番茄苗期耐低溫性主基因-多基因聯合遺傳分析
2024-01-08 09:32:00蘇百童閻世江
中國瓜菜 2023年12期
蘇百童,閻世江
(1.許昌職業技術學院 河南許昌 461000; 2.山西農業大學園藝學院 太原 030031)
番茄(Lycopersicon esculentumMill.)起源于美洲,傳入我國之后,種植面積日益擴大,尤其在我國北方設施栽培中面積居第一位,但在設施栽培的過程中如遇低溫逆境,對其生長發育、品質形成有較大的影響[1]。因此,培育耐低溫性較強的番茄專用品種成為學者的研究熱點。為加快選育過程,首先應明確番茄耐低溫性的遺傳規律。國內外研究者已有報道,Vos 等[2-3]研究在低溫下番茄種子的發芽能力、花粉萌發能力的遺傳,認為環境影響較大;Scott 等[4]研究在低溫下番茄坐果能力的遺傳規律;Femandez 等[5]研究在低溫下番茄果實發育速度的遺傳規律,結果顯示控制坐果能力、果實發育速度的性狀屬于數量性狀,其遺傳規律符合加性-顯性模型。林多等[6]給予番茄幼苗19 ℃/10 ℃的低溫處理,研究幼苗生長的遺傳規律,研究認為生長速度符合加性-顯性模型,該性狀的遺傳屬于不完全顯性,加性效應較強,狹義遺傳力達93.98%。上述學者在研究時采用的均是經典的Hayman 方法,可以從總體上估算加性效應、顯性效應,無法估算單一基因的遺傳,有一定的局限性。近年來,蓋鈞鎰等[7]經過深入的研究總結出一套全新的數量性狀遺傳分析方法,即主基因+多基因聯合遺傳模型分析法。禹山林等[8]對花生種子含油量,韓柱強等[9]對花生油酸、亞油酸含量,馬雪霞等[10]對亞洲棉纖維品質和產量性狀,閆世江等[11-12]對黃瓜在低溫下的生長速度、MDA 含量的遺傳研究中均采用這一方法,取得了良好的效果。在對番茄的耐低溫性相關研究中耐低溫性分級能直觀描述耐低溫性強弱[13-14],而運用主基因-多基因聯合遺傳分析方法研究番茄耐低溫性的報道較少見。為此,許昌職業技術學院蔬菜育種課題組通過前期的大量研究,對收集到的大量種質資源進行鑒定,從中篩選出耐低溫性不同的2 份番茄材料,均是經8 代以上自交的材料,然后構建P1、P2、BC1等群體,采用蓋鈞鎰等[7]的方法研究番茄耐低溫性的遺傳規律及遺傳參數,以期為培育番茄專用品種奠定基礎。
1 材料與方法
1.1 材料
番茄親本選擇XC-1(代號P1)、XC-2(代號P2),由許昌職業技術學院提供,其中XC-1 屬于大果型,表皮紅色,耐低溫性較強,屬無限生長型,是由瑞星大寶經8 代以上選育而成的自交系;XC-2 屬于小果型,表皮紅色,耐低溫性較弱,屬無限生長型,是由葡萄番茄經8 代以上選育而成的自交系。
1.2 方法
2021 年3 月1 日將P1、P2在許昌職業技術學院科研基地塑料大棚內進行播種,采用穴盤育苗法,35 d 后定植于大棚,于花期配制F1并留種,2021 年7 月5 日,將P1、P2、F1的種子在科研基地塑料大棚內播種,采用相同的育苗方法,35 d 后定植于大棚,同年于花期配制BC1(F1×P1)、BC2(F1×P2)、F2,并留種。2022 年3 月將上述6 個世代番茄材料播種于科研基地塑料大棚內,P1、P2、F1各播種200 粒,BC1、BC2、F2等分離世代各播種1000 粒,35 d 后放入人工氣候箱內模擬低溫環境,對幼苗進行低溫處理,日間溫度14 ℃,光照處理9 h(早8:00 至晚5:00),光照度為30μmol·m-2·s-1,夜間7 ℃(晚5:00 至早8:00),不進行光照,相對濕度為75%,共處理15 d。處理前與處理后照片詳見圖1~2。然后調查幼苗耐低溫性。耐低溫性采用分級數表示。0 級:植株瀕臨死亡;1 級:全部葉片萎蔫,心葉嚴重失水;2 級:全部葉片出現脫水斑,心葉略失水;3 級:2 片葉葉緣黃化,1 片葉無受害癥狀;4 級:2 片葉葉緣失水,1 片葉無明顯癥狀;5 級:無明顯癥狀。不同耐低溫性分級詳見圖3。調查樣本數:P1、P2、F1各調查40 株,BC1、BC2各調查120 株,F2調查150 株。采用隨機區組設計,3 次重復。

圖2 處理后Fig.2 After treatment

圖3 耐低溫性分級(0~5 級)Fig.3 Low temperature resistance level(0~5)
1.3 數據分析方法
數據由Excel 2016 進行整理,分析采用蓋鈞鎰等[7]的方法,并估算遺傳參數。
2 結果與分析
2.1 耐低溫性6世代次數分布
由表1、表2 可知,2 個親本具有不同的耐低溫性,在P1中表現為4 級的有37 株,表現為3 級、5 級的分別有1 株、2 株;在P2中表現為2 級的有38 株,表現為1 級的有1 株,3 級的有1 株;F1略偏向于P1,表現為3 級 的有38 株;BC1略偏向于P1,BC2略偏向于P2。從總體上看,BC1、BC2、F2等世代耐低溫性的表現分離呈正態分布。

表1 6 世代耐低溫指數次數分布Table 1 Frequency distribution in 6 family generations of chilling tolerance index

表2 耐低溫指數方差分析Table 2 Variance analysis of chilling tolerance index
2.2 遺傳模型適應性檢測
分別算出MLV 值(極大似然函數值)和AIC 值(赤池信息量準則值)見表3,按照蓋鈞鎰等[7]的理論,應選擇AIC 值較小的作為備選模型,再進行詳細比選。筆者在研究中發現,AIC 值較小的模型有E-3 模 型、B-2 模型、B-4 模型,AIC 值分別為359.861、369.544、388.036,因此確定為備選模型。

表3 各遺傳模型的MLV 值和AIC 值Table 3 The MLV values and AIC values in various genetic models
由表4 可知,E-3 模型的統計量中有7 個數據差異達到顯著或極顯著水平,分別為5.723、8.178、21.557、58.954、14.729、40.979、21.811。B-2 模型中有80.351、54.399 等9 個數據差異達到顯著或極顯著水平。B-4 模型中達到顯著或極顯著水平的統計量為11 個。經比較發現,E-3 模型中達到差異顯著水平的統計量較少,因此確定研究性狀的遺傳符合E-3 模型。
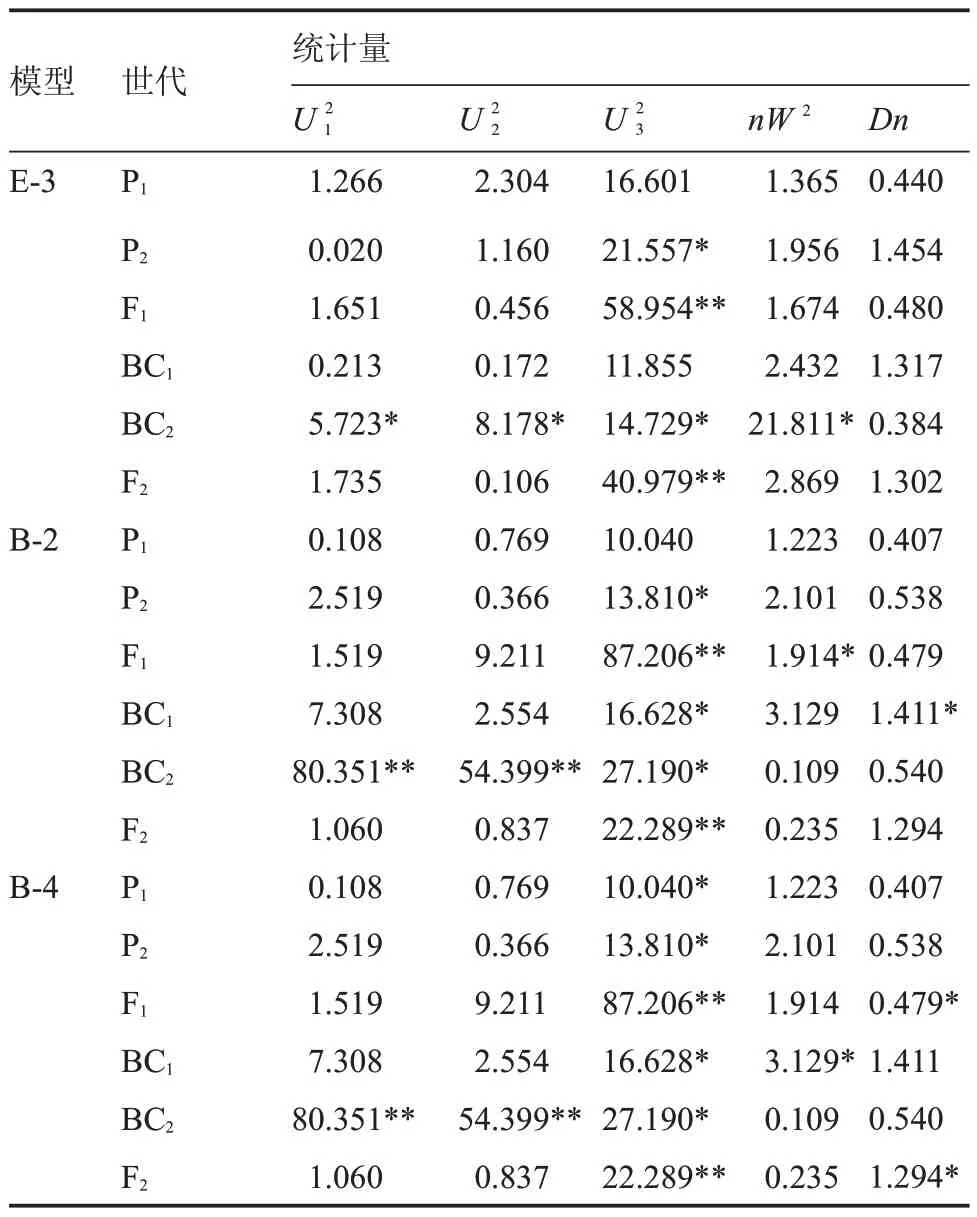
表4 耐低溫性備選模型適合性檢驗Table 4 Test for goodness of fit to standby model of chilling tolerance
2.3 遺傳參數估計
在E-3 模型下,該模型的極大似然估計值見表5,采用μ1、μ2等參數可以進一步估算一階參數、二階參數。

表5 耐低溫性遺傳模型參數的極大似然估計值Table 5 Maximum likelihood estimates of genetic models of chilling tolerance
估算一階參數、二階參數見表6。由表6 可知,中親值為2.664,2 對基因的加性效應分別為0.893、-1.103,說明親本的耐低溫性對后代的影響較大。在育種實踐中,要選擇耐低溫性均較強的雙親,以提高后代的耐低溫性。多基因加性效應為1.176,由于主基因起加性作用,因此,2 對基因的顯性效應均為0。但多基因顯性效應為0.130,有一定的作用。

表6 耐低溫性的遺傳參數估計值Table 6 The estimate of genetic parameter of chilling tolerance
分離世代中主基因遺傳率以F2為最高,達68.463%,其次為BC1、BC2;多基因遺傳率以BC2為最高。環境方差占表型方差的比率分別達36.036%、34.957%、28.543%。說明環境對后代的選擇影響較小。
綜上所述,番茄幼苗耐低溫性的遺傳受2 對加性主基因+加性-顯性多基因控制,主基因遺傳率在BC1、BC2、F2世 代 分 別 達57.357% 、54.441% 、68.463% ,多基因的遺傳率分別達6.607% 、10.602%、2.994%,基因型占表型方差的比率為63.964%、65.043%、71.457%,環境方差占表型方差的比率分別占36.036%、34.957%、28.543%。由于基因型占比較高,針對耐低溫性的選育可在早代進行。
3 討論與結論
對于蔬菜耐低溫性的遺傳研究已有報道。閆世江等[15]研究黃瓜苗期耐低溫性及相關形態學性狀的遺傳,發現黃瓜耐低溫性分級的遺傳符合加性-顯性模型。耐低溫性廣義遺傳力高,達92.133%,適合于早代選擇。該研究采用Hayman的方法,使用的是親本與F1的數據,用時較短,受限于方法本身的限制,僅能估算基因的整體作用,未能估算各個基因的作用。閆世江等[16]進一步采用蓋鈞鎰等[7]的方法研究黃瓜耐低溫性的遺傳,結果表明,耐低溫分級的遺傳符合E-3 模型,估算了各個基因的作用,2 對基因的加性效應分別為1.034、-1.277,總的效應為1.361,總的顯性效應為正向0.151。F2主基因+多基因遺傳率為82.759%,3個分離世代的主基因遺傳率均比多基因遺傳率高。環境方差占表型方差的比例在17.241%~25.974%之間。有關番茄耐低溫性分級的遺傳較少見報道,前人研究[2-6]番茄耐低溫性遺傳時采用的研究對象多為番茄低溫下的發芽能力、花粉萌發能力、坐果能力、果實發育速度等,上述性狀與番茄耐低溫性密切相關,但均為間接性狀,筆者采用的耐低溫性分級標準與上述間接性狀相比更加直觀。
筆者運用主基因-多基因聯合遺傳分析方法,結論認為番茄耐低溫性的遺傳符合E-3 模型,即2對加性主基因+加性-顯性多基因模型。結論與閆世江等[16]的相似,但估算出的遺傳參數不同,其原因可能是蔬菜種類及品種、親本遺傳來源、低溫處理的方法、分級標準不同。今后為明確番茄耐低溫性的遺傳機制應選用不同的材料進行深入的研究,并規范處理方法及耐低溫性評價標準,降低其影響程度。
筆者研究了耐低溫性,借鑒查丁石等[17]的方法制定分級標準,并進行一定的改進,該方法操作簡單,結果直觀,以后可以在此基礎上進行QTL 定位研究,為培育設施專用品種奠定基礎。番茄耐低溫性的遺傳受2 對加性主基因+加性-顯性多基因控制。今后為培育耐低溫性較強的品種,親本應選擇耐低溫性較強的材料,并在早代選擇。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
光學精密工程(2016年6期)2016-11-07 09:07:19
