奇孢根霉強化中溫大曲對糟醅發酵過程的影響
2024-01-19 09:20:26萬營東唐秋香周榮清張宿義李德林王小軍
中國釀造 2023年12期
萬營東,唐秋香,黃 鈞,周榮清,2*,張宿義,秦 輝,董 異,王 超,張 程,李德林,王小軍,張 柱
(1.四川大學 輕工科學與工程學院,四川 成都 610056;2.國家固態釀造工程技術研究中心,四川 瀘州 646699;3.瀘州老窖股份有限公司,四川 瀘州 646699)
中國白酒是世界六大蒸餾酒之一,具有悠久的歷史[1]。濃香型白酒是中國三大傳統白酒之一,其生產工藝獨樹一幟[2-3]。濃香型白酒的發酵生產環節主要包括大曲制作和糟醅發酵過程[4]。大曲的菌系、酶系與白酒的產量和風格密切相關[5]。篩選功能菌株并應用于強化大曲,不僅可以提高大曲的酶活力,還能改善白酒的特征風味。研究表明,將Bacillus用于強化中溫大曲,顯著提高了其糖化力和液化力,并且改變了與糖化活性和液化活性相關的微生物群落組成,從乳桿菌屬和根毛霉屬變為芽孢桿菌屬、魏斯氏菌屬和絲孢畢赤酵母屬[6]。此外,HONG L等[7]研究發現,使用功能性酵母強化大曲,代替傳統大曲模擬特香型白酒發酵,提高了糟醅的酒精度和揮發性風味物質的含量,同時并未改變優勢細菌屬的相對豐度。
根霉(Rhizopus)是釀酒曲中的主要功能菌之一[8-9],其主要特點是水解酶譜廣,如分泌α-淀粉酶、糖淀粉酶、糖苷酶、糖基轉移酶等[10],且具有產酒精能力。根霉的高糖化力使其在發酵工業中得到廣泛應用[9]。黃艷等[11]從酒曲中篩選出根霉,應用于米醋生產,成功地提升了米醋的風味和品質。此外,麥麩通過接種米根霉進行固態發酵后,不僅降低了不愉悅風味物質的相對含量,還提高了多種芳香揮發性成分的相對含量,從而改善了其感官品質和營養價值[12]。黃科屹等[13]的研究表明,從中高溫大曲中分離的米根霉可以制成米曲,添加至白酒釀造中,顯著提高了出酒率,說明出酒率與大曲體系的糖化能力呈正相關。在之前的研究中,TANG Q等[14]利用源于優質小曲的奇孢根霉(Rhizopus azygosporus)強化小曲發酵,提高了小曲的淀粉水解力和酯化活力,并改善了小曲糟醅的理化性質和風味特點。馬鵬[15]通過混合根霉、米曲霉和紅曲霉制備米曲,與傳統大曲混合用于白酒固態發酵,提升了白酒的酒精度,豐富了白酒的風味物質。
總的來說,目前的研究主要集中在小曲和米曲中強化根霉的應用,以及將米曲與傳統大曲混合用于糟醅發酵,重點關注了白酒的理化和風味改善。然而,對于在大曲制作過程引入根霉對大曲和發酵糟醅的影響規律的相關研究還相對較少。需要注意的是,大曲真菌屬相對豐度可能受到微生物間競爭的影響,在高溫大曲中,更耐熱的嗜熱真菌屬和紅曲霉屬競爭優勢更強,而根霉則生長緩慢[16]。因此,在中溫大曲中引入根霉可能更為合適,且中溫大曲具有較高的糖化力,與根霉的特點相輔相成。
本研究以根霉強化的中溫大曲為發酵劑,通過模擬窖池發酵,多角度地探討了根霉強化對糟醅微生物群落結構和揮發性風味成分影響的規律,旨在調控白酒釀造周期和改善風味成分,為開發固態白酒生產新工藝奠定理論基礎。
1 材料與方法
1.1 材料與試劑
奇孢根霉(Rhizopus azygosporus)CGMCC 20728:由本實驗室篩選自中國典型小曲酒廠,菌株保存于本實驗室;Fast DNA SPIN提取試劑盒:美國MP Biomedicals公司;Q5脫氧核糖核酸(deoxyribonucleic acid,DNA)高保真聚合酶:美國New England Biolabs公司;瓊脂糖、瓊脂糖凝膠電泳緩沖液、Quant-iT PicoGreen dsDNA Assay Kit:美國Invitrogen公司;VAHTSTM DNA Clean Beads:南京Vazyme公司;其他化學試劑(分析純):成都金山化學試劑有限公司。
1.2 儀器與設備
Trace 1300-TSQ 9000氣相色譜-質譜聯用儀:美國Thermo Fisher Electron公司;VF-WAX-MS毛細管色譜柱(30 m×0.25 mm×0.25 μm):美國Bellefonte公司;NanoDrop ND-1000分光光度計:美國Thermo Fisher Scientific公司;GL-20G-II立式高速冷凍離心機:上海安亭科學儀器有限公司;S100 TM Thermal Cycler聚合酶鏈式反應(polymerase chain reaction,PCR)儀、Gel Doc TM XR凝膠成像儀:美國Bio-Rad公司;85-2型數顯恒溫磁力攪拌器:上海雙捷實驗設備有限公司。
1.3 方法
1.3.1 樣品制備與收集
大曲樣品取自瀘州著名釀酒企業制曲中心,參照ZHENG X等[17]所述的工藝過程及參數控制生產,根霉大曲(RQ)接種0.05%的Rhizopus azygosporusCGMCC 20728,普通大曲(NQ)作為對照不接種根霉。
模擬發酵實驗采用改良的堆積糖化發酵工藝,發酵時間28 d。將RQ和NQ添加到蒸熟的高粱中(比例為干質量的15%),堆積糖化24 h,裝入模擬發酵池(6 L塑料容器,內徑為25 cm×17 cm×14 cm),在30 ℃條件下發酵28 d,相應的糟醅記為根霉糟醅(RZP)和普通糟醅(NZP)。兩組樣品各設置3個平行,在發酵的0 d、7 d、14 d、21 d和28 d采用五點法取樣,每個平行樣品在取樣后均被分為兩份,一份置于-20 ℃保藏用于理化性質和揮發性風味物質檢測,另一份置于-80 ℃保藏用于微生物群落結構分析。
1.3.2 理化性質測定
大曲樣品的水分含量、酸度、酯化力、糖化力、液化力和發酵力的測定:參照QB/T 4257—2011《釀酒大曲通用分析方法》所述方法;糟醅還原糖(reducing sugar,RS)含量、淀粉(starch)含量、酸度(total acidity,TA)、酒精度的測定:參照文獻[18]所述方法檢測。
1.3.3 揮發性風味物質的檢測
揮發性物質通過頂空固相微萃取結合氣相色譜-質譜法(headspace solid-phase microextraction gas chromatographymass spectrometry,HS-SPME-GC-MS)測定。
樣品的預處理[19]:將1.00 g樣品和10 μL內標物質(辛酸甲酯,0.007 3 g/100 mL)加入20 mL頂空瓶中,將頂空瓶置于恒溫磁力攪拌器中60 ℃平衡15 min,使用50/30 μm DVB/CAR/PDMS 纖維萃取吸附揮發性組分45 min。萃取頭在250 ℃解吸5 min。
氣相色譜條件:進樣口溫度250℃,載氣為流速1mL/min的高純氦氣(He)(>99.999%),不分流模式;升溫程序:初始溫度40 ℃,維持5 min,以4 ℃/min升溫至100 ℃,以6 ℃/min升溫至230 ℃,維持10 min。
質譜條件:離子源溫度250 ℃,傳輸線溫度300 ℃,電離方式為電子電離(electronic ionization,EI)源,電子能量70 eV,掃描范圍35~400 amu。
定性定量分析:檢測所得質譜圖與美國國家標準與技術研究院(national institute of standards and technology,NIST)2017質譜庫對比后,僅保留相似度>80%化合物進行進一步分析;根據內標辛酸甲酯的含量與其峰面積的比值,計算出各揮發性化合物的含量。
1.3.4 微生物群落多樣性檢測
依據FastDNASPIN提取試劑盒的操作說明從樣品中提取總DNA。同時采用0.8%瓊脂糖凝膠電泳檢測DNA提取質量和用分光光度計檢測DNA含量。用通用引物338F/806R和ITS5/ITS1分別擴增細菌16S rRNA V3-V4區和真菌ITS1區。
聚合酶鏈式反應(PCR)擴增體系:5 μL 5×Q5高保真反應緩沖液,5 μL 5×Q5高保真氣相色譜緩沖液,0.25 μL Q5高保真DNA聚合酶(5 U/μL),2 μL脫氧核糖核苷三磷酸(deoxyribonucleoside triphosphates,dNTPs)(2.5 mmol/L),正反向引物各1 μL(10 μmol/L),2 μL DNA模板和8.75 μL ddH2O。PCR擴增條件:98 ℃預熱2 min,之后25個循環(包括98 ℃變性15 s,55 ℃退火30 s,72 ℃延伸30 s),最后72 ℃延伸5 min。PCR產物純化:用VAHTSTMDNA Clean Beads純化擴增產物,用Quant-iT PicoGreen dsDNA Assay Kit定量。樣品純化后,委托上海派森諾生物科技有限公司在Illumina Novaseq平臺完成DNA片段雙端(2×300 bp)測序。
原始測序數據主要通過QIIME2處理。使用demux插件對原始序列進行多路分解,使用cutadapt去除引物,接著使用DADA2對序列進行質量過濾、去噪、合并和嵌合體移除。進一步篩選僅在一個樣本和單體中發現的擴增子序列(amplicon sequence variants,ASV)。最后,根據Silva(v132)和UNITE(v8.0)數據庫進行比對。
1.3.5 數據統計分析
使用OriginPro 2019b對理化性質進行可視化。在派森諾基因云平臺(https://www.genescloud.cn/home)進行以下分析:分析微生物群落的α多樣性、β多樣性,包括Chao1指數、Shannon指數等,并用線性判別分析效應大小(linear discriminant analysis effect size,LEfSe)分析(LDA>4,P<0.05)確定微生物群落的標志微生物。基于KEGG數據庫和Metacyc數據庫,利用PICRUSt2預測代謝功能。在SIMCA14.1中進行偏最小二乘-判別分析(partial least squares-discriminant analysis,PLS-DA)和排列檢驗。使用TB Tools(v1.108)生成熱圖。在圖圖云平臺(https://www.cloudtutu.com/#/index)和OmicShare Tools(https://www.omicshare.com/tools)分別執行冗余分析(redundancy analysis,RDA)和趨勢分析。
2 結果與分析
2.1 理化性質變化
水分、酸度及水解能力是衡量大曲質量的主要參數[20],大曲的理化性質差異見表1。由表1可知,RQ的水分、酸度和酯化力略低于NQ,糖化力、液化力、發酵力高于NQ,除酯化力外的其余相關參數都因接種根霉得到改善。將兩種大曲用于發酵糟醅,糟醅間理化參數的變化見圖1。由圖1可知,堆積糖化階段結束時,RZP和NZP的淀粉含量分別是31.53%和34.42%,前者的水解率高于后者,發酵結束淀粉含量分別是25.07%和27.51%。發酵前中期(0~14 d)淀粉被水解,后期因代謝產物(總酸、乙醇)的抑制及淀粉底物含量的降低,降解幅度減弱。還原糖含量和酸度在RZP和NZP中均是逐漸增高的,且RZP的增幅大于NZP,這可能是因為RQ具有更高的糖化力和液化力,使RZP的底物更有效地水解為糖,同時糖發酵成酸。酒精度在前期增幅較大,在發酵14 d時,RZP的酒精度(8.57%vol)顯著高于NZP(7.28%vol)(P<0.05),發酵結束時,RZP的酒精度(7.70%vol)僅略高于NZP(7.20%vol)。以上結果表明,RZP和NZP理化參數的變化趨勢一致,但根霉強化降低了糟醅的淀粉含量,同時提高了糟醅的酸度、還原糖含量和酒精度。

圖1 糟醅理化性質的變化趨勢Fig.1 Change trends in physicochemical properties of fermented grains

表1 兩種大曲的理化性質測定結果Table 1 Determination results of physicochemical properties of two kinds of Daqu
2.2 揮發性風味成分的差異
2.2.1 揮發性成分含量差異
糟醅樣品的揮發性風味成分測定結果見圖2。由圖2可知,兩種樣品中共檢出了69種揮發性組分,包括酯類(45種)、醇類(4種)、酸類(1種)、烷類(2種)、烯類(3種)、酮類(3種)、酚類(1種)、醛類(6種)和其他類(2種)組分。

圖2 糟醅揮發性風味成分含量差異及共有揮發性成分聚類分析熱圖Fig.2 Difference of volatile flavor component content and clustering analysis heatmap of shared volatile components in fermented grains
由圖2A可知,揮發性風味成分總量隨發酵時間增加而逐漸升高,且在發酵的初始階段(0~7 d)和中后期(14~21 d)增量明顯,RZP的揮發性風味成分總量增量分別是20.15 mg/kg和19.86 mg/kg,較同期NZP的總量增量高39.07%和37.05%。在發酵后期(21~28 d),RZP揮發性風味成分總量趨于穩定,NZP卻降低,發酵結束時,RZP揮發性風味成分總量為59.52 mg/kg,NZP僅為28.11 mg/kg。酯類組分占總揮發性風味成分含量的74.23%~88.68%,經7 d發酵,酯類含量從74.23%~74.34%增至86.52%~87.81%,后趨于穩定。發酵結束時,RZP酯類化合物的含量為51.22 mg/kg,NZP僅為23.88 mg/kg。酯類中主要是賦予基酒果香、花香和甜味的乙酯類[21],作為骨架成分的己酸乙酯和乙酸乙酯[22]含量經發酵顯著增高(P<0.05),且根霉強化也使它們的含量增加,發酵結束時,RZP中二者的含量比NZP分別增加了71.22%和44.36%。此外,糟醅中棕櫚酸乙酯的含量也因根霉強化而增加,發酵結束時,其在RZP中的含量比NZP中增加了93.66%。棕櫚酸乙酯呈微弱蠟香、果香和奶油香氣,能賦予酒體醇厚感、降低干澀味[23]。發酵后棕櫚酸甲酯等成分的比例減少。醇類物質賦予酒體醇厚綿甜風味[24-25],在0 d時比例最高,為7.88%~19.39%,且變化趨勢與酯類相反,可能與參與酯類合成有關,發酵結束時,RZP醇類化合物的含量為4.75 mg/kg,NZP為2.45 mg/kg。RZP和NZP中異戊醇的比例分別從初期的10.80%和12.45%降至結束時的3.21%和3.46%。
由圖2B可知,檢出的揮發性代謝組分中有22種是共有的,包括酯類17種、醇類3種、酚類1種和醛類1種。這些成分的聚類分析的結果見圖2C。由圖2C可知,揮發性風味物質可為兩個簇,簇Ⅰ的成分包括酯類2種和酚類1種,而簇Ⅱ則包括酯類15種、醇類3種和醛類1種。這些成分在RZP中的含量都高于NZP,在發酵過程中,簇I的呈相對穩態,而簇II的則逐漸增高,且在21 d時增幅最大。
2.2.2 特征揮發性成分分析
通過偏最小二乘判別分析(PLS-DA)法探討RZP和NZP間的差異性,結果見圖3。

圖3 糟醅揮發性風味成分的偏最小二乘判別分析結果及熱圖Fig.3 Partial least squares discriminant analysis results and heat map of volatile flavor components in fermented grains
由圖3A可知,RZP和NZP被明顯分離,而R2X(模型在X空間的擬合度)、R2Y(模型在Y空間的擬合度)、Q2(模型預測能力)分別為0.872、0.919、0.855,表明構建的模型可靠性好。由圖3B可知,200次排列檢驗證明PLS-DA模型無過擬合,適合篩選樣品間特征揮發性代謝物。基于預測變量重要性投影(variable importance in the projection,VIP)值(VIP>1.0)的結果,確定了12種成分,包括10種酯、1種醇和1種酚,其中9種是共有的。這些成分隨發酵的變化如圖3C所示,大部分乙酯類成分(棕櫚酸乙酯、己酸乙酯、乙酸乙酯、反油酸乙酯、琥珀酸二乙酯、十七酸乙酯)的含量經發酵均有明顯的增加,且在14~21 d增幅最高,而棕櫚酸甲酯、辛酸乙酯、異戊醇和4-乙基愈創木酚在發酵過程中呈相對穩態,癸酸乙酯和亞油酸甲酯在NZP中前中期含量較高而后期并未檢測到。絕大部分成分在RZP的含量高于NZP,表明根霉強化可以提高糟醅中特征揮發性組分的含量。
2.2.3 揮發性風味成分表達趨勢分析
研究風味代謝積累表達的時間趨勢,對發酵過程的揮發性成分含量進行趨勢分析,并將相關系數>0.7的趨勢聚為一類。在RZP和NZP中各觀察到了2種趨勢顯著聚類,結果見圖4。由圖4可知,RZP和NZP分別有79.48%和77.09%的揮發性風味成分呈漸增趨勢。其中,聚類1代表揮發性風味成分在發酵過程中含量一直增加,聚類2代表揮發性成分含量在21 d之前呈漸增趨勢,21 d之后略降。值得注意的是,RZP和NZP分別有46.15%和66.67%的揮發性成分在發酵21 d之前含量增加,而在21 d之后含量略降,揭示了21 d是糟醅揮發性成分變化的關鍵時間節點。

圖4 發酵過程中揮發性風味成分的趨勢分析Fig.4 Trend analysis of volatile flavor components in fermentation process
2.3 微生物多樣性分析
2.3.1 微生物群落Alpha多樣性指數差異
微生物的Alpha多樣性差異通常可以由豐富度(Chao1指數、觀察物種指數)和多樣性(香農指數、辛普森指數)解釋。揮發性代謝成分的差異分析的結果表明,21 d是發酵系統揮發性成分變化的關鍵時間節點,因此僅對大曲和發酵21 d糟醅的微生物群落結構進行分析。群落的Alpha多樣性指數的差異見表2。

表2 大曲和發酵21 d糟醅的Alpha多樣性指數Table 2 Alpha diversity indexes of Daqu and fermented grains fermented for 21 d
由表2可知,大曲和對應的發酵糟醅群落的Alpha多樣性呈現一致的趨勢。對于真菌群落而言,RQ的豐富度低于NQ,但多樣性高于NQ,同時RZP的豐富度也低于NZP。這可能是因為根霉強化提高了RQ中根霉屬(Rhizopus)的豐度,亦改變了菌群結構,尤其降低了真菌群落的豐富度。
對于細菌群落而言,RQ的豐富度和多樣性均高于NQ,同樣地,RZP的豐富度和多樣性也高于NZP。根霉強化提高了水解底物的速率,導致在大曲制作前期和糟醅堆積糖化階段,積累了豐富的營養物質,加快了細菌定植速率[14],從而使RQ和RZP細菌群落豐富度都高于NQ和NZP。
2.3.2 微生物群落組成差異
大曲和糟醅的微生物群落組成見圖5。由圖5A可知,大曲和糟醅真菌群落由6個門構成,主要為毛霉門(Mucoromycota)和子囊菌門(Ascomycota),相對豐度之和為88.08%~99.49%。Mucoromycota在RQ和NQ中的相對豐度分別為27.40%和4.73%,而在RZP和NZP中分別增至57.64%和39.54%。相反地,Ascomycota在RQ和NQ中的相對豐度分別為72.09%和93.31%,而在RZP和NZP中分別降至30.44%和53.26%。屬水平的優勢真菌如圖5B所示,大曲的優勢真菌屬包括根霉屬(Rhizopus)、嗜熱真菌屬(Thermomyces)和嗜熱子囊菌屬(Thermoascus),其中Rhizopus是常見真菌之一,分泌多種水解酶,具有糖化和發酵且產生乙酸乙酯等代謝成分的能力[26]。RQ中Rhizopus的相對豐度是27.35%,而NQ僅為3.86%。值得關注的是根霉強化還顯著提高了大曲中Thermomyces的相對豐度(48.06%),但降低了Thermoascus的相對豐度(22.92%)。這兩個真菌屬在NQ中的相對豐度分別是14.64%和69.47%。Thermoascus亦能分泌多種酶,如過氧化氫酶、內切葡聚糖酶、葡萄糖苷酶等[27],且Thermomyces和Thermoascus皆具有較強的耐熱性,在發酵過程中能保持穩定的催化效率[28]。發酵21 d時,RZP的優勢真菌屬分別是Rhizopus(57.32%)、Thermomyces(3.22%)和酵母屬(Saccharomyces)(1.17%),NZP的優勢真菌屬則是Rhizopus(32.16%)、橫梗霉屬(Lichtheimia)(7.30%)、Pichia(2.26%)和Saccharomyces(1.37%)。由于根霉強化提高了Rhizopus的相對豐度,同時也占據了其余真菌的生存空間,導致真菌群落的豐富度降低。

圖5 大曲和糟醅微生物群落組成差異Fig.5 Composition difference of microbial community of Daqu and fermented grains
由圖5C可知,大曲和糟醅的細菌群落由12個門組成,主要為厚壁菌門(Firmicutes)和變形菌門(Proteobacteria),其相對豐度之和為99.18%~99.88%。大曲中Firmicutes的相對豐度最高,在RQ和NQ中的相對豐度分別為93.58%和98.94%。Firmicutes在RZP和NZP中其相對豐度降至50.30%和44.89%,而Proteobacteria的相對豐度分別增至49.58%和54.96%。兩種大曲的優勢細菌屬組成僅略有差異,但相對豐度差異明顯(圖5D)。乳桿菌屬(Lactobacillus)、克羅彭斯特菌屬(Kroppenstedtia)和魏斯氏菌屬(Weissella)是共享的優勢屬,Lactobacillus和Weissella在RQ中的相對豐度(29.43%和25.53%)高于NQ(10.20%和11.37%),而NQ中Kroppenstedtia(76.68%)顯著高于RQ(22.86%)。高溫放線菌屬(Thermoactionmyces)(13.02%)也是RQ的優勢菌之一。發酵21 d時,RZP的優勢細菌屬包括片球菌屬(Pediococcus)(28.17%)、腸桿菌屬(Enterobacter)(10.46%)、Lactobacillus(10.31%)、雷爾氏菌屬(Ralstonia)(22.91%)等8個屬,NZP則僅包括Pediococcus(38.45%)、Ralstonia(37.39%)、鞘氨醇單胞菌屬(Sphingomonas)(10.21%)等6個屬。Lactobacillus、Pediococcus、Weissella等乳酸菌的主要代謝產物包括乙酸、乳酸等多種有機酸,這些代謝產物不僅是白酒的呈味成分,也是乙酸乙酯、乳酸乙酯等特征代謝物的前體[29-30]。
根霉強化顯著改變了大曲和糟醅的群落結構。它增加了大曲中Rhizopus、Thermomyces、Lactobacillus、Weissella和Thermoactionmyces的相對豐度,降低了Thermomyces和Kroppenstedtia的相對豐度。對糟醅而言,根霉強化增加了Rhizopus、Thermomyces等的相對豐度,同時降低了Lichtheimia、Pichia等的相對豐度,此外,根霉強化還增加了糟醅細菌優勢屬的數量,并提高了Enterobacter、Lactobacillus等的相對豐度。
2.3.3 特征微生物分析
通過線性判別分析效應大小(LEfSe)(LDA>4,P<0.05)法,識別了21 d時RZP和NZP之間的特征微生物,結果見圖6。由圖6A可知,RZP在綱、目、科、屬水平上鑒定出特征真菌,NZP在門、綱、目、科、屬等五個分類水平上均鑒定出特征真菌。在屬水平上,Thermomyces是RZP的特征真菌,NZP的則是Pichia和Lichtheimia。由圖6B可知,RZP在界、目、科、屬水平上鑒定出特征細菌,且RZP的特征細菌屬是Thermoactinomyces、Lactobacillus和Enterobacter,而在各分類水平上均未識別出NZP的特征細菌。

圖6 微生物群落的線性判別分析效應大小結果Fig.6 Linear discriminant analysis effect size analysis results of microbial communities
2.3.4 綜合代謝途徑分析
對涉及糟醅原料利用、代謝產物生成以及酶的表達的代謝途徑進行分析,結果見圖7。由圖7可知,在RZP中,由于根霉的強化,提高了Rhizopus的相對豐度,葡糖淀粉酶、糖原磷酸化酶、內切葡聚糖酶的表達顯著上調(P<0.05),提高了淀粉原料的代謝速率,降低了RZP的淀粉含量,所以RZP中的RS含量高于NZP。糖酵解途徑中乙醇代謝的關鍵酶顯著下調(P<0.05),包括丙酮酸脫羧酶、乙醇脫氫酶,解釋了RZP在發酵后期逐漸降低的乙醇含量。甲酸C-乙酰轉移酶的顯著上調(P<0.05)和乙酰輔酶A合成酶的顯著下調(P<0.05)導致RZP中乙酸鹽的積累增加。

圖7 涉及原料利用、代謝物生成和酶的代謝途徑分析Fig.7 Metabolic pathway analysis involving raw material utilization,metabolite production and enzymes
2.4 微生物群落與揮發性組分的相關性
通過對應分析與多元回歸分析相結合的冗余分析(RDA)揭示酒醅中優勢微生物屬與重要揮發性成分的相關性,結果見圖8。由圖8可知,采用Hellinger距離方法獲得了19種揮發性組分與5個優勢真菌屬和5個優勢細菌屬的相關性。Rhizopus、Lactobacillus和棕櫚酸乙酯、反油酸乙酯、丁酸乙酯、9-十六碳烯酸乙酯、乙酸乙酯、苯乙醇等呈正相關,這可能是因為乳酸、乙酸等多種有機酸是Lactobacillus等乳酸菌的主要代謝產物,而Rhizopus具有較強的酯化作用[31],能促進前體物質形成酯類,因此RZP中高相對豐度的Rhizopus和Lactobacillus協同作用提高了棕櫚酸乙酯、反油酸乙酯、丁酸乙酯、9-十六碳烯酸乙酯、苯乙醇等成分的含量。由此可見,根霉強化有助于增加糟醅中優勢揮發性組分的含量,提升其風味和香氣。Thermomyces、Enterobacter和9-十六碳烯酸乙酯、乙酸乙酯、苯乙醇、肉豆蔻酸乙酯、己酸乙酯、γ-亞麻酸甲酯、異戊醇、辛酸乙酯等呈正相關,Ralstonia、Sphingomonas、Saccharomyces和γ-亞麻酸甲酯、己酸乙酯、辛酸乙酯、棕櫚酸甲酯、乳酸乙酯、亞油酸甲酯、癸酸乙酯等正相關,Pichia、Lichtheimia、Pediococcus和苯乙酸乙酯、4-乙基愈創木酚、月桂酸乙酯、琥珀酸二乙酯、癸酸乙酯、亞油酸甲酯、反油酸乙酯、丁酸乙酯等呈正相關。根霉強化通過提高RZP中Rhizopus、Lactobacillus、Thermomyces、Enterobacter等的豐度,使與這些優勢微生物屬相關的風味代謝物含量增加,提升糟醅的風味和香氣。
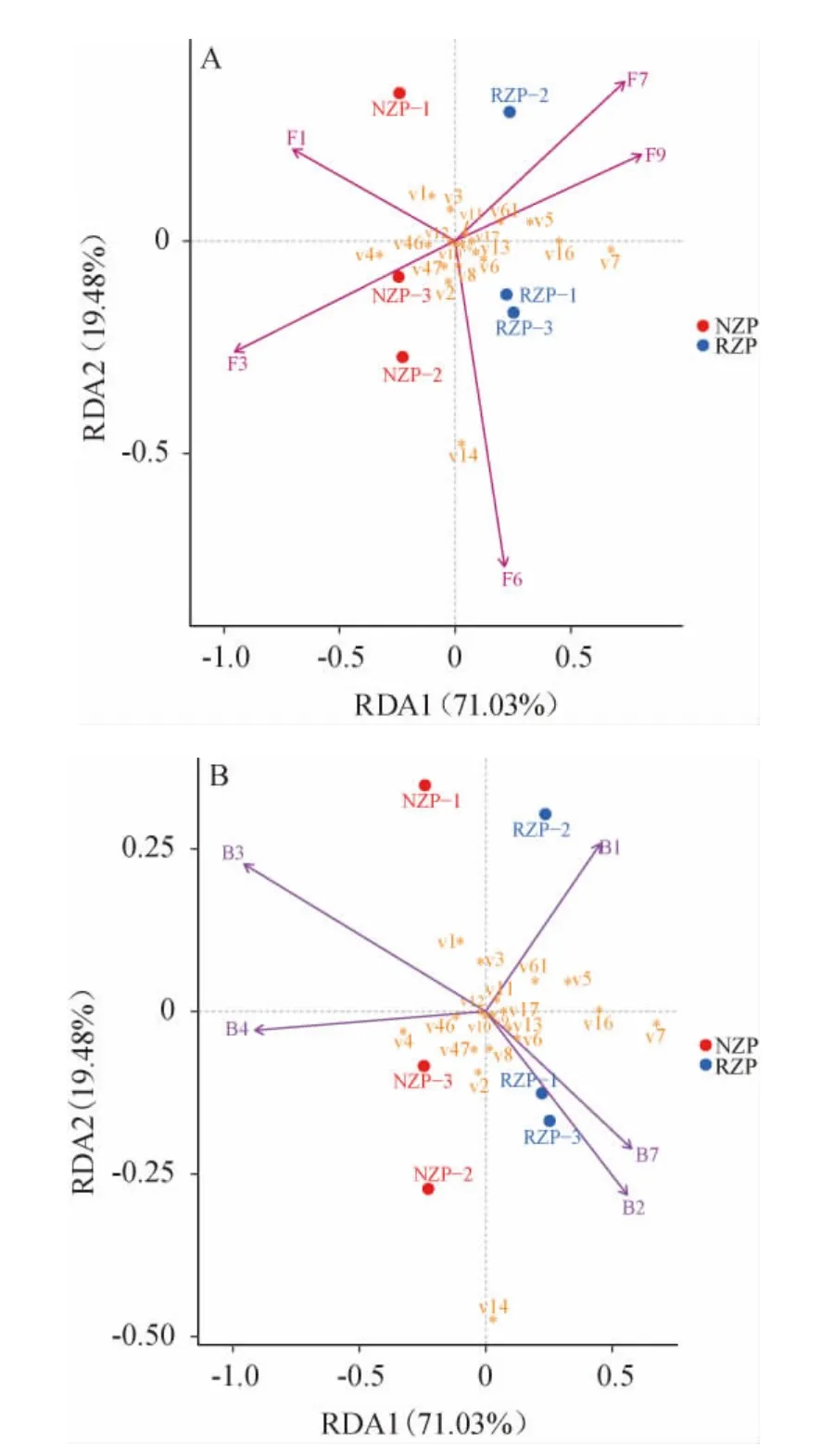
圖8 優勢微生物與主要揮發性化合物的冗余分析Fig.8 Redundancy analysis of dominant microorganisms and main volatile compounds
3 結論
本研究采用HS-SPME-GC-MS和高通量測序技術揭示了根霉強化對糟醅揮發性風味成分和微生物群落結構的影響規律。根霉強化顯著改變大曲和糟醅微生物群落結構,其顯著提高了RQ和RZP中Rhizopus、Thermomyces的相對豐度。Thermomyces、Thermoactinomyces、Lactobacillus、Enterobacter是RZP的標志微生物,Pichia、Lichtheimia則是NZP的標志微生物。糟醅中Rhizopus和大多數優勢揮發性成分呈正相關,根霉強化提高了糟醅中優勢揮發性成分的含量。發酵過程提高了乙酸乙酯和己酸乙酯的含量,并增大了酯類組分的比例,降低了醇類的比例。兩種糟醅間導致差異風味成分共12種,且21 d是系統代謝變化的一個關鍵時間點。研究結果為調節微生物群落結構和調控風味物質奠定了方法學和理論的基礎。
