三港雨蛙的兩性異形
2023-12-07 06:33:48蘇海龍湯艷青林心昀尤錫靜
麗水學院學報 2023年5期
關鍵詞:差異
蘇海龍,羅 艷,湯艷青,林心昀,尤錫靜,馬 力
(麗水學院生態學院,浙江 麗水 323000)
兩性異形(sexual dimorphism,SD)是指同一物種因性別不同而導致其體色、生理、體型大小、局部形態特征等方面產生差異的現象[1],其中體型大小和局部形態特征的兩性異形在動物界普遍存在[2-3],并受到人們的廣泛關注[4]。目前,對動物兩性異形產生的機理進行解釋的假說主要有3 種。傳統的性選擇假說認為,個體更大的雄性在同性競爭或異性配偶選擇中具有優勢[5],從而獲得更多的交配機會和較高的繁殖成功率,并影響雌性對雄性的選擇,使個體更大的雄性具有更高的適合度[6],從而產生兩性異形;而生育力選擇假說則認為,雌性向較大體型發展更有利于其增加生育力和繁殖輸出[5],是雌性具有較大體型的主要形成動力,這對雌性的體型進化作出了合理解釋[6];此外,生態位分離假說認為,雌雄兩性因對生態位需求的不同而產生不同的生態位分化,從而影響兩性體形的進化,導致兩性異形的出現[7-8]。另外,兩性間性成熟年齡、繁殖策略、食性分離及其生長發育過程中的物質分離及能量分配上的差異均有可能導致雌雄個體之間兩性異形的形成[9-16]。
現有關于兩性異形的研究涉及多種脊椎動物類群,其中兩棲動物因其物種多樣性高、數量繁多、分布范圍極廣、棲息地多樣、具有終生生長的生態學特性,被證明是研究兩性異形的理想模型[17]。已有的研究已對黑斑側褶蛙(Pelophylax nigromaculata)[9]、澤陸蛙(F. multistriata)[10]、金線側褶蛙(Pelophylax plancyi)[12]、虎紋蛙(Hoplobatra-chus rugulosus)[13]、大璞鈴蟾(Bombina maxima)[18]、棘腹蛙(Quasipaa boulengeri)[19]、北侖姬蛙(Microhyla beilunensis)[20]等多個物種進行了兩性異形研究,發現不同兩棲類的兩性異形模式存在顯著性差異。因此,對特定物種開展性別間形態差異的分析將有助于理解兩棲類兩性異形進化的產生機制[3,9,12,19],同時對于目標物種的生殖繁育以及物種多樣性保護具有重要意義[19]。
三港雨蛙(Hyla sanchiangensis),隸屬于兩棲綱(Amphibian)無尾目(Anura)雨蛙科(Hylidae)雨蛙屬(Hyla)[21-22]。在中國,該物種主要分布于東部和東南部,棲息于海拔500~1 560 m的山區稻田及其附近的雜草灌木與山林之中[22]。目前,有關三港雨蛙的相關研究較少,除了分類和分布區域調查外,僅見其染色體組及C-帶型的相關研究[21],關于三港雨蛙兩性異形的研究尚未見報道。鑒于此,本研究以麗水地區的三港雨蛙為研究對象,通過收集三港雨蛙雌雄成體的形態數據,研究其成體在頭體長、頭長、頭寬等外部形態特征上的差異,為理解和探究三港雨蛙兩性異形進化提供基礎資料,同時也為后期開展三港雨蛙的其他研究和保護工作提供參考。
1 材料與方法
2020—2022年,每年3月29日—4月16日,筆者開展麗水地區兩棲動物多樣性調查的同時,在山地稻田間共采集到46只三港雨蛙(♀♀/♂♂=12/34)個體。對其雌雄個體的各項形態指標進行測量,用數顯游標卡尺測定三港雨蛙的頭體長(snoutvent length)、頭長(head length)、頭寬(head width)、前肢長(fore-limb length)、后肢長(hind-limb length)、足長(foot length)等形態指標,精確到0.01 mm[8-10]。無尾兩棲類的頭體長是其吻端至泄殖腔孔的距離,頭長是吻端至上下頜關節后緣的距離,頭寬是頭部最寬處的距離,前肢長是肩關節至第三指末端的長度,后肢長是體后端正中部位至第四趾端的長度,足長是內跖突近端至第四指末端的長度[11]。用電子天平稱量體質量(Body mass),精確至0.01 g。
所有的數據用軟件(Statistica 8.0,Tulsa,USA)進行統計分析。在統計分析之前用Kolmogorov-Smirnov檢驗數據正態性,用Bartlett檢驗數據均質性。用單因素方差分析(one-way ANOVA)分析雌性個體和雄性個體的頭體長。用單因素協方差(one-way ANCOVA)分析兩性的頭長、頭寬、前肢長、后肢長、足長和體質量,均以性別為因子,頭體長為協變量。用主成分分析法分析三港雨蛙頭長、頭寬、前肢長、后肢長、足長和體質量對其形態的負載系數。本文中所有的描述性統計值均用平均值±標準誤表示,顯著性水平設置為P=0.05。
2 實驗結果
本研究共用46只三港雨蛙(♀♀/♂♂=12/34)。雌性個體和雄性個體的頭體長分別在36.49~40.88 mm和23.17~37.81 mm 之間波動,且兩性之間具有顯著性差異(F1,44=30.61,P<0.000 1;雌性>雄性);除頭長在兩性間無顯著性差異外(F1,43=0.24,P=0.63);三港雨蛙的頭寬(F1,43=8.81,P=0.004 8;雌性>雄性)、前肢長(F1,43=17.95,P<0.001;雌性>雄性)、后肢長(F1,43=32.02,P<0.000 1;雌性>雄性)、足長(F1,43= 38.68,P<0.000 1;雌性>雄性)和體質量(F1,43=20.36,P<0.000 1;雌性>雄性)在兩性間均具有顯著差異,且雌性個體大于雄性個體(表1、圖1)。
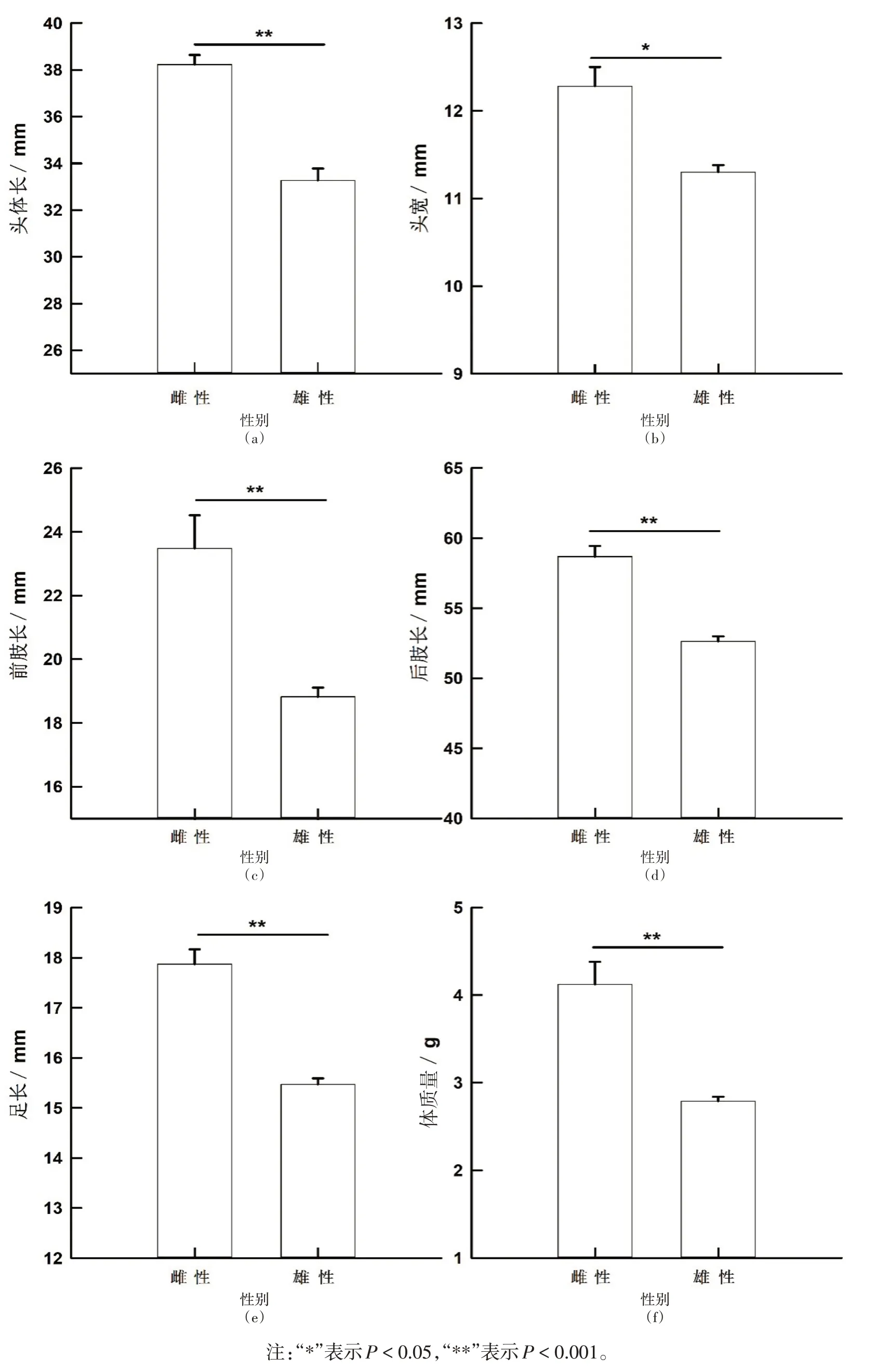
圖1 三港雨蛙成體頭體長、頭寬、前肢長、后肢長、足長和體質量的平均值±標準誤

表1 三港雨蛙的形態特征數據
對三港雨蛙6 個特定體長(SVL)的形態變量的主成分進行分析(特征值≥1),辨析出兩個主成分,共解釋了原始數據73.9%的變異(表2)。第一主成分(解釋50.8%的變異)顯示體質量、后肢長和足長具有較高的負負載系數,第二主成分(解釋23.1%的變異)顯示頭長具有較高的負負載系數,前肢長具有較高的正負載系數(表2)。

表2 6個三港雨蛙形態變量主成分分析前兩個向量的負載系數
3 討論
已有研究顯示,兩棲動物的兩性異形主要有雌性成體體型大于雄性成體、雄性成體體型大于雌性成體和雌雄個體間體型差異不大3種類型[23]。第一種是最為常見的[15],因為這對雌體提高繁殖輸出有利,在兩棲動物中,有90%的種類雌性體型大于雄性,如金線側褶蛙(Pelophylax plancyi)[12]、黑斑側褶蛙(Pelophylax nigromaculata)[9]、澤陸蛙(F.multistriata)[10]都是這一類型;第二種是雄性通常具有較大的個體或頭部,使其在種內爭斗中存在明顯優勢,有利于雄性個體獲得配偶,提高繁殖成功率[18],如飾紋姬蛙(M.fissipes)[24];第三種類型雌雄個體間體型差異不大,但某些特定的局部形態特征差異明顯,這一種類的兩性異形一般表現在第二性征上,比如齒幻蟾(Adelotus brevis)[25]雄性個體下頜出現肉質刺[14,19]。
本研究發現,三港雨蛙的頭長、頭體長、頭寬、前肢長、后肢長、足長和體質量,這7項形態特征中除頭長在兩性間沒有顯著性差異外,其余形態特征在兩性間均具有顯著差異,且雌性個體的形態特征大于雄性個體的形態特征,因此,三港雨蛙的兩性異形屬于第一種類型,這與黑斑側褶蛙[9]、澤陸蛙[10]、虎紋蛙(Hoplobatrachus rugulosus)[13]、非洲爪蟾(Xenopus laevis)[26]相似。導致三港雨蛙雌雄成體體型出現這種差異可能與性成熟年齡和繁殖投入有關。通常而言,雌性個體頭體長大于雄性說明雌性性成熟比雄性晚[27-28]。在初期的生長發育過程中,雄性將更多能量用于繁殖生長,體型較小的雄性個體相較于體型較大的雌性個體將會更早地進入繁殖期,而在較長的繁殖期中,雄性成體則將更多的能量消耗在例如鳴叫、尋覓配偶和雄性間斗爭等繁殖活動中[29],導致其用于生長和發育的能量銳減,并最終限制了其個體生長[12-13],如華南湍蛙(Amolops ricketti)[4]與康縣隆肛蛙(Feirana kangxianensis)[14]。在繁殖投入方面,雌性要大于雄性[30],從而具有更高的生育力,并使其進化趨勢朝較大體型的方向發展[14],以增加腹腔容量,占據更多的繁殖資源來提高繁殖輸出和繁殖成功率,這是生育力選擇后的結果[10,14]。
另外,頭長的兩性異形相對更為常見,具有相對較大的頭部會提高動物獲得交配權和食物的概率[14]。本研究結果顯示,三港雨蛙的頭長大小在兩性間無顯著性差異,而頭寬有顯著性差異。筆者推測這可能由以下原因造成:1)三港雨蛙的頭長對其獲得交配權和食物的概率影響較小,而頭長以外的其他形態特征則能提高其獲得交配權和食物的概率,這就促使了在兩性形態分化過程中雌性在頭長生長上能量分配較少,將更多的能量投入到其他形態的增長上[21]。2)更寬的頭寬有利于捕食更大的食物,增加了獲取食物的概率,使兩性間競爭減少,提高種群存活率[19]。3)兩棲類通過聲帶發聲來吸引異性和進行種間交流[31],而口腔是發聲系統的重要組成部分,三港雨蛙雄性具有相對短的頭長和相對小的頭寬可能與其獨特叫聲有關[32]。
綜上所述,三港雨蛙具有顯著的兩性異形,且雌性個體大于雄性個體。已有研究表明,兩性之間導致兩性異形的因素可能還有生態位、食性及其生長發育各階段的能量分配上的差異等[10,13,32]。因此,三港雨蛙兩性異形與雌雄兩性間在食物選擇、空間利用、食性及能量分配策略之間的關聯性有待于進一步研究。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
