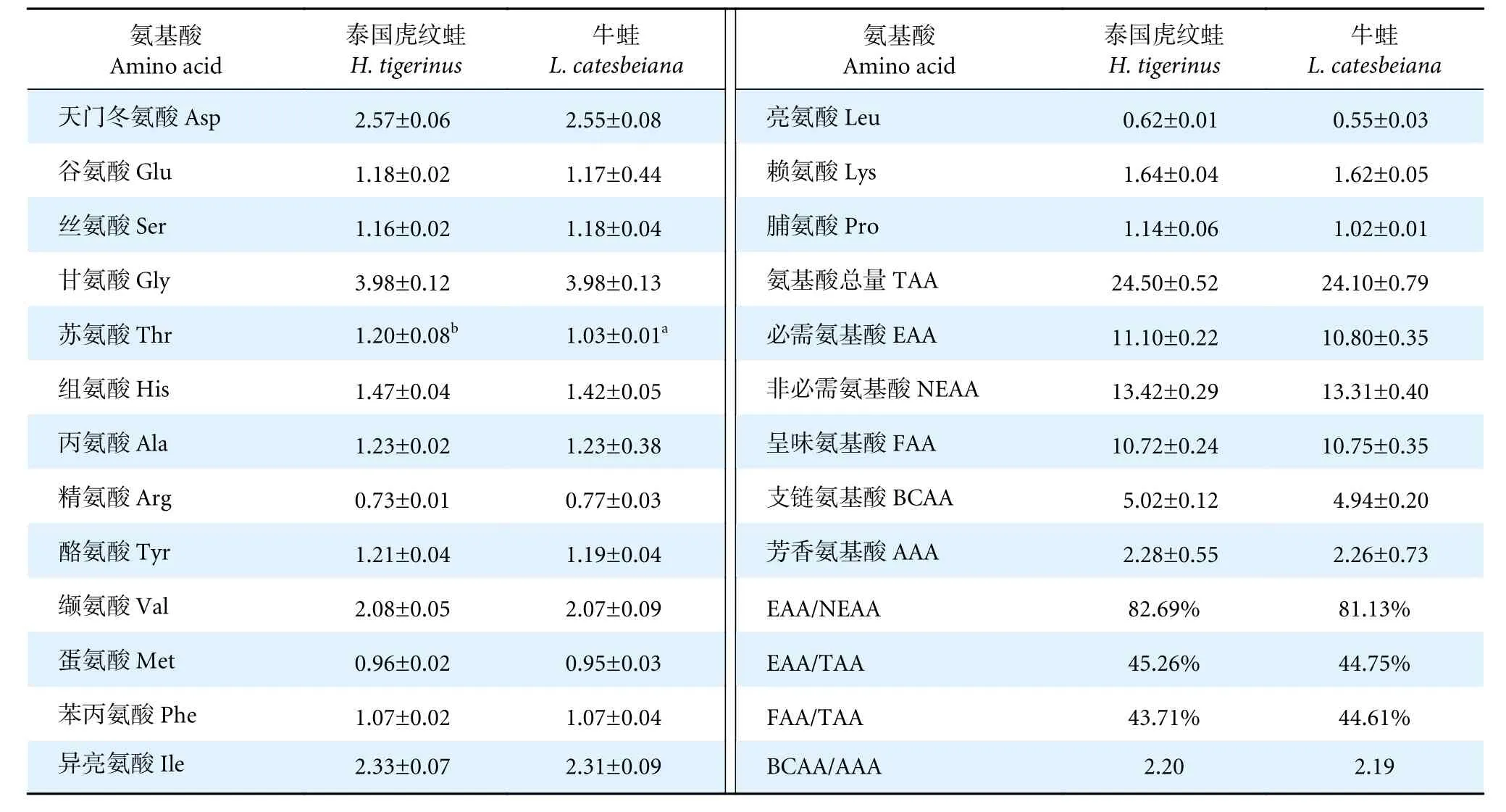
表2 泰國虎紋蛙和牛蛙每100 g 肌肉氨基酸比較Table 2 Comparison of amino acids per 100 g of muscle of H. tigerinus and L. catesbeiang
2.3 氨基酸營養品質評價
對泰國虎紋蛙和牛蛙肌肉必需氨基酸進行氨基酸評分(表3),兩者除異亮氨酸的AAS (0.58)外,其他均小于0.50;兩者異亮氨酸的化學評分最高。
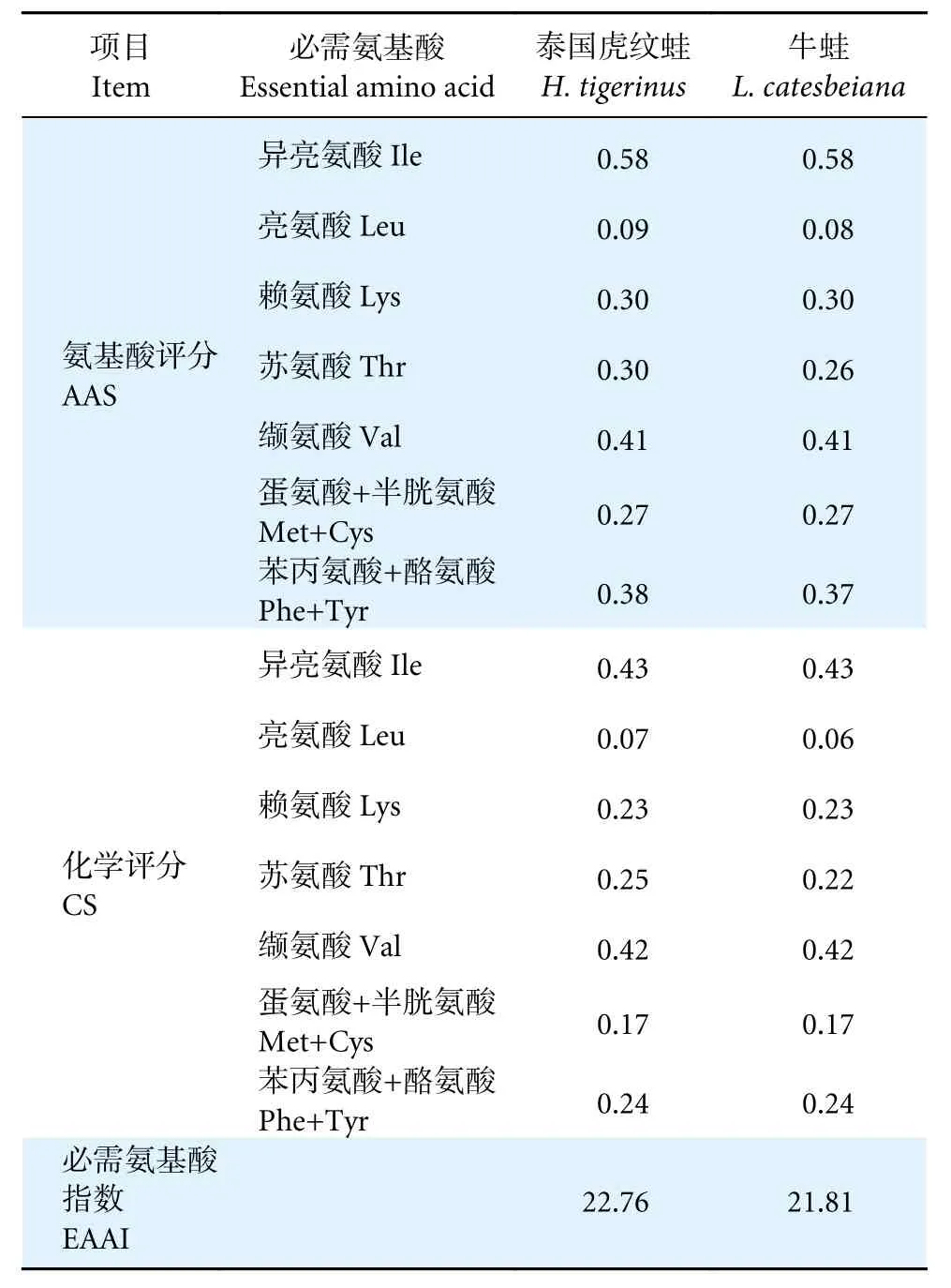
表3 泰國虎紋蛙和牛蛙肌肉氨基酸營養價值評價Table 3 Evaluation of amino acids traits in muscles of H. tigerinus and L. catesbeiana
2.4 脂肪酸含量
兩種蛙的肌肉中均含有22 種脂肪酸 (表4),包括飽和脂肪酸酸 (Saturated fatty acids, SFA)8 種,不飽和脂肪酸 (Unsaturated fatty acids, UFA)14 種;單不飽和脂肪酸 (Monounsaturated fatty acids, MUFA) 6 種,多不飽和脂肪酸酸 (Polyunsaturated fatty acids, PUFA) 8 種。從總體脂肪酸含量來看,泰國虎紋蛙為多不飽和脂肪酸>飽和脂肪酸>單不飽和脂肪酸,牛蛙為多不飽和脂肪酸>單不飽和脂肪酸>飽和脂肪酸,其中泰國虎紋蛙肌肉中飽和脂肪酸含量顯著高于牛蛙 (P<0.05);牛蛙肌肉中單不飽和脂肪酸含量顯著高于泰國虎紋蛙 (P<0.05),而多不飽和脂肪酸含量顯著低于泰國虎紋蛙 (P<0.05)。
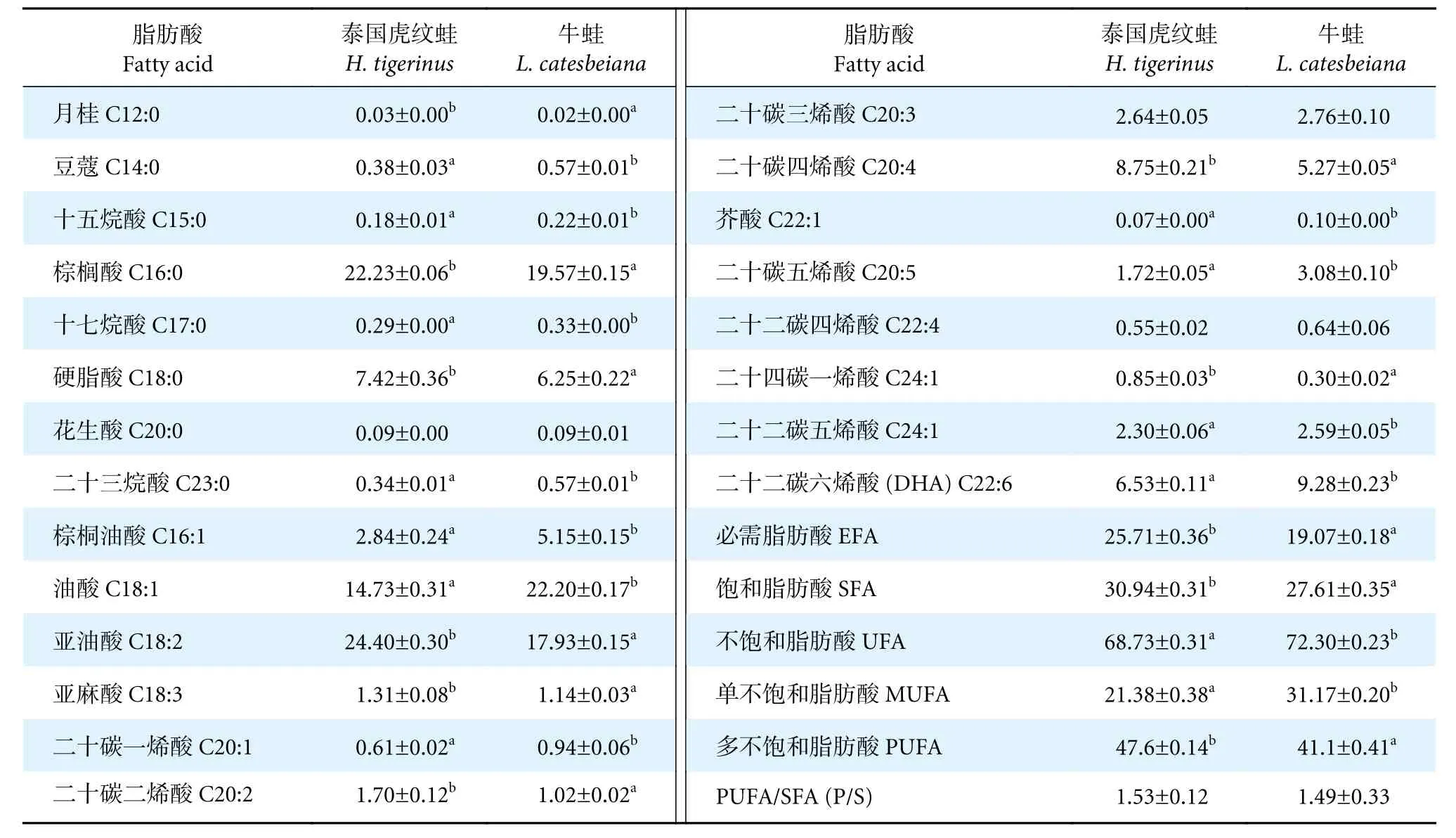
表4 泰國虎紋蛙和牛蛙每100 g 肌肉脂肪酸比較Table 4 Comparison of fatty acids in per 100 g muscles of H. tigerinus and L. catesbeianag
泰國虎紋蛙的必需脂肪酸 (Essential fatty acids,EFA) 含量顯著高于牛蛙 (P<0.05),其中,亞油酸和亞麻酸顯著高于牛蛙 (P<0.05)。SFA 中,除C20:0 含量在兩種蛙間不顯著外 (P>0.05),其余各脂肪酸含量在兩者之間均有顯著性差異 (P<0.05)。UFA 中,除C20:3 和C22:4 在兩種蛙間不顯著外(P>0.05),其余各脂肪酸含量在兩者間均有顯著性差異 (P<0.05),此外,P/S 介于1.49~1.53,符合WHO 建議值 (P/S≥0.4)。
2.5 肌肉質構特性
如表5 所示,牛蛙肌肉的硬度、咀嚼性、膠黏性均顯著高于泰國虎紋蛙 (P<0.05),而黏性顯著低于泰國虎紋蛙 (P<0.05)。此外,兩者肌肉的彈性和內聚性無顯著性差異 (P>0.05)。
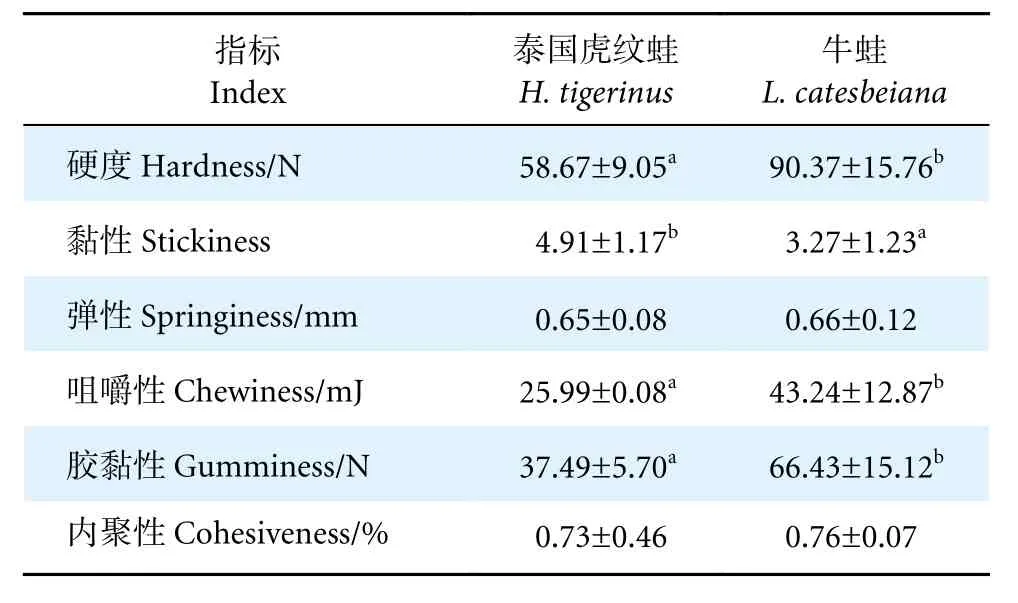
表5 泰國虎紋蛙和牛蛙肌肉質構特性的比較Table 5 Comparison of texture properties in muscle of H. tigerinus and L. catesbeiana
2.6 肌肉和腸道形態結構比較
泰國虎紋蛙和牛蛙的肌肉和腸道形態結構如圖1 所示。兩者肌纖維完整,排列規則,纖維大小與間隙均勻一致。牛蛙肌纖維直徑顯著大于泰國虎紋蛙 (P<0.05),而兩者的肌纖維密度無顯著性差異(P>0.05,表6)。
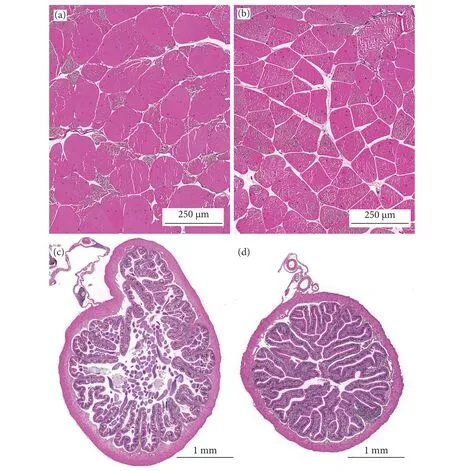
圖1 泰國虎紋蛙和牛蛙腸道和肌肉的組織結構Fig. 1 Tissue structures of intestine and muscle of two frogs species
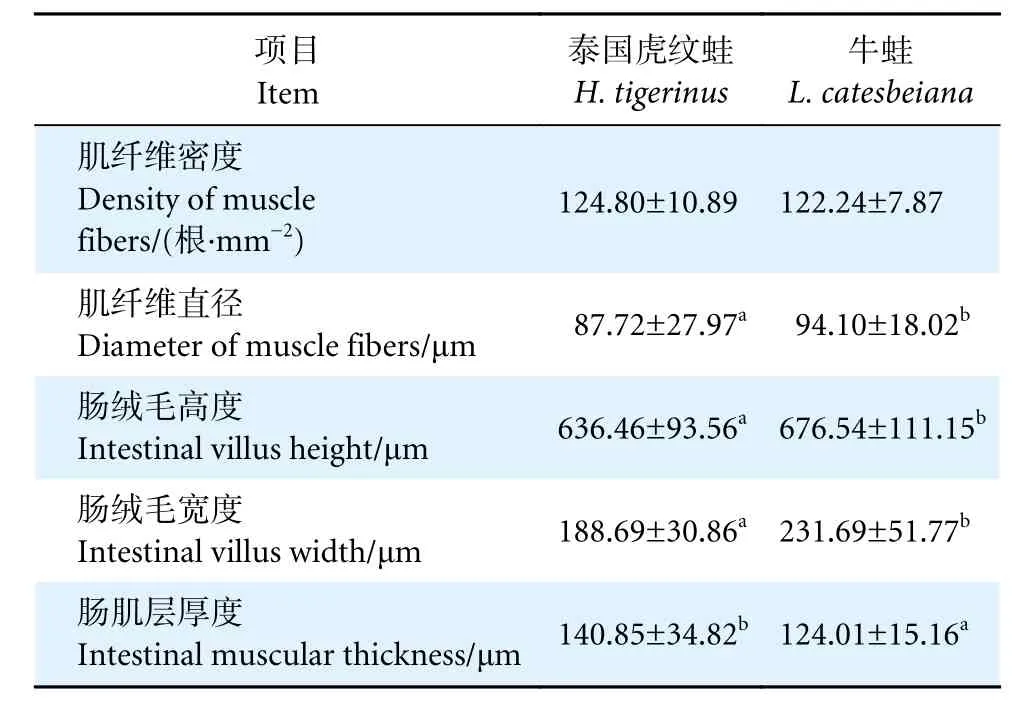
表6 泰國虎紋蛙和牛蛙肌纖維和腸道組織結構比較Table 6 Comparison of muscle fibe and intestine morphology of H. tigerinus and L. catesbeiana
泰國虎紋蛙和牛蛙的腸道組織結構正常,腸壁較厚,結構完整性較好,腸絨毛密且粗壯,末端無破損,在兩者的腸絨毛上皮細胞間均可觀察到大量的杯狀細胞。牛蛙的腸絨毛高度和寬度顯著高于泰國虎紋蛙 (P<0.05),而腸肌層厚度則顯著低于后者(P<0.05)。
2.7 腸道微生物群落結構
2.7.1 Alpha 多樣性指數
高通量測序篩選后共獲得945 594 個有效序列,序列長度為201~451 bp,平均長度為426 bp。按照97%的序列相似性可聚類為618 個 OTU。Alpha 多樣性分析顯示,兩種養殖蛙Good_coverage 均高于0.99,說明樣本中腸道微生物被充分檢出,測序質量良好 (表7)。微生物群落多樣性由Shannon 和Simpson 指數來反映,微生物群落豐富度由Ace 和Chao1 指數來反映。泰國虎紋蛙Shannon 指數顯著高于牛蛙 (P<0.01),表明泰國虎紋蛙腸道微生物群落多樣性高于牛蛙。泰國虎紋蛙腸道的Ace 和Chao1 指數顯著高于牛蛙 (P<0.05),表明泰國虎紋蛙腸道微生物的物種豐富度高于牛蛙。

表7 Alpha 多樣性指數Table 7 Alpha diversity indices of six samples
2.7.2 OTU 分布
兩種養殖蛙之間有98 個 (15.86%) 相同OTU,泰國虎紋蛙有487 個 (78.80%) 獨有OTU,牛蛙有33 個 (5.34%) 獨有OTU (圖2)。泰國虎紋蛙的OTU 數量顯著高于牛蛙 (P<0.05),這與Shannon指數的結果一致。

圖2 OTU 水平下兩種養殖蛙腸道微生物的Veen 圖Fig. 2 Veen diagrams of intestinal microbiota in H. tigerinus and L. catesbeiana at OTU level
2.7.3 基于門水平腸道菌群分析
如圖3 所示,牛蛙腸道優勢菌門是厚壁菌門(92.26%) 和變形菌門 (7.36%)。泰國虎紋蛙優勢菌門是厚壁菌門 (63.41%)、變形菌門 (19.43%)、梭桿菌門 (8.70%) 和擬桿菌門 (5.76%)。單因素方差分析顯示,牛蛙腸道的厚壁菌門豐度顯著高于泰國虎紋蛙 (P<0.05),而放線菌門豐度顯著低于后者(P<0.05)。
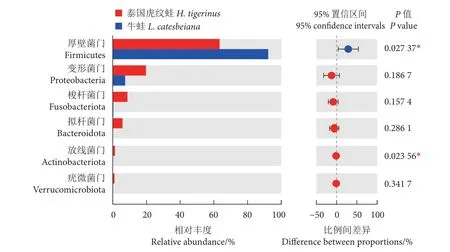
圖3 門水平上的腸道菌群組成Fig. 3 Intestinal microbiota composition at phylum level
2.7.4 基于屬水平腸道菌群分析
如圖4 所示,泰國虎紋蛙腸道菌群中豐度排名前5 的菌屬為乳球菌屬 (Lactococcus, 32.34%)、埃希桿菌-志賀菌屬 (Escherichia-Shigella, 10.36%)、腸球菌屬 (Enterococcus, 10.11%)、鯨桿菌屬 (Cetobacterium, 8.62%) 和魏斯氏菌屬 (Weissella, 6.90%);牛蛙腸道優勢菌屬為乳桿菌屬 (Lactobacillus,59.30%)、魏斯氏菌屬 (15.37%)、明串珠菌屬 (Leuconostoc, 10.98%)、檸檬酸桿菌屬 (Citrobacter,6.82%) 和乳球菌屬 (5.19%)。單因素方差分析顯示,泰國虎紋蛙的乳球菌屬和埃希桿菌-志賀菌屬豐度顯著高于牛蛙 (P<0.05),而乳桿菌屬和明串珠菌屬豐度顯著低于后者 (P<0.05)。
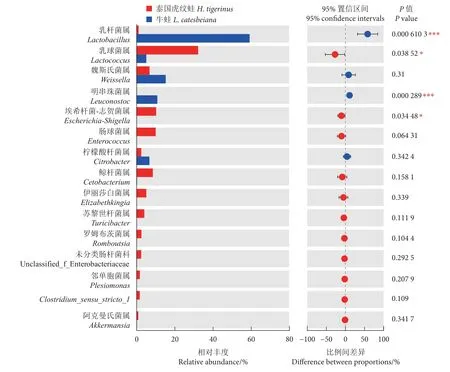
圖4 屬水平上的腸道菌群組成Fig. 4 Intestinal microbiota composition at genus level
2.7.5 腸道微生物功能預測
泰國虎紋蛙和牛蛙腸道菌群功能基因中占比較高的均為與新陳代謝相關的功能基因 (圖5),包括碳水化合物代謝、氨基酸代謝、輔助因子和維生素代謝、能量代謝和脂代謝等新陳代謝功能基因,此外,膜轉運和信號轉導等功能基因也在兩者腸道菌群中占有優勢。功能預測顯示泰國虎紋蛙和牛蛙腸道菌群可能主要參與宿主營養物質轉運及代謝等相關進程。
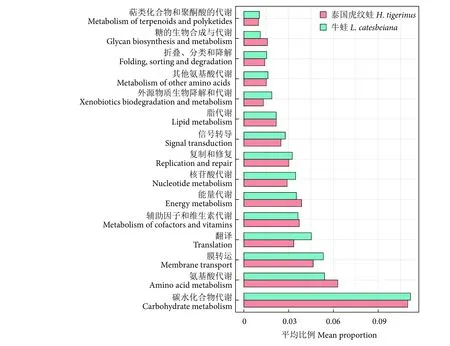
圖5 泰國虎紋蛙和牛蛙腸道微生物功能預測Fig. 5 Functional prediction of intestinal microbiota in H. tigerinus and L. catesbeiana
3 討論
3.1 肌肉常規營養成分
水分是肌肉含量最多的成分,一定程度上來說,肌肉水分充足則肉質更鮮嫩[8],這也是兩種養殖蛙類口感鮮嫩多汁的主要原因。灰分是肌肉經高溫處理后殘留的無機物質,結果顯示泰國虎紋蛙的灰分含量顯著高于牛蛙。粗蛋白、粗脂肪水平是評價肌肉品質的重要指標,本研究中兩種蛙的粗脂肪含量遠低于15 種常見的經濟淡水魚類[15],泰國虎紋蛙相比于牛蛙脂肪含量偏低,這可能是由于兩者的規格不同。此外,與棘胸蛙[6]、黑斑蛙[10]和雙團棘胸蛙 (Paayunnanensis)[16]等養殖蛙類相比,泰國虎紋蛙和牛蛙的粗蛋白含量依然占據優勢。泰國虎紋蛙和牛蛙肌肉營養總體呈現高蛋白低脂肪的特點,符合現代人們對高品質膳食的營養需求。
3.2 肌肉氨基酸組成
泰國虎紋蛙和牛蛙氨基酸總量中必需氨基酸的占比分別為45.26%和44.75%,必需氨基酸與非必需氨基酸的比值分別為82.69%和81.13%,優于FAO/WHO 建議的理想模式40%和60%,因此在營養學上,泰國虎紋蛙和牛蛙肌肉均屬于優質蛋白質來源。此外,泰國虎紋蛙和牛蛙的第一限制性氨基酸是亮氨酸,第二限制性氨基酸為蛋氨酸。
泰國虎紋蛙和牛蛙肌肉中必需氨基酸含量最豐富的是異亮氨酸,其屬于人體必需氨基酸。異亮氨酸具有增強腸道黏膜免疫功能的作用[17],此外其通過調節機體對葡萄糖的攝取來調節腸道發育[18]。適量食用泰國虎紋蛙和牛蛙能夠大量補充異亮氨酸,具有一定的保健功效。泰國虎紋蛙中蘇氨酸含量顯著高于牛蛙,蘇氨酸是一種重要的營養強化劑,有緩解人體疲勞、促進生長發育的效果[19]。支鏈氨基酸具有促進蛋白質的合成、減緩衰老和防治肝腎功能衰竭的作用[20]。高支鏈氨基酸、低芳香氨基酸混合物對人體肝臟具有保護作用[21]。泰國虎紋蛙和牛蛙肌肉支鏈氨基酸與芳香氨基酸的比值基本一致,分別為2.20 和2.19,均屬于高支鏈氨基酸、低芳香氨基酸的食品。因此,食用泰國虎紋蛙和牛蛙肌肉可能有助于維持機體肝臟健康。肉類產品鮮美程度一般受呈味氨基酸含量和構成比例的影響,其中天冬氨酸、谷氨酸呈鮮味特性,甘氨酸、丙氨酸則呈甘味特性,且以谷氨酸的鮮味最強[22]。本研究中泰國虎紋蛙和牛蛙肌肉中含量最高的呈味氨基酸均為甘氨酸。泰國虎紋蛙和牛蛙肌肉中呈味氨基酸與總氨基酸的比值 (FAA/TAA) 較高,分別為43.71%和44.61%,這也是其味道鮮美的主要原因。
3.3 肌肉脂肪酸組成
泰國虎紋蛙和牛蛙肌肉中飽和脂肪酸以C16:0 居多,單不飽和脂肪酸以C18:1 居多,多不飽和脂肪酸以C18:2 居多。兩者飽和脂肪酸中含量最高的是C16:0,其次是C18:0;前者能使血清膽固醇降低,起到改善胰島素抵抗的作用[23],后者可以減少腸道膽固醇吸收進而使肝臟膽固醇含量降低[24]。不飽和脂肪酸具有降血脂、預防心血管疾病和促進生長發育的功能[25],此外,不飽和脂肪酸含量對于肉質風味的形成具有重要作用[26],牛蛙的不飽和脂肪酸含量顯著高于泰國虎紋蛙,說明其具有更好的風味和保健效果。單不飽和脂肪酸的適量攝入可以使低密度脂蛋白水平和冠心病發生率顯著降低[27]。C18:1 可降低血液總膽固醇含量,減少動脈粥樣硬化與心臟病的患病風險[28]。牛蛙的單不飽和脂肪酸和C18:1 含量顯著高于泰國虎紋蛙,擁有更好的心臟保健效果。多不飽和脂肪酸含量與肉質加熱過程中產生的香氣成正比[29],一定程度上體現了肌肉的多汁性;多不飽和脂肪酸具有控制血壓、血脂和抗腫瘤的作用[30],還具有延緩衰老等保健功效。泰國虎紋蛙的多不飽和脂肪酸含量顯著高于牛蛙,其肌肉口感更具多汁性。C22:6(DHA) 是人體必需脂肪酸,具有保護心肌及血管、治療癌癥、增強視力和健腦的功能[22]。牛蛙DHA 含量比泰國虎紋蛙更高,因此從DHA 含量角度來看,食用牛蛙的保健效果更佳。P/S 是衡量脂肪酸營養價值的重要指標,WHO 建議P/S 應高于0.4[31]。泰國虎紋蛙和牛蛙的P/S 介于1.49~1.53,超過一般的經濟魚類 (0.35~1.25)[32],說明泰國虎紋蛙和牛蛙是很好的脂肪酸營養食物源。可見,泰國虎紋蛙和牛蛙具有較高的脂肪酸營養價值,可對其營養和保健功效進行開發利用。
3.4 肌肉形態結構與質構特性
肌纖維是肌肉的基本組成單位,肌纖維直徑會影響肌肉的質地和結構參數,如硬度、彈性和咀嚼性等[33],且肌纖維直徑越大,肌肉硬度越高[34],而高彈性、高硬度的肌肉口感更佳[33]。肌肉形態學顯示牛蛙肌纖維直徑大于泰國虎紋蛙,這與肌肉質構特性的結果相一致。因此,從肌肉質構特性的角度看,牛蛙肌肉的食用口感更佳。
3.5 腸道形態結構和微生物多樣性
腸絨毛高度、寬度和肌層厚度常用于評價腸道形態健康[35]。腸道絨毛的形態結構對其消化吸收營養物質的能力有著直接影響[36],腸絨毛的高度和寬度影響營養物質消化的速度;而食糜主要通過腸道肌層的收縮進行機械消化,因而腸肌層厚度的增加有利于機體對食糜的充分消化[37]。牛蛙腸絨毛高度和寬度均大于泰國虎紋蛙,表明牛蛙腸道消化吸收能力更優;而泰國虎紋蛙腸肌層厚度顯著高于牛蛙,說明其對食糜的機械消化能力更勝一籌。
腸道微生物群落參與宿主的消化、代謝調節、免疫系統調控等過程。劉文舒等[12]發現虎紋蛙腸道優勢菌門為厚壁菌門、變形菌門、擬桿菌門和梭桿菌門,與本研究結果相似。邵闖等[38]研究發現牛蛙腸道優勢菌門為解糖微小寄生菌門、放線菌門及變形菌門,與本研究結果存在差異。以全豆粕蛋白為飼料來源的牛蛙,其腸道優勢菌門為變形菌門、梭桿菌門和厚壁菌門[39],與本研究結果相似。因此,不同飼料可能會影響牛蛙腸道的優勢菌。江昀等[40]推測蛙類腸道固有菌群為厚壁菌門、擬桿菌門和變形菌門,基因、年齡、性別以及養殖環境等多種因素會影響其組成比例。泰國虎紋蛙和牛蛙的厚壁菌門和放線菌門豐度差異顯著,可能是由于兩者基因不同對腸道優勢菌產生了影響。
益生菌對維持腸道健康有重要作用,作為添加劑可以提高水產動物的生長率,增強其免疫力和抗病性,但是非宿主來源的益生菌在使用過程中存在潛在風險,宿主來源的益生菌更值得關注和挖掘[41]。乳球菌屬[42]、乳桿菌屬[43]、腸球菌屬[44]、魏斯氏菌屬[45]和明串珠菌屬[46]是泰國虎紋蛙和牛蛙腸道菌群的優勢菌屬,均被證實是水產動物的潛在益生菌,對營養吸收、生長發育和抑制有害菌有積極作用;泰國虎紋蛙的乳球菌屬豐度顯著高于牛蛙,而乳桿菌屬和明串珠菌屬豐度則顯著低于牛蛙,表明兩者潛在的益生菌比例構成存在種間差異。鯨桿菌屬能提高氨基酸轉運和代謝率[47],泰國虎紋蛙中該菌屬相對豐度最高,這與功能預測中氨基酸代謝的功能基因豐度的結果相一致。泰國虎紋蛙和牛蛙腸道菌群功能基因中占比較高的均為與新陳代謝相關的功能基因,這有利于提高宿主的生長發育、營養物質轉運及代謝性能。
4 結論
泰國虎紋蛙和牛蛙肉質總體呈現高蛋白、低脂肪的特點,具有高質量的必需氨基酸組成以及豐富的不飽和脂肪酸和必需脂肪酸;牛蛙的FAA/TAA和肌肉質構特性優于泰國虎紋蛙,具有更優質的風味和口感。高通量測序顯示,泰國虎紋蛙的腸道菌群Alpha 多樣性更高;乳球菌屬在泰國虎紋蛙腸道中顯著富集,而乳桿菌屬和明串珠菌屬在牛蛙腸道中顯著富集;功能預測顯示兩者腸道菌群可能主要參與宿主營養物質轉運及代謝等相關進程。泰國虎紋蛙和牛蛙是平衡膳食營養結構的優質來源,具有良好的開發利用與人工養殖前景。
