鄱陽湖刀鱭繁殖生物學特征
2024-01-01 00:00:00孔赤平許群黃平徐強張寶高小平李樂康
江西水產科技 2024年4期

摘要:為掌握鄱陽湖刀鱭(Coilia nasus)群體繁殖生物學特征,于2020年和2021年4—7月對鄱陽湖刀鱭雌雄比、性腺發育情況、性腺指數和繁殖力等參數進行了研究。刀鱭性別抽樣結果顯示,繁殖初期刀鱭雌雄比接近于1.0∶1.0,繁殖期間雌雄比呈現先升后降的趨勢。通過對性腺發育和性腺指數分析發現,刀鱭在鄱陽湖的繁殖期為5月上半月至7月上半月,其中6月下半月為繁殖高峰期。鄱陽湖刀鱭絕對繁殖力均值為(37 689±26 694粒),相對繁殖力為(385±216)粒/g,絕對繁殖力與體長和體重呈正線性相關。
關鍵詞:刀鱭;鄱陽湖;性腺;雌雄比;繁殖力
中圖分類號:S931 文獻標識碼:A
基金項目:九江市科技局重點研發計劃“鄱陽湖刀鱭種群生物學研究”(S2022ZDYFN311);農業農村部淡水生物多樣性保護重點實驗室開放性課題(LFBC1009);江西省水生生物資源調查監測體系(JXSS[2023]2)
作者簡介:孔赤平(1993—),男,水產師,研究方向:漁業資源。E-mail:kongchipin@126.com
*通訊作者:李樂康(1988—),工程師,研究方向:漁業資源。E-mail:lekangli1987@126.com
刀鱭(Coilia nasus)屬鯡形目(Cluperiformes)、鳀科(Engraulidae)、鱭屬(Coilia)魚類,是我國長江流域重要經濟魚類之一[1-2]。歷史上,刀鱭資源曾十分豐富,20世紀70年代最高捕撈量曾達3545 t/a[3]。近30年來,在多種人類活動影響下,刀鱭洄游通道受阻、資源持續衰退[3-4],2011—2012年,年捕撈量已不足10 t/a[5]。為了保護刀鱭資源,農業農村部自2019年開始停止發放刀鱭專項捕撈許可證[6],禁止刀鱭天然資源的生產性捕撈以后,其下滑趨勢才得到一定程度遏制[7]。
鄱陽湖是我國面積最大的淡水湖,也是刀鱭重要的產卵場,在刀鱭繁殖、育肥和棲息等方面發揮著重要作用[8-10]。目前關于刀鱭繁殖生物學研究主要集中在長江下游江段[11-14],對鄱陽湖刀鱭繁殖生物學研究相對較少。長江流域全面禁捕后,鄱陽湖刀鱭資源呈現出的迅速恢復態勢[7],更突顯出對禁捕后鄱陽湖刀鱭種群繁殖生物學特征調查的必要性。本研究基于2020和2021年刀鱭繁殖期所采集的刀鱭樣本雌雄比、性腺成熟度、性腺指數和繁殖力等參數的調查結果分析,探討刀鱭的繁殖時間,揭示刀鱭群體繁殖生物學特征,為刀鱭的資源管理和有效保護提供基礎資料。
1 材料與方法
1.1 樣本采集
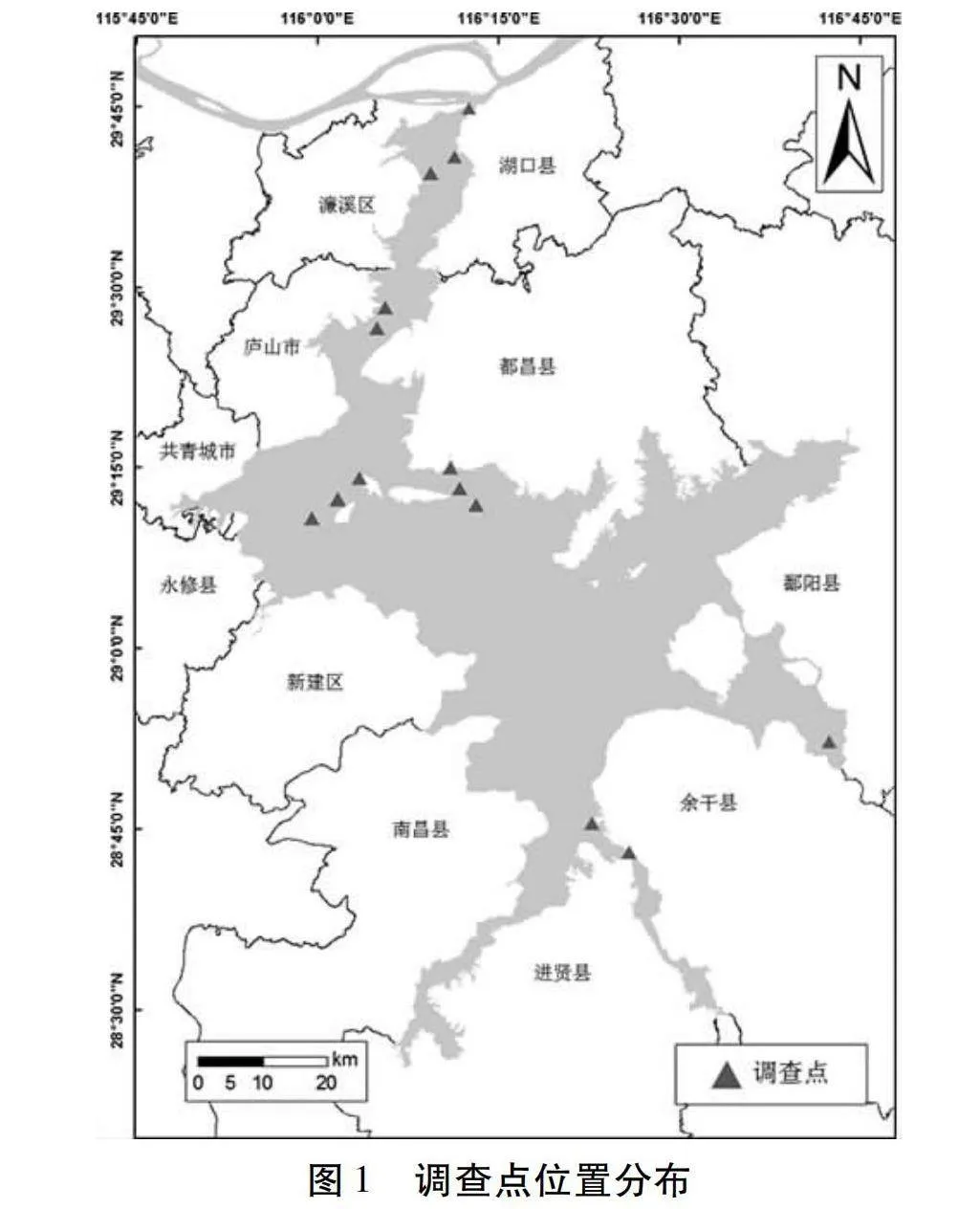
經江西省農業農村廳漁業漁政局許可辦理特許捕撈證后,于2020年和2021年4—7月,在鄱陽湖湖口縣、濂溪區、廬山市、都昌縣、永修縣、余干縣和鄱陽縣等水域設調查點(見圖1),捕撈網具為單層刺網(網長50 m,高2 m,網目2a=4 cm),按照橫向距離布設4條刺網,每次捕撈時間約12 h。野外采集樣本后帶回實驗室進行生物學測量,將目視上頜骨長大于頭長的個體初步判定為刀鱭[15]。部分樣本進行生物學解剖以鑒定性別,所獲刀鱭樣本冷凍保存,均用于調查研究。
1.2 形態學測量及性腺發育分期判定
樣本生物學測量內容包括標準體長(SL)、體質量(BW)、性別、性腺成熟度、性腺重等。體長用量魚板測定,精確至1 mm;體質量用電子秤稱量,精確至0.1 g;性腺重用電子天平稱量,精確至0.0001 g;刀鱭性腺成熟度劃分標準參考陳文銀等對刀鱭性腺發育分期特征的描述,將雌雄刀鱭性腺成熟度都分為六個時期[16]。本研究采集到性腺發育II~VI期個體,將性腺發育達到II期的刀鱭群體視為繁殖群體,性腺發育達到II~III期為性未成熟個體,性腺發育達到IV~V期為性成熟個體,性腺發育達到VI為產卵后個體[12]。
1.3 數據分析
1.3.1 性別比例
分別計算總樣本、性成熟樣本和性未成熟個體的雌雄比例。
1.3.2 性腺成熟度
分別計算總體及各半月雌雄個體性腺成熟度組成,并分別計算未成熟個體、成熟個體和產卵后個體在每個時期所占的比例。
1.3.3 性腺指數
性腺指數(Gonad somatic index,GSI)計算公式如下
GSI=GW/BW×100
式中,GSI為性腺指數,GW為性腺重(Gonad weight),單位g;BW為體重(Body weight),單位g。
1.3.4 繁殖力
刀鱭繁殖力的測算參考劉必林等建立的方法[17],分別在刀鱭IV~V期卵巢的前中后各稱取卵重0.05~0.20 g,記取卵粒數量,刀鱭的絕對繁殖力(Absolute fecundity,AF)和相對繁殖力(Relative fertility,RF)由以下公式計算:
絕對繁殖力(AF)=(卵巢重/卵巢樣品重)×樣本卵粒數
相對繁殖力(RF)=絕對繁殖力/體重
使用直線式AF=a+bSL和AF=a+bBW分別擬合刀鱭的F(卵粒數)與SL(體長)、BW(體質量)的關系,式中a和b為常數。
所有數據均在Microsoft Excel 2016和SPSS 22.0進行處理和分析,部分圖表繪制由ARC GIS 10.2和R 4.2.1軟件完成。
2 結果與分析
2.1 性別比例
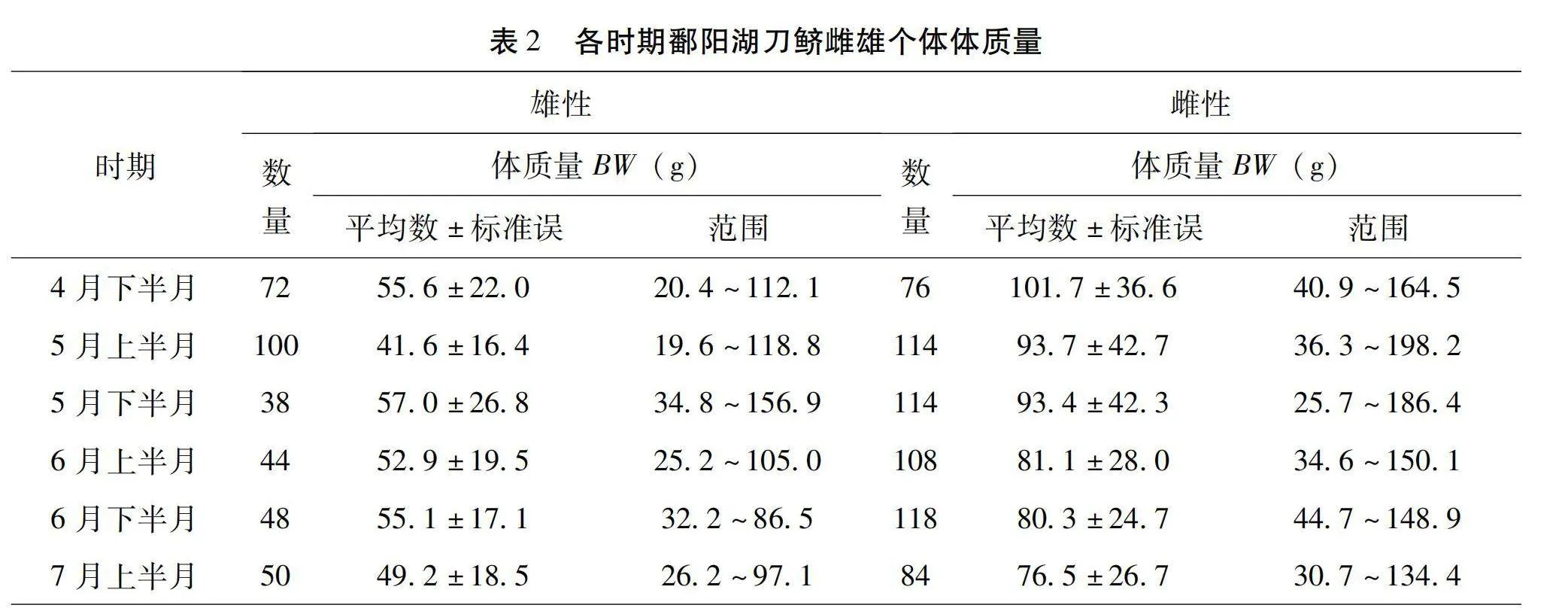
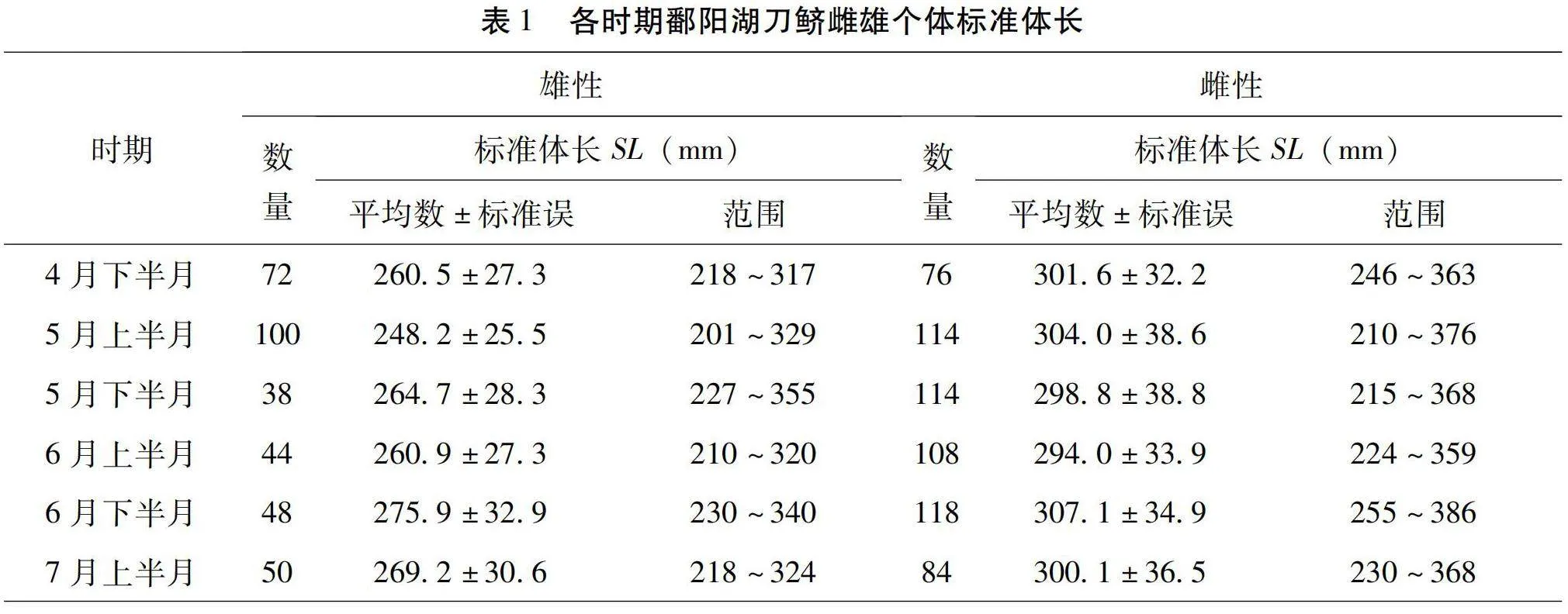
本研究共解剖觀察了966尾刀鱭,其中雄性個體為352尾,各時期雌雄個體標準體長情況見表1,各時期雌雄個體體質量情況見表2。
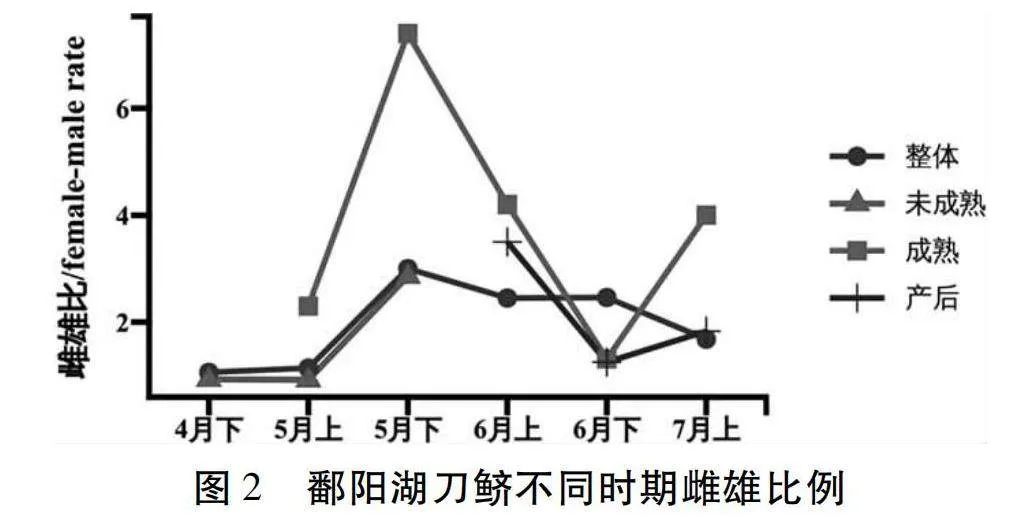
圖2所示,按月份分析發現,整體上4月下半月、5月上半月、5月下半月、6月上半月、6月下半月和7月上半月各月刀鱭雌雄比分別為1.1∶1.0、1.1∶1.0、3.0∶1.0、2.5∶1.0、2.5∶1.0、1.7∶1.0。未成熟個體雌雄比例4月下半月、5月上半月、5月下半月分別是1.0∶1.0、1.0∶1.0、2.9∶1.0。成熟個體雌雄比例5月上半月、5月下半月、6月上半月、6月下半月和7月上半月分別是2.3∶1.0、7.4∶1.0、4.2∶1.0、1.3∶1.0、4.0∶1.0。產卵后個體雌雄比例4月下半月、5月上半月、5月下半月分別是3.5∶1.0、1.3∶1、1.8∶1.0。
2.2 性腺成熟度
圖3所示,按不同月份分析:雌性樣本中,4月下半月73.7%為性腺未成熟個體,性腺成熟個體占26.3%(n=38);5月上半月59.6%為性腺未成熟個體,性腺成熟個體占40.4%(n=57);5月下半月性腺成熟個體數量開始超過成熟個體,其中35.1%為性腺未成熟個體,性腺成熟個體占64.9%(n=57);6月上半月開始出現產卵后個體,占13.0%,性腺未成熟個體占9.3%,性腺成熟個體占77.8%(n=54);6月下半月產卵后個體逐漸增多,產卵后個體占46.9%,性腺成熟個體占53.1%(n=32);7月上半月產后個體占大部分,為65.9%,性腺成熟個體占34.1%(n=41)。
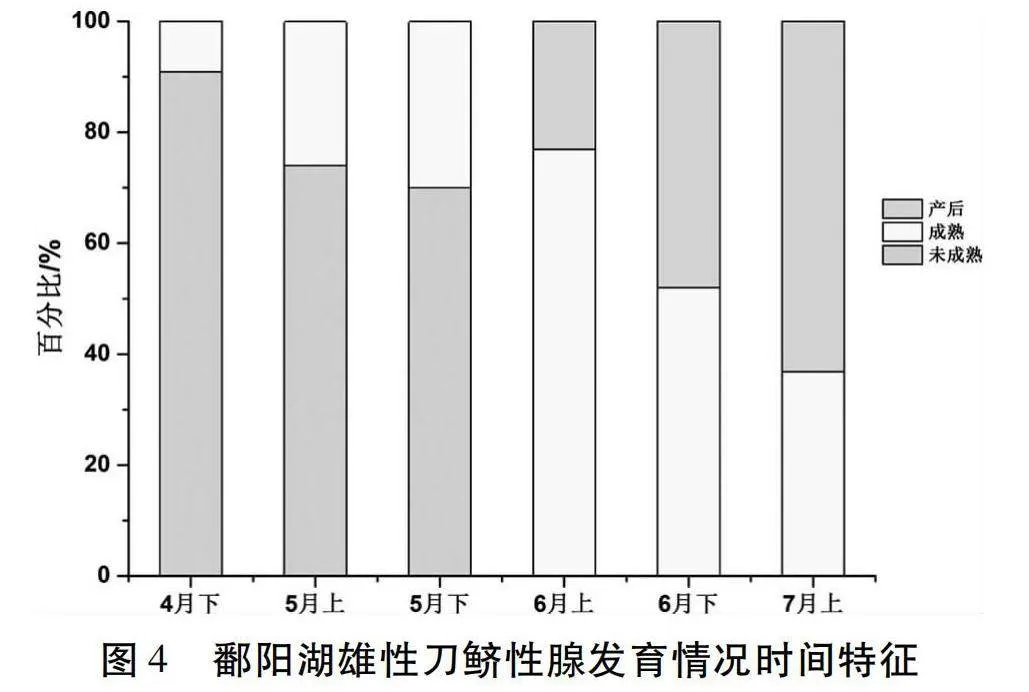
圖4所示,按不同月份分析:雄性樣本中,4月下半月90.9%為性腺未成熟個體,性腺成熟個體僅占9.1%(n=33);5月上半月74.0%為性腺未成熟個體,性腺成熟個體占26.0%(n=50);5月下半月70.0%為性腺未成熟個體,性腺成熟個體占30.0%(n=20);6月上半月開始出現產卵后個體,占23.1%,性腺成熟個體占76.9%(n=13);6月下半月產卵后個體逐漸增多,性腺成熟個體占52.0%,產卵后個體占48.0%(n=25);7月上半月產后個體占大部分,為63.2%,性腺成熟個體占36.8%(n=19)。
2.3 性腺指數
對采集到的部分刀鱭樣本統計其GSI,雌性個體共統計173尾,雄性個體共統計121尾。
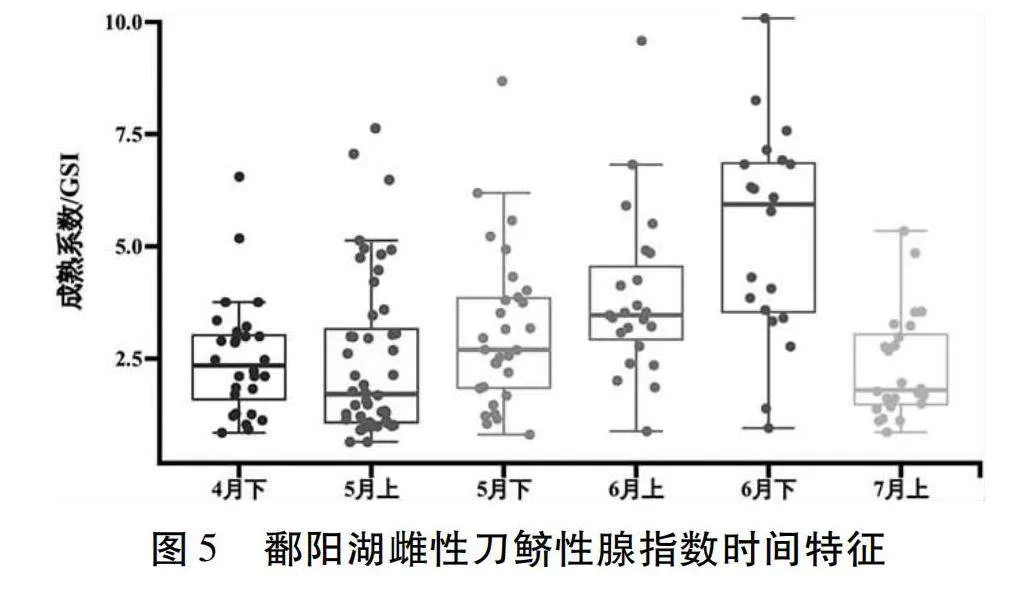
按不同性腺成熟度等級分析雌性GSI顯示(圖5),4月下半月至6月下半月刀鱭GSI隨時間的推移不斷增大,7月上半月刀鱭GSI相對于6月下半月有所下降,推測可能是排卵作用所導致。4月下半月平均GSI為2.5±1.3(n=28);5月上半月平均GSI為2.5±1.8(n=48);5月下半月平均GSI為3.1±1.8(n=30);6月上半月平均GSI為3.9±1.8(n=23);6月下半月平均GSI為5.3±2.4(n=20);7月上半月平均GSI為2.3±1.2(n=24)。
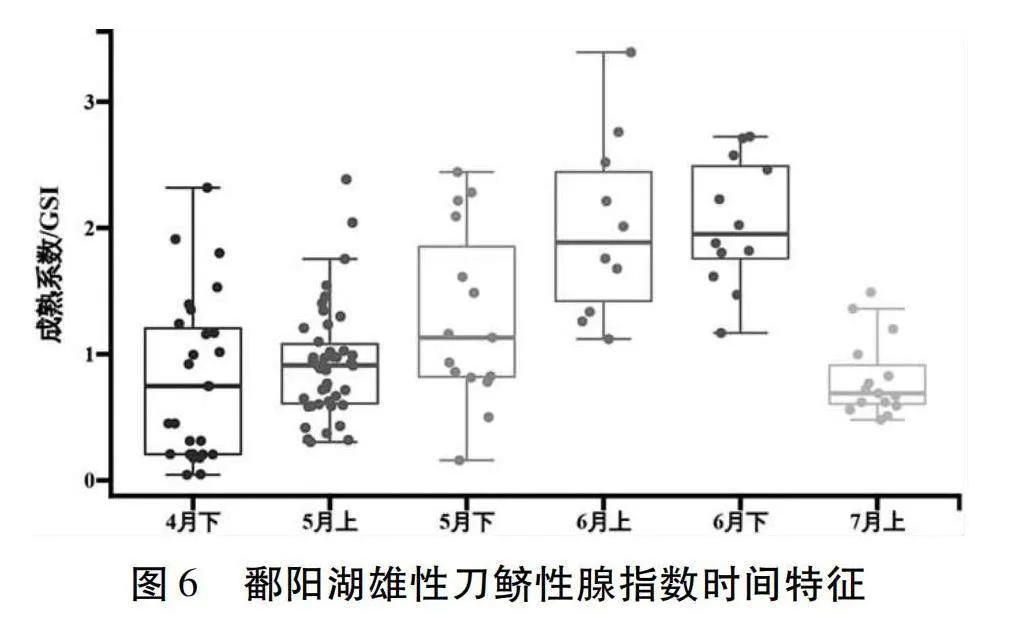
按不同性腺成熟度等級分析雄性GSI顯示(圖6),4月下半月至6月下半月刀鱭GSI隨時間的推移不斷增大,7月上半月刀鱭GSI相對于6月下半月有所下降,推測可能是精子釋放所導致。4月下半月平均GSI為0.8±0.6(n=27);5月上半月平均GSI為0.9±0.5(n=42);5月下半月平均GSI為1.3±0.7(n=15);6月上半月平均GSI為2.0±0.7(n=10);6月下半月平均GSI為2.0±0.5(n=12);7月上半月平均GSI為0.8±0.3(n=15)。
2.4 繁殖力
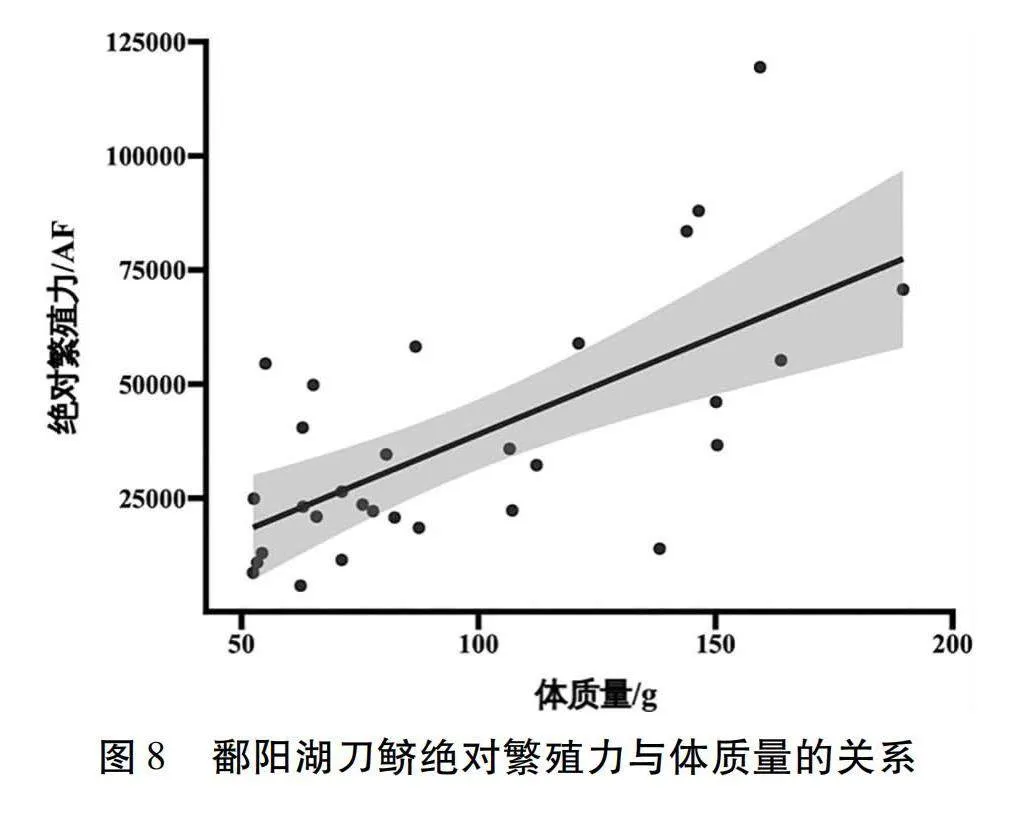
鄱陽湖刀鱭絕對繁殖力為5 841~119 411粒,均值為(37 689±26 694)粒,相對繁殖力為93~990粒/g,均值為(385±216)粒/g。
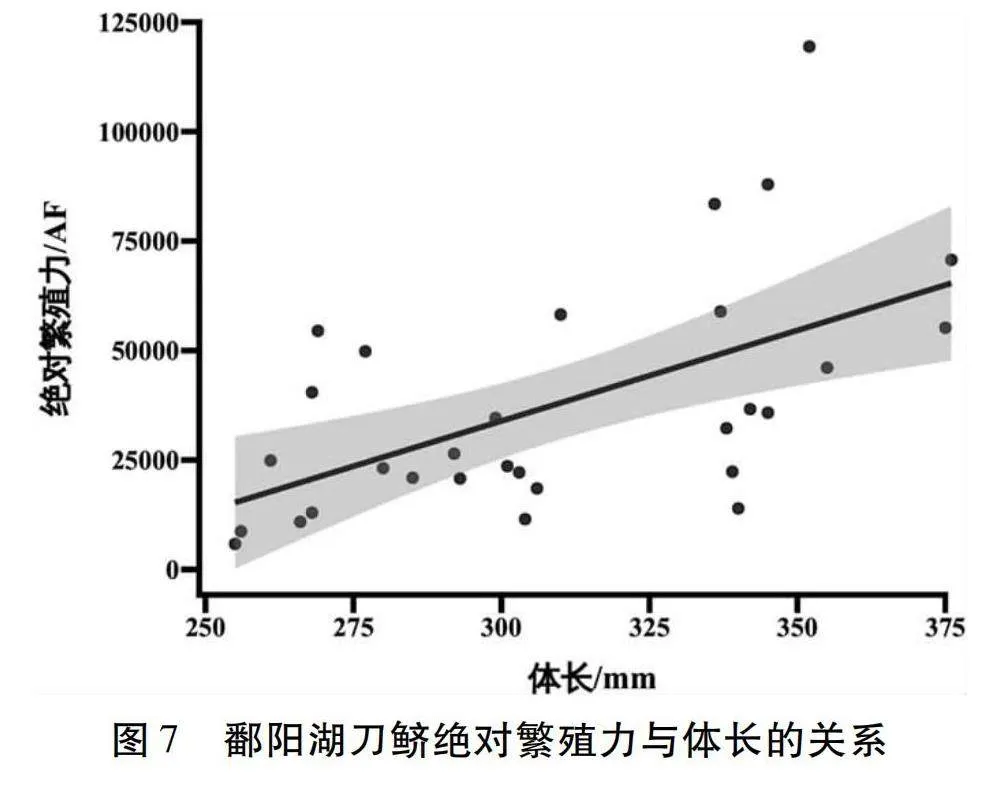
個體絕對繁殖力和體長的函數關系式為:AF=413.85×SL-90231.17(R2=0.32,n=30),如圖7所示。
個體絕對繁殖力和體質量的函數關系式為:AF=428.97×BW-3903.32(R2=0.43,n=30),如圖8所示。
3 討論
3.1 雌雄比
有研究顯示,刀鱭在進入鄱陽湖后,在雌雄比上表現為產卵前期雄性多于雌性,產卵期雄性與雌性是趨向于平衡的[18]。代培等[19]對長江安慶江段刀鱭的研究表明,溯河洄游的刀鱭群體在進入產卵場后,就時間特征而言,雌雄比表現為上升趨勢,在5月上半月最低,7月上半月最高。本研究結果與其基本一致,繁殖初期(4月下半月),進入鄱陽湖的刀鱭雌雄比接近于1.0∶1.0(1.1∶1.0),繁殖盛期(5月下半月—6月下半月),雌性數量明顯多于雄性(雌雄比最高為3.0∶1.0),繁殖后期(7月上半月),雌魚數量降低明顯,雄魚占比相對提高,雌雄比下降到1.7∶1.0。
刀鱭繁殖群體在鄱陽湖的雌雄比要高于長江下游江段(長江安慶段),推測是受到刀鱭雌雄肥滿度差異的影響。刀鱭是長距離洄游型魚類,且在上溯過程中不攝食[20],洄游過程主要依靠體內的脂肪含量[21],如果體內儲藏的脂肪不夠,很可能導致刀鱭到達不了最終的產卵場,雌性刀鱭肥滿度明顯要大于雄性[22],其體內蘊含的脂肪也更多,更利于雌性刀鱭的遠距離洄游。
3.2 性腺發育
20世紀70年代研究結果顯示,刀鱭在鄱陽湖的繁殖時間仍然是5—7月份,6月份達到峰值[18]。本研究調查結果顯示,刀鱭于每年4月中旬開始洄游至鄱陽湖,此時大多數為性腺未發育成熟個體,隨著時間的推移,性腺成熟個體不斷增多,6月上半月開始出現產卵后個體,6月下半月為其產卵高峰期,7月上半月大部分產卵完畢。
20世紀70年代,鄱陽湖刀鱭IV期雌性個體的GSI是5.2~13.0,IV期雄性個體GSI是2.0~4.5;本研究中,IV期雌性個體平均GSI僅有3.5±1.7;IV期雄性個體平均GSI僅有1.2±0.7。與歷史上相比,雌雄個體的GSI均出現了大幅度下降,表明洄游至鄱陽湖的刀鱭繁殖群體已出現了嚴重的資源衰退。為了恢復這一重要經濟漁業資源,對于刀鱭在鄱陽湖的重要棲息場所,尤其是其產卵場,應重點加強保護。
3.3 繁殖力
繁殖力是魚類繁衍生息的基礎,是評估種群動態重要指標之一[21]。本研究表明,鄱陽湖刀鱭絕對繁殖力與體長和體重呈線性相關,但體重較體長相關性優勢明顯。鄱陽湖刀鱭絕對繁殖力均值為(37 689±26 694)粒,相對繁殖力為(385±216)粒/g,但與70年代均值(絕對繁殖力106 050粒)相比還有較大差距,比大洋河刀鱭(絕對繁殖力44 617粒,相對繁殖力364粒/g)[23]和雙臺子河刀鱭(絕對繁殖力44 995粒,相對繁殖力362 粒/g)稍低[24],高于長江刀鱭(絕對繁殖力23 695 粒、229粒/g)[3]。
刀鱭作為一種小型經濟魚類,生命周期短,性成熟早,其生活史中面臨較大強度的捕撈作業和環境因素變化[25]。因此,刀鱭的繁殖策略是通過提高繁殖力的方式來保證種群的延續[26]。但在長江流域水質污染日益嚴重和長期高強度捕撈等因素影響下,刀鱭資源相比于20世紀70年代降幅達95.98%,漁汛幾近消失[14],且個體繁殖力與70年代相比也出現大幅度下降。長江流域全面禁捕以后,刀鱭面臨的捕撈壓力基本消失,刀鱭資源量恢復情況如何?其繁殖策略是否會發生變化?需要進行長期的調查研究。
3.4 刀鱭資源保護建議
歷史資料表明,鄱陽湖是刀鱭重要的產卵場[4-5],刀鱭在鄱陽湖的產卵場主要分布在東湖、草灣湖、程家池湖等20余個子湖約30萬畝的南部水域[2]。但是隨著人類活動的影響,這些原本是產卵場的水域,其環境因子已經產生了巨變,可能已不再具備刀鱭的產卵條件。因此,對刀鱭在鄱陽湖產卵場的定位,產卵場范圍的識別,仍需要進行更深入的研究。
鄱陽湖作為刀鱭重要棲息地,其生態功能的發揮對整個長江流域刀鱭資源都存在巨大的補充作用[24]。長江流域全面禁捕以后,刀鱭在鄱陽湖面臨的捕撈壓力基本消失,但諸如采砂等人類涉水活動仍對刀鱭產卵和洄游具有較大影響。對刀鱭在鄱陽湖的重要棲息地開展針對性的保護措施,確保其繁殖群體和關鍵棲息地的保護效果不受其他因素的影響、干擾和破壞。同時加強對刀鱭的深入研究,查明其在鄱陽湖的種群規模、種群結構和種群變動趨勢,以跟蹤評估刀鱭資源恢復效果,來達到恢復刀鱭資源,提升長江水生生物完整性,保護長江生物多樣性的目標。
參考文獻
[1]張世義.中國動物志:硬骨魚綱鱘形目海鰱目鯡形目鼠鱔目[M].北京:科學出版社,2001.
[2]袁傳宓,林金榜,秦安舲,等.關于我國鱭屬魚類分類的歷史和現狀——兼談改造舊魚類分類學的幾點體會[J].南京大學學報(自然科學版),1976,2:1-12.
[3]張敏瑩,徐東坡,劉凱,等.長江下游刀鱭生物學及最大持續產量研究[J].長江流域資源與環境,2005,14(6):22-26.
[4]Li Y,Xie S,Li Z,et al.Gonad development of an anadromous fish Coilia ectenes(Engraulidae)in lower reach of Yangtze River,China[J].Fisheries Science,2010,73(6):1224-1230.
[5]劉凱,段金榮,徐東坡,等.長江口刀鱭漁汛特征及捕撈量現狀[J].生態學雜志,2012,31(12):3138-3143.
[6]農業農村部.農業農村部關于調整長江流域專項捕撈管理制度的通告(農業農村部通告﹝2018﹞5號)[EB/OL].(2018-12-28)[2024-04-02].http://www.cjyzbgs.moa.gov.cn/tzgg/201904/t20190428_6220295.htm.
[7]吳金明,李樂康,程佩琳,等.鄱陽湖刀鱭的鑒定與資源動態研究[J].中國水產科學,2021,28(6):743-750.
[8]張堂林,李鐘杰.鄱陽湖魚類資源及漁業利用[J].湖泊科學,2007,19(4):434-444.
[9]吳斌,方春林,張燕萍,等.鄱陽湖刀鱭繁殖群體生物學參數及生長特性的初步分析[J].水產科學,2016,35(2):142-146.
[10]王生,方春林,周輝明,等.鄱陽湖湖口刀鱭幼魚出現的時間及漁獲量[J].江蘇農業科學,2017,45(10):122-124.
[11]何為,李家樂.長江刀鱭性腺發育規律的研究[J].中國水產,2006(5):70-72.
[12]黎雨軒.長江洄游性刀鱭的繁殖生態學研究[D].武漢:中國科學院水生生物研究所,2009.
[13]管衛兵,陳輝輝,丁華騰,等.長江口刀鱭洄游群體生殖特征和條件狀況研究[J].海洋漁業,2010,32(1):73-81.
[14]賀剛,方春林,吳斌,等.鄱陽湖刀鱭生殖群體特征及狀況分析[J].水生態學雜志,2017,38(3):83-88.
[15]程萬秀,唐文喬.長江刀鱭不同生態型間的某些形態差異[J].動物學雜志,2011,46(5):33-40.
[16]陳文銀,李家樂,練青平.長江刀鱭性腺發育的組織學研究[J].水產學報,2006,30(6):773-777.
[17]劉必林,陳新軍,李建華,等.漁業資源生物學實驗[M].北京:科學出版社,2017:66.
[18]長江流域刀鱭資源調查協作組.長江流域刀鱭資源調查報告[R].1977:54-61.
[19]代培,嚴燕,朱孝彥,等.長江刀鱭國家級水產種質資源保護區(安慶段)刀鱭資源現狀[J].中國水產科學,2020,27(11):1267-1276.
[20]姜濤,劉洪波,李孟孟,等.溯河洄游長江刀鱭(Coilia nasus)攝食蝦類的調查[J].湖泊科學,2018,30(2):458-463.
[21]董文霞,唐文喬,王磊.長江刀鱭繁殖群體的生長特性[J].上海海洋大學學報,2014,23(5):669-674.
[22]王丹婷,楊健,姜濤,等.不同水域刀鱭形態的分析比較[J].水產學報,2012,36(1):78-90.
[23]張健,楊培民,胡宗云,等.大洋河刀鱭繁殖生物學特性[J].淡水漁業,2021,51(6):91-96.
[24]霍堂斌,鄭文軍,李耕,等.雙臺子河口刀鱭個體生殖力[J].水產學雜志,2016,29(6):31-36.
[25]畢雪娟,張濤,馮廣朋,等.長江口鳳鱭個體生殖力的研究[J].海洋漁業,2015,37(3):223-232.
[26]Jiang T,Yang J,Lu M J,et al.Discovery of a spawning area for anadromous Coilia nasus Temminck et Schlegel,1846 in Poyang Lake,China[J].Journal of Applied Ichthyology,2017,33(2):189-192.
