日糧添加丁酸梭菌對小尾寒羊脂肪酸代謝和肉品質的影響
2023-11-07 11:08:44王威皓康樂天王宏迪趙麗華武海峰
食品科學 2023年19期
王威皓,竇 露,康樂天,王宏迪,段 艷,蘇 琳,趙麗華,武海峰,靳 燁,*
(1.內蒙古農業大學食品科學與工程學院,內蒙古 呼和浩特 010018;2.察哈爾右翼中旗農畜產品質量安全中心,內蒙古 烏蘭察布 013550)
羊肉具有肉嫩味鮮、低脂肪和高蛋白等特點,隨著生活水平的提升和健康意識的增強,人們對肉類食品的食用品質以及營養價值的追求越來越高。肉中的脂肪酸不僅具有較高的營養價值,還可以影響肉類食品的風味。前人研究發現脂質化合物(如磷脂酰膽堿、甘油三酯、飽和脂肪酸和不飽和脂肪酸)是肉類風味形成中重要的前體物質,多不飽和脂肪酸氧化產生醛、醇、酮及酯類等揮發性物質在適宜范圍內能提高肉質的香味[1]。在肌肉中,脂肪酸主要存在于肌內脂肪,與肉質的多汁性、適口性及嫩度密切相關,因此提高肌內脂肪沉積也成為高檔羊肉生產過程中的關鍵技術難點和突破點[2]。
反芻動物產品中的脂肪酸組成及含量主要受日糧營養、瘤胃微生物代謝以及脂肪代謝相關分子調控網絡的影響,其中瘤胃微生物可通過生物氫化作用和瘤胃發酵影響肌肉的脂肪酸組成[3]。Gillis等[4]研究發現瘤胃微生物可將68%~84%的不飽和脂肪酸氫化為飽和脂肪酸。Dai Xiaoxia等[5]研究發現瘤胃內Selenomonas_l屬的豐度與α-亞麻酸的含量呈負相關,該菌屬通過參與琥珀酸-丙酸途徑產生ATP,為氫化α-亞麻酸提供能量。因此調控瘤胃微生物結構組成可以促進肌肉中有益脂肪酸的沉積。已有研究發現益生菌在調節動物胃腸道微生態平衡、改善動物生長性能和提高肉品品質方面均具有良好的特性[6]。丁酸梭菌(Clostridium butyricum)是一種革蘭氏陽性益生菌,能夠在機體中發揮免疫、抗氧化以及抑制致病菌的作用。此外,丁酸梭菌代謝產生的短鏈脂肪酸-乙酸和丁酸是肌內脂肪合成過程中重要的前體物質[7]。Cai Qiaoli等[8]在環江小型豬日糧中添加丁酸梭菌,發現環江小型豬肌肉中非必需氨基酸含量、單不飽和脂肪酸含量和大理石紋評分顯著提高。Tian Zhimei等[9]發現羅伊氏乳桿菌能促進育肥豬肌肉中硬脂酰輔酶A去飽和酶(stearoyl-CoA desaturase,SCD)基因表達,提高肌內脂肪的含量。以上研究證明益生菌能影響機體脂肪酸代謝和肌肉品質,但通過飼喂益生菌建立反芻動物瘤胃微生物和脂肪代謝相關基因與脂肪酸代謝的關系,進而改善肉品質的研究相對較少。因此本實驗基于宏基因組測序及實時熒光定量聚合酶鏈式反應(real time quantitative polymerase chain reaction,qPCR)分析日糧添加丁酸梭菌對小尾寒羊脂肪酸代謝和肉品質的影響,為改善羊肉品質提供新的營養干預策略。
1 材料與方法
1.1 材料、菌株與試劑
3 月齡小尾寒羊共24 只(體質量(28.85±2.01)kg)選取自內蒙古自治區呼和浩特市和林格爾縣牧場。
丁酸梭菌 山東寶來利來生物工程股份有限公司。 E.Z.N.A.D N A 試劑盒 美國Omega 公司;瓊脂糖 西班牙Biowest公司;37 種脂肪酸甲酯混合標準品 美國Sigma公司;RNAiso Plus總RNA提取試劑盒、SYBR?Premix ExTaqTMII qPCR試劑盒 大連寶生物工程有限公司。
1.2 儀器與設備
TU-1810型紫外分光光度計 北京普析通用儀器有限公司;TCP2全自動測色色差計 上海生物生化實驗儀器公司;DYY-6C電泳儀 北京市六一儀器廠;8890氣相色譜儀 美國安捷倫公司;LightCycler?480 qPCR儀羅氏診斷產品(上海)有限公司;pH-STAR型胴體直測式pH計 北京賽多利斯科學儀器有限公司;C-LM3B型數顯式肌肉嫩度儀由東北農業大學研發制備。
1.3 方法
1.3.1 實驗設計
采用單因素隨機設計,將24 只小尾寒羊分為對照組和丁酸梭菌組,公母各半,每組3 個重復,每個重復4 只。對照組每日8∶00和15∶00提供基礎日糧(青貯33.4%(以體系質量計,下同)、玉米46.6%、豆粕15.7%、石粉1.1%、食鹽0.8%、磷酸氫鈣0.4%、礦物質2%),丁酸梭菌組每只羊在飼喂基礎日糧的基礎上,參考Tian Zhimei等[9]的實驗方法進行飼喂,每天添加5 g丁酸梭菌于晨飼前與精料混合。基礎日糧營養成分含量(以干質量計)為粗蛋白35.6 g/100 g、粗灰分24.8 g/100 g、粗纖維7.6 g/100 g、鈣1.13 g/100 g、磷0.67 g/100 g、氯化鈉0.9 g/100 g、賴氨酸0.6 g/100 g、增重凈能9.62 MJ/kg,除增重凈能為計算值外,其余均為實測值。丁酸梭菌由山東寶來利來生物工程股份有限公司提供,活菌數為5×108CFU/g。實驗期間自由采食和飲水,并定期清理羊舍。預試期7 d,正試期90 d。
1.3.2 樣品采集與處理
實驗期結束禁食禁水12 h,屠宰后每組取6 只羊的瘤胃內容物,用4 層紗布過濾出瘤胃液,分裝于無菌無酶的5 mL凍存管中,置于液氮中存放。屠宰后每只羊采集10~13 根肋間背最長肌2 份,每份約150 g,其中一份沿肌纖維方向進行分割處理,取3 塊1 cm2肉塊置于無菌無酶管于液氮中存放,用于脂肪酸代謝相關基因測定,剩余肉樣在宰后1 h內用于肌肉色澤、蒸煮損失率和嫩度等品質指標的測定。另一份肉樣在-20 ℃條件下冷凍運送回實驗室,用于脂肪酸分析。
1.3.3 肉品質的測定
pH值的測定:用pH-STAR型胴體直測式pH計測定肌肉45 min和4 ℃排酸24 h的pH值(pH45min和pH24h),每塊樣品重復測定3 次。
色澤的測定:使用TCP2全自動測色色差計對背最長肌色澤進行測定,每個樣品測定3 次后取平均值,L*值表示亮度,a*值表示紅度,b*值表示黃度,飽和度按照公式(1)計算。
蒸煮損失率的測定:取100 g左右的肉樣記錄煮前質量(m1/g),肉樣中心插入溫度計,放入85 ℃水浴鍋中加熱40 min,待肉樣中心溫度達到70 ℃后取出冷卻至室溫,擦干表面水分記錄煮后質量(m2/g),蒸煮損失率按照公式(2)計算。
剪切力的測定:將測定蒸煮損失剩余肉樣沿肌纖維方向切成3 cm×1 cm×1 cm的肉條,使用C-LM3B型嫩度儀測定6~8 次,以剪切力表征嫩度。
肌內脂肪質量分數的測定:根據GB 5009.6—2016《食品安全國家標準食品中脂肪的測定》[10]中索氏抽提法測定樣品中肌內脂肪質量分數,每塊樣品重復測定3 次。
1.3.4 脂肪酸組成測定
參照Floch等[11]與GB 5009.168—2016的方法制備脂肪酸甲酯,采用氣相色譜儀分析,每個樣品重復測定2 次。氣相色譜條件:使用Rt-2560石英毛細管色譜柱(100 m×250 μm,0.20 μm),程序升溫分3 步:溫度由100 ℃升高至180 ℃,速率為10 ℃/min,穩定6 min后繼續將溫度提高至200 ℃,速率為1 ℃/min,穩定20 min后繼續升溫至230 ℃,速率為4 ℃/min,穩定20.5 min后結束升溫。離子源溫度和傳輸線溫度分別為250 ℃和280 ℃。通過Sigma-Aldrich混合標準品保留時間確定各峰對應脂肪酸,根據峰面積對單個脂肪酸進行量化,并表示為總脂肪酸的相對含量。
1.3.5 脂肪酸代謝相關基因表達量的測定
參照王柏輝[12]的方法,稱取約150 mg的肉樣提取RNA。提取后的RNA樣品測定吸光度及RNA濃度,用質量分數1%瓊脂糖凝膠電泳檢測總RNA質量,然后將RNA樣品質量濃度稀釋至500 ng/μL,采用反轉錄試劑盒反轉錄獲得cDNA。采用SYBR染料法和LightCycler?480系統進行qPCR分析,每個基因重復測定2 次。相關引物的設計如表1所示,均由上海生工生物工程有限公司設計并合成,以β-actin為內參基因,基因相對表達量用2-ΔΔCt法計算。
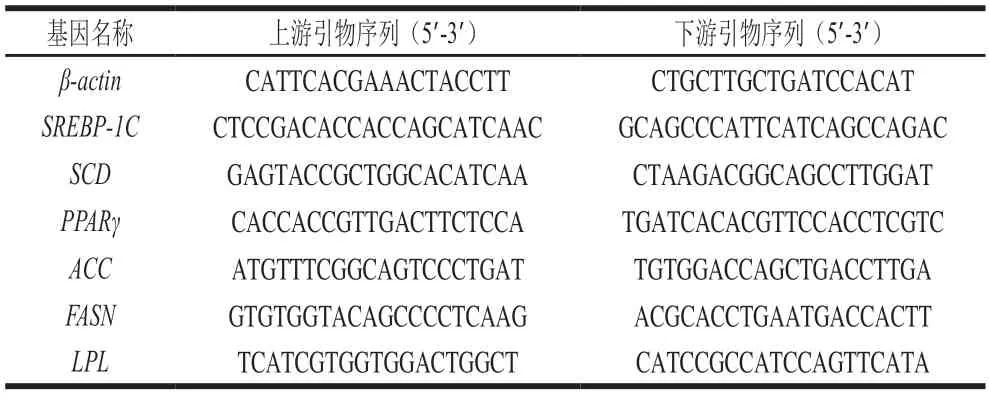
表1 qPCR引物序列Table 1 Primer sequences used for qPCR
1.3.6 瘤胃宏基因組測序分析
將采集好的瘤胃液樣品取出,按照E.Z.N.A.DNA試劑盒說明書提取樣品總DNA,選取樣品DNA經過質量分數1%瓊脂糖凝膠電泳后,選取條帶較好、無明顯降解、OD260nm/OD280nm在1.8~2.0之間的DNA送至北京諾禾致源科技股份有限公司進行Illumina PE150平臺測序。
下機后的原始數據采用Readfq(version 8)軟件(https://github.com/cjf!elds/readfq)進行預處理,得到Clean Data,參考文獻進行基因組裝[13]、開放閱讀框預測[14]、去冗余基因[15]等信息處理,獲得最終用于后續分析的Unigenes。將Unigenes與美國國家生物技術信息中心(National Center for Biotechnology Information,NCBI)的NR數據庫(version 2018-01-02)(https://www.ncbi.nlm.nih.gov/)進行比對,獲得各個分類層級上的豐度信息表。
1.4 數據處理與分析
利用SPSS 26.0 軟件進行獨立樣本t檢驗,以P<0.05表示差異顯著,結果以平均值±標準差表示。使用R v3.6.3軟件中Pearson相關系數法進行相關性分析,使用Prism 8.0軟件作圖。宏基因組原始數據已上傳https://submit.ncbi.nlm.nih.gov/subs/sra/(查詢號PRJNA882361)。
2 結果與分析
2.1 丁酸梭菌對小尾寒羊肉品質的影響
pH值是影響肉質的重要因素,畜禽屠宰后肌肉pH值下降會降低蛋白質在肌肉中的溶解度,對肉質產生不利影響。由表2可知,兩組的初始pH值(pH45min)無顯著差異(P>0.05),靜置排酸24 h后,丁酸梭菌組的pH24h顯著高于對照組(P<0.05),說明日糧添加丁酸梭菌能夠顯著減緩宰后肌肉的糖酵解進程,這與白艷蘋等[16]的研究結果一致。肉色是確保消費者滿意度的重要感官指標,飽和度越大表明肉的色澤越飽滿。先前的研究表明日糧添加益生菌可以增強肌肉氧化穩定性,減緩組織內氧合肌紅蛋白被氧化為高鐵血紅蛋白的速率,保持肉紅度穩定[17]。本研究中,對照組的L*值、a*值和飽和度均顯著低于丁酸梭菌組(P<0.05),說明丁酸梭菌對改善小尾寒羊肉色具有積極作用。Liu Yanhan等[18]也同樣證實了日糧添加丁酸梭菌能顯著提升鴨肉的紅度。剪切力能客觀反映嫩度,剪切力越小肉質越嫩。本實驗中,對照組的剪切力顯著高于丁酸梭菌組(P<0.05),這與董改香等[19]的研究結果一致,即用益生菌發酵飼糧可以顯著降低綿羊肌肉的剪切力。研究表明,肌內脂肪沉積在肌束之間,與肌肉中的膜蛋白緊密結合,其含量與肉質多汁性、嫩度及風味呈顯著正相關[20]。本實驗中丁酸梭菌組的肌內脂肪質量分數顯著高于對照組(P<0.05),說明丁酸梭菌的添加可促進脂肪在肌肉中的沉積,這也為益生菌改善羊肉嫩度提供了合理解釋。
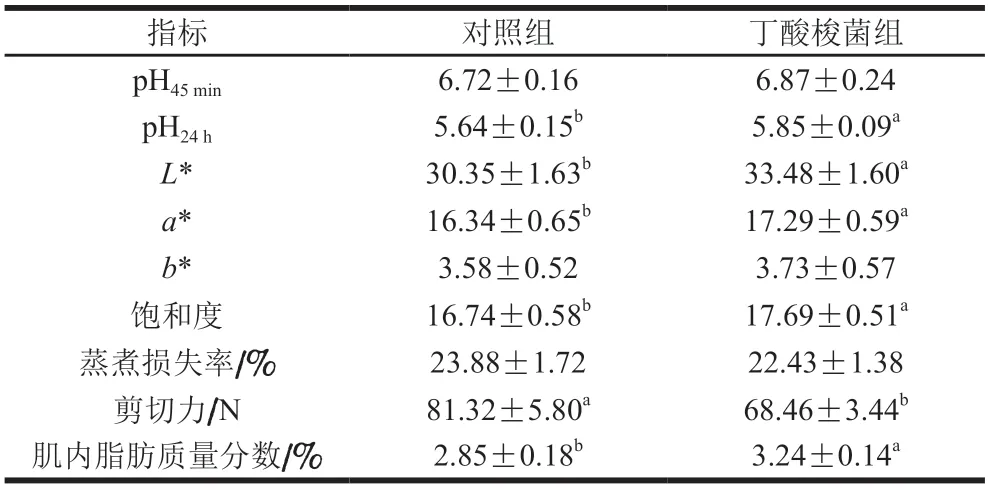
表2 丁酸梭菌對小尾寒羊肉品質的影響Table 2 Effect of Clostridium butyricum on meat quality of small-tailed Han sheep
2.2 丁酸梭菌對小尾寒羊脂肪酸組成的影響
肌肉中的脂肪酸組成可決定肌肉的氧化穩定性和硬度,進而影響其風味、嫩度和色澤[21]。反芻動物由于瘤胃中不飽和脂肪酸的高度生物氫化,導致肉類中飽和脂肪酸含量較高,而人體攝入過量的飽和脂肪酸則會增加血液膽固醇含量,從而危害身體健康[22]。丁酸梭菌對小尾寒羊飽和脂肪酸組成的影響如表3所示,對照組總飽和脂肪酸的相對含量顯著高于丁酸梭菌組(P<0.05)。飽和脂肪酸中的硬脂酸是形成羊肉膻味的主要物質,其含量越高,膻味越大[23]。本研究中,丁酸梭菌組小尾寒羊背最長肌中硬脂酸的相對含量顯著低于對照組(P<0.05),可能的原因是硬脂酸主要來源于不飽和C18脂肪酸的生物氫化,日糧添加丁酸梭菌產生的丁酸等代謝產物降低了瘤胃pH值,導致聚離子脂肪酸的生物加氫活性降低,進而減少C18:0脂肪酸的沉積[24]。與丁酸梭菌組相比,對照組棕櫚酸的相對含量顯著更高(P<0.05),而月桂酸和棕櫚酸攝入過高容易引起血清低密度脂蛋白水平升高,增加患心血管疾病的風險[25]。
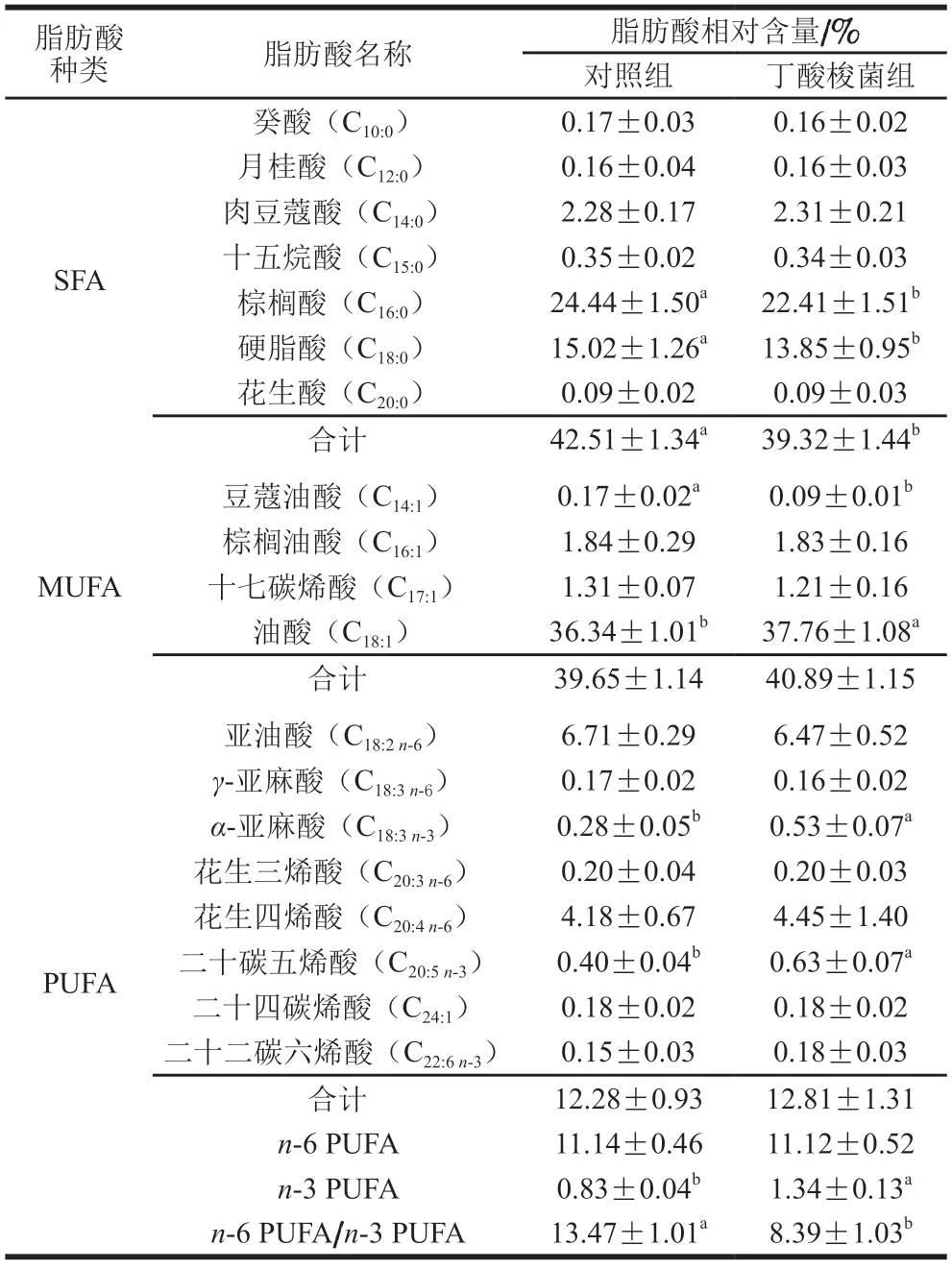
表3 丁酸梭菌對小尾寒羊背最長肌中脂肪酸的影響Table 3 Effect of Clostridium butyricum on fatty acid composition in Longissimus dorsi muscle of small-tailed Han sheep
油酸是羊肉中主要的不飽和脂肪酸,對降膽固醇和調節血脂有明顯的改善作用[26]。本研究中丁酸梭菌組小尾寒羊背最長肌中油酸的相對含量顯著高于對照組(P<0.05),這與Tang Xiaopeng等[27]的研究結果相一致。有研究表明益生菌的添加有利于增加肌肉組織中Δ9-去飽和酶活性,促進機體將硬脂酸內源性轉化成油酸[28]。肉類中功能性脂肪酸多為不飽和脂肪酸,其中亞麻酸作為中長鏈脂肪酸合成的底物,對羊肉風味的形成起重要作用[29]。本研究發現,與對照組相比,丁酸梭菌組α-亞麻酸相對含量顯著升高(P<0.05),亞油酸和γ-亞麻酸的相對含量降低,但差異不顯著(P>0.05)。二十碳五烯酸和二十二碳六烯酸屬于功能性長鏈脂肪酸,具有促進神經系統健康的作用,有利于抗衰老和維持認知功能[30]。本研究發現丁酸梭菌干預顯著提高了小尾寒羊肌肉的二十碳五烯酸相對含量(P<0.05),進一步說明益生菌能提高小尾寒羊肉的保健功效。研究表明,當n-6多不飽和脂肪酸(polyunsaturated fatty acids,PUFA)/n-3 PUFA的比值處于4~10時,更符合人們的食用健康標準[31],本實驗中丁酸梭菌組n-6 PUFA/n-3 PUFA的比值顯著低于對照組,更加符合推薦標準。
2.3 丁酸梭菌對小尾寒羊脂肪酸代謝相關基因表達量的影響
脂肪沉積是多種基因、蛋白以及多條信號通路共同作用調控的結果,相關基因表達可調控與脂肪代謝相關的物質,最終影響脂肪的沉積與降解。固醇調節元件結合蛋白1C(sterol regulatory element binding protein 1C,SREBP-1C)是細胞核內調控脂質代謝的重要轉錄因子,能夠調控膽固醇和脂肪酸的合成。SREBP-1C通過激活SCD的表達來調控下游過氧化物酶體增殖物激活受體γ(peroxisome proliferator-activated receptor γ,PPARγ)基因[32]。PPARγ進一步調控乙酰輔酶A羧化酶(acetyl-CoA carboxylase,ACC)、脂肪酸合成酶(fatty acid synthetase,FASN)和脂蛋白脂酶(lipoprotein lipase,LPL)來誘導骨骼肌中脂滴的積累,進而增加肌內脂肪的含量[33]。
由圖1 可知,丁酸梭菌組小尾寒羊背最長肌的SREBP-1C、SCD、PPARγ和ACC基因表達量顯著高于對照組(P<0.05)。Hsieh等[34]在對雄性大鼠的研究中發現羅伊氏乳桿菌能激活SREBP-1C,從而促進大鼠脂肪組織中脂肪的沉積。SCD能催化飽和脂肪酸脫氫生成單不飽和脂肪酸,如硬脂酸轉化成油酸,并且與前體脂肪細胞分化密切相關[35]。本研究中丁酸梭菌組的SCD基因表達量顯著高于對照組,并且丁酸梭菌組背最長肌中油酸的相對含量更高,可見日糧添加丁酸梭菌能夠改變小尾寒羊背最長肌SCD基因表達水平,進而影響肌肉組織中脂肪酸的組成。基因可以調控脂肪酸代謝,而脂肪酸也會對基因產生影響,如油酸會促進PPARγ的表達[36]。PPARγ是核受體超家族成員之一,能促進機體內脂肪的分化和積累,在調節肌內脂肪代謝過程中起核心作用[37]。本研究中丁酸梭菌組小尾寒羊背最長肌的PPARγ基因表達量顯著高于對照組(P<0.05),其原因可能與丁酸梭菌代謝產生的短鏈脂肪酸——丁酸有關,該脂肪酸通過調控PPARγ信號通路影響脂肪的生成[38]。Ma Jie等[39]在沙子嶺豬的飼糧中補充約氏乳桿菌,發現沙子嶺豬腹部脂肪組織中PPARγ基因表達量顯著上調,促進了腹部脂肪的積累。ACC是催化長鏈脂肪酸合成的限速酶,還可催化乙酰輔酶A縮合為丙二酸單酰輔酶A,進而調控脂肪酸的氧化進程[40]。因此,本研究對該基因進行了測定,結果顯示丁酸梭菌組ACC相對表達量(1.15)顯著高于對照組(0.81)(P<0.05),ACC的高表達能增加丙二酸單酰輔酶A活性,促進小尾寒羊肌內脂肪的沉積。張月等[41]利用乳酸菌飼喂蘇尼特羊發現,乳酸菌通過抑制腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)磷酸化,提高ACC基因的表達,進而促進脂肪酸沉積。

圖1 丁酸梭菌對小尾寒羊脂肪酸代謝相關基因表達量的影響Fig.1 Effect of Clostridium butyricum on expression levels of genes associated with fatty acid metabolism in small-tailed Han sheep
2.4 脂肪酸相關調控基因表達量與脂肪酸組成相關性分析
動物脂肪代謝是脂肪合成和脂肪分解動態博弈的結果,受脂肪酸代謝相關基因的調控,因此對小尾寒羊背最長肌中脂肪酸代謝相關基因表達量與脂肪酸組成進行Person相關性分析。由圖2可知,PPARγ基因相對表達量與α-亞麻酸和二十二碳六烯酸相對含量呈顯著或極顯著正相關(P<0.05或P<0.01),ACC和SREBP-1C基因相對表達量與α-亞麻酸和二十碳五烯酸相對含量呈顯著或極顯著正相關(P<0.05或P<0.01),推測PPARγ、SREBP-1C和ACC基因的高表達有利于不飽和脂肪酸的沉積。張秋旭[42]的研究表明n-3 PUFA能提高PPARγ基因的表達水平,活化的PPARγ通過調控LPL、ACC、SREBP-1C等靶基因,上調脂肪酸去飽和程度,從而促進肉中不飽和脂肪酸的沉積。SCD基因相對表達量與飽和脂肪酸相對含量呈極顯著負相關(P<0.01),但與油酸、α-亞麻酸和二十碳五烯酸相對含量呈顯著或極顯著正相關(P<0.05或P<0.01),Li Changxi等[43]研究發現,SCD基因的高表達有助于不飽和脂肪酸的沉積以及調控棕櫚酸和硬脂酸的去飽和化過程,這與本實驗結果一致。
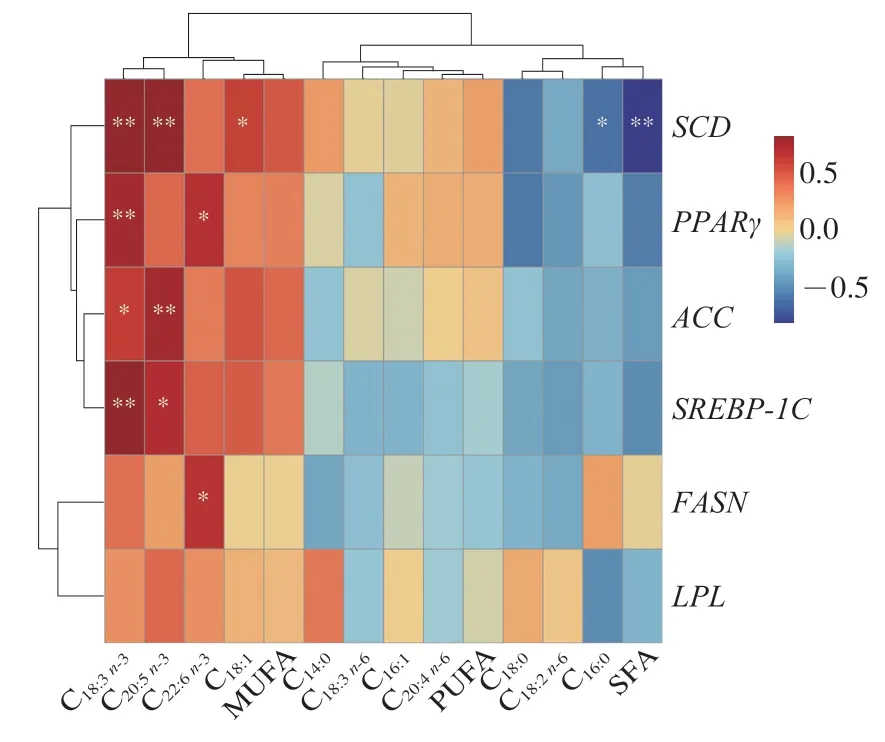
圖2 脂肪酸代謝相關基因表達量與脂肪酸組成相關性熱圖Fig.2 Heatmap showing correlation between expression of fatty acid metabolism-related genes and fatty acid compositions
2.5 丁酸梭菌對小尾寒羊瘤胃微生物組成的影響
瘤胃微生物是“第二基因組”,在調控肌肉脂肪酸組成和風味物質變化中發揮重要作用。由表4門水平上相對豐度(排名前6的菌門)可知,厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)和變形菌門(Proteobacteria)是瘤胃樣本中的優勢菌門。其中丁酸梭菌組中厚壁菌門的相對豐度顯著高于對照組(P<0.05),而擬桿菌門的相對豐度在兩組間無顯著差異(P>0.05)。劉宇陽[44]的研究也證實日糧添加復合益生菌能提高波爾山羊瘤胃中厚壁菌門的相對豐度。據報道,擬桿菌門和厚壁菌門均能調節宿主脂質代謝,提高能量效率,其中擬桿菌門能促進非纖維碳水化合物的消化,而厚壁菌門能促進纖維物質的分解[45]。本研究中,添加丁酸梭菌可以增加瘤胃中厚壁菌門的數量,對纖維物質的分解有益。變形菌門中的細菌包括多數致病菌,如大腸桿菌、沙門氏菌、幽門螺桿菌[46]。本實驗中丁酸梭菌組瘤胃中變形菌門的相對豐度顯著低于對照組(P<0.05),原因可能是丁酸梭菌能夠合成和分泌短鏈脂肪酸和細菌素,破壞致病菌的細胞膜,從而殺死或抑制致病菌生長[47]。

表4 丁酸梭菌對小尾寒羊瘤胃微生物相對豐度的影響Table 4 Effect of Clostridium butyricum on the relative abundance of rumen microflora in small-tailed Han sheep %
從表4 中屬水平相對豐度排名前10 的菌屬可知,相對豐度最高的3 個菌屬分別為普雷沃氏菌屬(Prevotella)、擬桿菌屬(Bacteroides)和梭菌屬(Clostridium)。最新的研究表明梭菌屬能夠利用纖維素、細胞壁多糖、木質纖維素等原料,提高畜禽的纖維降解能力[48]。本研究中,丁酸梭菌組梭菌屬的相對豐度顯著高于對照組(P<0.05),說明丁酸梭菌成功定植于小尾寒羊瘤胃中,一定程度上提高了小尾寒羊瘤胃的纖維降解能力。纖維桿菌屬(Fibrobacter)可代謝產生乙酸、丙酸等短鏈脂肪酸,通過三羧酸循環為宿主提供能量[49]。本研究中丁酸梭菌組纖維桿菌屬的相對豐度顯著高于對照組(P<0.05),與Miguel等[50]的研究結果一致,即在奶牛日糧中添加丁酸梭菌能顯著增加瘤胃纖維桿菌的豐度。丁酸弧菌屬(Butyrivibrio)是主要參與多不飽和脂肪酸生物氫化的瘤胃微生物,在共軛亞油酸和SFA合成過程中發揮重要作用[51]。本研究中丁酸梭菌組瘤胃中丁酸弧菌屬相對豐度顯著低于對照組(P<0.05),這可能有利于更多的多不飽和脂肪酸沉積在肉中。副擬桿菌屬(Parabacteroides)是擬桿菌門中的一種革蘭氏陰性細菌,代謝終產物主要是乙酸和琥珀酸,另外其代謝合成的主要脂肪酸為飽和直鏈脂肪酸和anteiso-甲基支鏈脂肪酸[52]。Young等[53]研究發現,甲基支鏈脂肪酸是造成羊肉中膻味的重要物質,本研究結果發現丁酸梭菌會顯著降低副擬桿菌屬的相對豐度(P<0.05),這可能對羊肉風味的形成具有積極作用。總體而言,日糧添加丁酸梭菌后瘤胃中厚壁菌門、梭菌屬和纖維桿菌屬的相對豐度顯著提高,推測丁酸梭菌的添加可能會提高小尾寒羊對纖維素的分解效率,為肌肉脂肪酸合成提供更多的營養和能量,有利于肌內脂肪的沉積,但其具體作用機制還需進一步研究。
2.6 瘤胃優勢菌群與脂肪酸組成的相關性分析
為了進一步探究瘤胃優勢菌群與背最長肌中脂肪酸組成之間的關系,以屬水平上相對豐度前10的優勢菌群與11 種脂肪酸進行Pearson相關性分析。由圖3可知,副擬桿菌屬相對豐度與α-亞麻酸、二十碳五烯酸相對含量呈顯著負相關(P<0.05),與飽和脂肪酸相對含量呈顯著正相關(P<0.05)。瘤胃桿菌屬相對豐度與硬脂酸、亞油酸和γ-亞麻酸相對含量呈顯著負相關(P<0.05),與α-亞麻酸和二十二碳六烯酸相對含量呈顯著正相關(P<0.05)。而普雷沃氏菌屬相對豐度與二十碳五烯酸和單不飽和脂肪酸相對含量呈顯著正相關(P<0.05),與硬脂酸飽和脂肪酸相對含量呈顯著負相關(P<0.05)。瘤胃桿菌屬和普雷沃氏菌屬主要參與纖維素的降解,可產生大量的揮發性脂肪酸,揮發性脂肪酸被瘤胃壁吸收,轉化生成脂肪酸,這可能是丁酸梭菌增加肌肉不飽和脂肪酸含量的內在機制[49]。丁酸弧菌屬相對豐度與棕櫚油酸和多不飽和脂肪酸相對含量呈顯著負相關(P<0.05)。此外,乳酸桿菌屬相對豐度與油酸、α-亞麻酸、二十碳五烯酸和單不飽和脂肪酸相對含量呈顯著正相關(P<0.05),但與亞油酸和飽和脂肪酸呈顯著負相關關系(P<0.05)。劉婷等[54]研究發現乳酸桿菌屬的添加能顯著提高蘇尼特羊背最長肌中油酸的含量,顯著降低亞油酸的含量,本研究結果與其相似。

圖3 瘤胃優勢菌群與脂肪酸組成相關性網絡圖Fig.3 Correlation network between predominant rumen flora and fatty acid composition
3 結論
本研究結果表明,日糧添加丁酸梭菌提高了小尾寒羊背最長肌中油酸、α-亞麻酸和二十碳五烯酸的相對含量,降低了棕櫚酸和硬脂酸的相對含量,提升了肌肉的肌內脂肪含量、色澤和嫩度,延緩了宰后pH值的下降,對肉品質有明顯的改善作用。通過脂肪代謝相關基因分析得出,丁酸梭菌組SREBP-1C、SCD、PPARγ和ACC基因表達水平顯著上調,從分子水平證實了丁酸梭菌可以促進機體脂肪沉積。進一步研究瘤胃微生物對脂肪酸代謝的影響發現,飼喂丁酸梭菌提高了小尾寒羊瘤胃中厚壁菌門、梭菌屬和纖維桿菌屬的相對豐度,降低了變形菌門、丁酸弧菌屬和副擬桿菌屬的相對豐度。瘤胃微生物與脂肪酸的相關性分析中,瘤胃中瘤胃桿菌屬、普雷沃氏菌屬和乳酸桿菌屬相對豐度與不飽和脂肪酸相對含量呈正相關。綜上,通過飼喂益生菌調控反芻動物瘤胃微生物影響機體脂肪酸代謝或是將來改善羊肉品質的重要研究方向,可為微生物制劑改善肉質性狀提供新的機理解釋。
