不同生物酶協同植物乳桿菌發酵對紫甘薯生全粉理化特性的影響
2023-10-20 13:36:26鄒浩峰廖雨華黃師榮施建斌
中國糧油學報 2023年8期
關鍵詞:植物
鄒浩峰, 廖雨華, 黃師榮, 隋 勇, 熊 添,施建斌, 蔡 沙, 蔡 芳, 梅 新
(湘潭大學化工學院1,湘潭 411105) (湖北省農業科學院農產品加工與核農技術研究所2,武漢 430064) (重慶文理學院園林與生命科學學院3,重慶 402168)
紫薯[Ipomoeabatats(L.)Lam]又名紫心甘薯、紫肉甘薯或黑薯,是甘薯的一個特殊品種,富含鐵、鋅、硒等多種礦物元素,含有18種氨基酸和多種維生素[1]。紫甘薯全粉是紫甘薯的主要深加工產品,基本保留了紫甘薯的營養價值和天然風味,是一種優質的食品原料。紫甘薯熟全粉是對新鮮紫甘薯進行熟化處理,制備過程比較繁瑣,加熱過程會使部分淀粉轉化為還原糖,產品干燥效率較低,淀粉已基本糊化完全,會影響熟全粉的加工特性。紫甘薯生全粉制備工藝比較簡單,原料沒有經過熟化處理,淀粉含量較高,還原糖含量較低,細胞完整性較好,最終產品的持水、持油性、凍融穩定性和膠凝性等加工特性較紫甘薯熟全粉較好,此外,生全粉的糊化溫度較低,熱穩定性差,凝沉性強且不易老化[2]。
發酵是一種常見的食品加工方式,被用來提高食品的營養和風味。發酵過程可以產生多種高活性的酶,如糖苷酶、木聚糖酶、植酸酶等,可以將淀粉、蛋白質、纖維素等大分子物質水解為氨基酸、寡糖等小分子物質,有利于消化吸收,并增加食品中一些結合狀態的功能成分如多酚等的釋放[3],被廣泛運用于如面包、饅頭等淀粉類加工產品。發酵過程就是微生物生長繁殖過程,在此過程中微生物會向外界釋放多種酶類物質,通過這些酶將環境中的大分子轉化為易于吸收的小分子為其提供支撐生命活動的碳源和氮源[4]。Sun等[5]研究發現植物乳桿菌和酵母協同發酵使低分子量淀粉短鏈含量和結晶度下降,面團的抗回生能力、彈性和強度上升。Xu等[6]研究發現植物乳桿菌發酵使淀粉顆粒表面明顯出現麻點和凹陷,增加了馬鈴薯淀粉的凝膠硬度和咀嚼性。王宏茲等[7]采用植物乳桿菌發酵紫薯粉制作的面包顏色鮮艷,具有獨特芳香,比未添加紫薯粉的面包具有更高的抗氧化性和花青素含量。牛萌萌等[8]發現利用米根霉和乳酸菌混合發酵大麥仁發酵產品具有淡淡的酒香味,酸甜可口,抗氧化性也大幅提高。
目前研究主要是利用單一菌種如乳酸菌發酵淀粉,研究淀粉的理化特性和開發新產品,添加不同生物酶協同植物乳桿菌發酵的研究較少,特別是對紫甘薯生全粉的研究鮮有報道。因此研究使用紫甘薯生全粉為原料,利用植物乳桿菌(Lactobacillusplantarum)、植物乳桿菌+纖維素酶(Cellulase)+普魯蘭酶(Pullulanase)、米根霉(Rhizopusoryzae)、植物乳桿菌+纖維素酶+普魯蘭酶+米根霉4種方式發酵,研究不同發酵方式和時間對紫甘薯生全粉基本成分、理化性質和熱特性的影響,以期為開發高質量的紫甘薯發酵產品提供參考。
1 材料與方法
1.1 材料與試劑
紫甘薯生全粉(日本凌紫);植物乳桿菌;米根霉;普魯蘭酶(1 000 U/g)、纖維素酶(15 000 U/g)、馬鈴薯葡萄糖瓊脂、MRS瓊脂、平板計數瓊脂,BR;其余化學試劑均為分析純。
1.2 儀器與設備
DSC 200 差示掃描量熱儀,BCM-1000 生物潔凈工作臺,CF-100A 發酵箱,SHP-080 生化培養箱,YM-75 壓力蒸汽滅菌器,GZX-9240 MBE電熱鼓風干燥箱,SHA-B 水浴恒溫振蕩器,LGJ-25C 冷凍干燥機,UV-2800 紫外可見分光光度計,QJ-08 400 g 多功能粉碎機,TGL-24MC 臺式高速冷凍離心機,FE20實驗室pH計,SKD-08S2 紅外智能消化爐,K9840自動凱氏定氮儀,TE214S 電子分析天平。
1.3 方法
1.3.1 發酵凍干粉的制備
植物乳桿菌發酵(Lac):在無菌條件下將凍干粉在MRS斜面培養基上37 ℃活化24 h,挑取單菌落于10 mL MRS液體培養基中進行二次活化,24 h后取1 mL菌液于MRS液體培養基,37 ℃振搖培養至菌液濃度為108CFU/mL。將所得菌液于4 ℃ 3 000 r/min條件下離心20 min,用生理鹽水洗滌沉淀3次,將所得菌泥與無菌水按1∶10(質量體積比)的比例混勻后按紫甘薯生全粉質量的8%接種,水質量分數為71%。
植物乳桿菌+纖維素酶+普魯蘭酶發酵(Lac/Cel-Pul):將植物乳桿菌(質量分數為8%)、纖維素酶(30 U/g)和普魯蘭酶(20 U/g)混合均勻后接種于紫甘薯生全粉中。
米根霉發酵(Rhi):在無菌條件下于PDA平板培養基上劃線接種米根霉,30 ℃條件下培養至長滿孢子,用150 mL無菌水將孢子沖洗至250 mL錐形瓶后,振搖15 min后于無菌操作臺上用紗布過濾,用100 mL無菌水沖洗紗布,即獲得米根霉孢子懸濁液。將所得菌液按紫甘薯生全粉質量的8%接種。
植物乳桿菌+纖維素酶+普魯蘭酶+米根霉發酵(Lac/Cel-Pul/Rhi):將植物乳桿菌(質量分數為4%)、纖維素酶(30 U/g)、普魯蘭酶(20 U/g)和米根霉(質量分數為4%)混合均勻后接種于紫甘薯生全粉中。
參考徐忠等[9]的方法,稍作修改。將各菌液與紫薯生全粉混合均勻后于37 ℃、pH 6.0、85%濕度條件下發酵,選取發酵時間為3、6、9、12 h的樣品,凍干后粉碎過100目篩保存備用。
1.3.2 基本成分測定
發酵紫甘薯生全粉水分、灰分、脂肪、蛋白質、淀粉、膳食纖維含量分別參考GB/T 5009.3—2016、GB/T 5009.4—2016、GB/T 5009.6—2016、GB/T 5009.5—2016、GB 5009.9—2016、GB 5009.88—2014。
可溶性糖含量參考王佩芬等[10]的方法,稍作修改。稱取1 g發酵粉,加入10 mL 80%(體積分數)乙醇,60 ℃水浴振蕩30 min,冷卻后定容至50 mL。取濾液0.1 mL,以蒸餾水作空白,采用硫酸-蒽酮比色法測定。葡萄糖標準曲線為:y=14.058x-0.178 5,R2=0.999。式中:y為吸光度,x為葡萄糖質量濃度(mg/mL)。根據標準曲線計算可溶性糖含量。
1.3.3 pH和總酸度(TTA)的測定
參考王宏茲等[7]的方法。取1 g發酵粉,加入9 mL純水,渦旋混勻后靜置10 min,測定溶液的pH;以0.1 mol/L的NaOH溶液滴定該懸濁液pH為8.5,所消耗的NaOH的體積即為發酵粉的TTA值。
1.3.4 直鏈淀粉含量的測定
參考何潔等[11]的方法采用雙波長法測定直鏈淀粉含量。
1.3.5 總酚的測定
參考杜俊娜等[12]的方法測定總酚含量。沒食子酸標準曲線為:y=10.187x+0.040 3,R2=0.998 8。式中:y為吸光度,x為沒食子酸質量濃度(mg/mL)。根據沒食子酸標準曲線換算出總酚含量。
1.3.6 DPPH自由基清除能力的測定
參考王宏茲等[7]的方法測定DPPH自由基清除能力。
1.3.7 花青素含量的測定
參考Meng等[13]的方法測定花青素含量。
1.3.8 色澤測定
將樣品平鋪在平皿中,采用色彩色差計分別測定各樣品的L*(明暗度)、a*(紅綠色)、b*(黃藍色)值。計算與未發酵樣品的色差ΔE,計算公式為:
1.3.9 水/油吸附能力測定
參考Yousif等[14]的方法測定吸水/油性。
1.3.10 吸水膨脹性測定
參考郭亞姿等[15]的方法測定吸水膨脹性。
1.3.11 熱力學特性分析
參照Pham等[16]的方法測定發酵粉熱力學特性。稱取(5.0±0.1)mg樣品至鋁坩堝中,加入10 μL去離子水,加蓋密封,于室溫下平衡2 h后采用DSC分析其熱力學特性,空坩堝作為參比。測定參數為:以10 ℃/min升溫速率從20 ℃升溫至 120 ℃后以40 ℃/min速率從120 ℃迅速降至20 ℃,氮氣流量20 mL/min,根據吸熱曲線分析計算得糊化起始溫度(To)、峰值溫度(Tp)、終止溫度(Tc)及糊化焓(ΔH)。
1.4 數據處理
所有實驗均重復3次,結果以平均數±標準差表示;采用Excel和SPSS 26.0分析處理數據,Origin 2017繪圖。
2 結果與分析
2.1 發酵對紫甘薯生全粉基本成分的影響
由表1可見,發酵使紫薯生全粉淀粉含量下降,植物乳桿菌參與的發酵淀粉含量下降更為明顯,這可能是因為淀粉酶和有機酸加速了對淀粉的分解[17],Rhi發酵的淀粉水解程度更低,可能是因為其酸度變化最弱。植物乳桿菌參與的3種發酵方式的蛋白質含量升高,這可能是因為植物乳桿菌使用紫甘薯生全粉作為碳源和能源產生的真菌菌絲體蛋白使蛋白質含量升高[18]。發酵3 h后,Lac-Cel/Pul-Rhi脂肪含量顯著降低,除Lac發酵外,其余發酵方式灰分含量均顯著上升。乳酸菌能在一定程度上降解纖維素[19],并且在酶的作用下降解的更顯著。Rhi發酵12 h后SDF含量要明顯小于Lac/Cel-Pul/Rhi發酵,這表明米根霉和植物乳桿菌對膳食纖維有明顯的協同作用,單純的生物酶處理效果遠遜于植物乳桿菌與生物酶共同作用[20]。
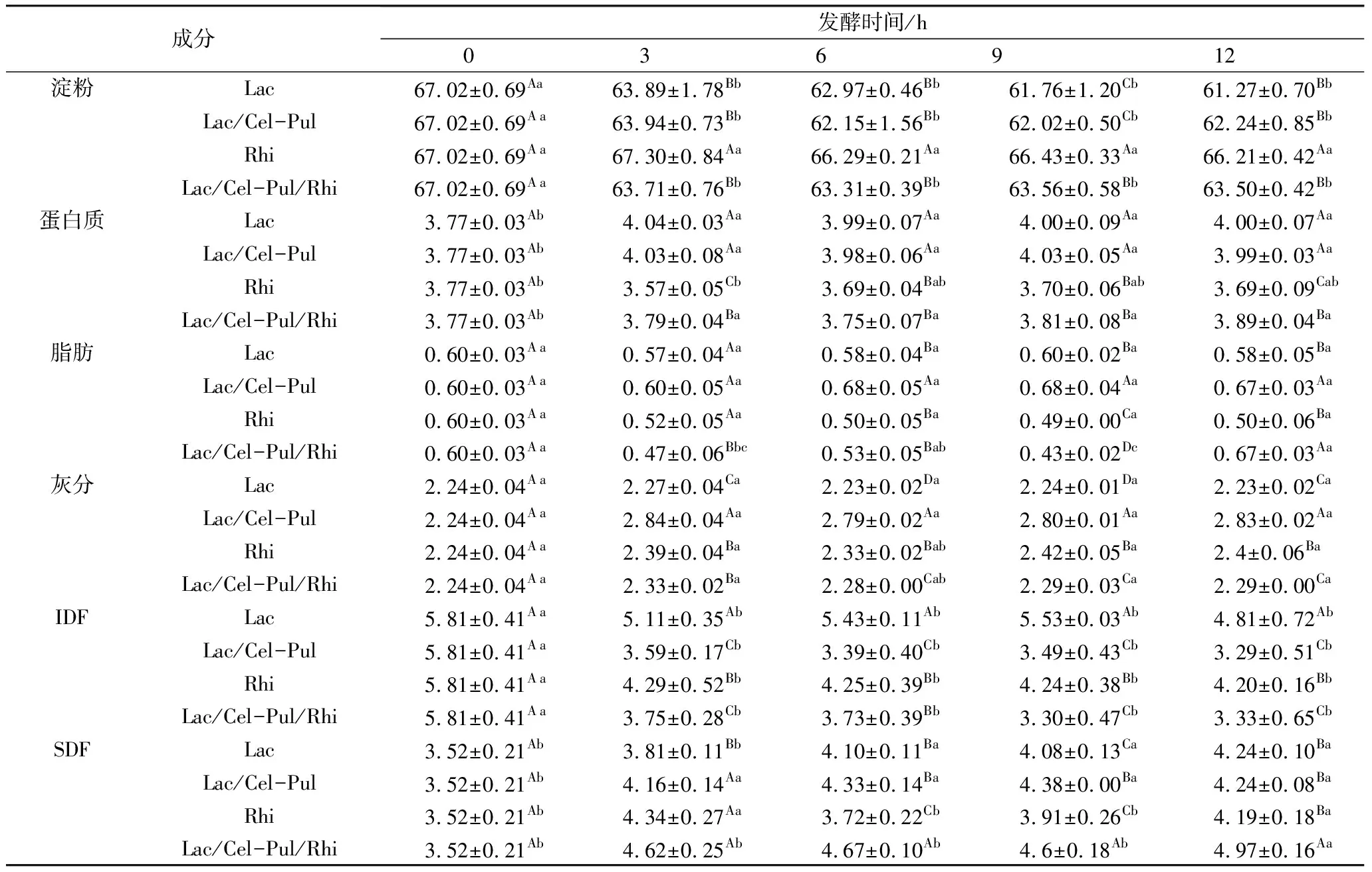
表1 發酵前后紫甘薯生全粉主要組成成分(干基)質量分數/%
由圖1可見,4種發酵方式的可溶性糖含量在0~3 h內快速上升,在3 h后呈下降趨勢。在0~3 h內植物乳桿菌和米根霉迅速繁殖,向環境內分泌大量的淀粉酶和糖化酶等活性物質,使淀粉含量下降,可溶性糖含量上升;3~9 h內,由于可溶性糖含量達到一定濃度且有機酸的積累開始抑制酶類物質的釋放,植物乳桿菌和米根霉的生長速度減慢,部分可溶性糖被微生物消耗以在后期發酵中繁殖和代謝;9 h后,由于大量的有機酸等有害物質的積累,使得酶類物質的活性被嚴重抑制,植物乳桿菌等的生長能源只能依靠可溶性糖供給,因此可溶性糖含量快速下降。此外,多種菌混合發酵可溶性糖含量比單一菌種增加的更明顯,這可能是因為真菌產生的淀粉酶、菊粉酶、纖維素酶等多種生物活性酶更多,以及大分子淀粉轉化為更多的小分子糖有關[21]。

圖1 發酵前后紫甘薯生全粉可溶性糖含量的變化
2.2 發酵對紫甘薯生全粉pH和TTA的影響
如圖2所示,不同發酵方式的產酸能力差異性顯著(P<0.05)。在發酵過程中生物酶能利用碳水化合物產生大量乳酸,使紫甘薯生全粉的pH降低,同時也能抑制有害微生物的生長繁殖[22]。4種不同發酵方式的紫甘薯生全粉在3 h內酸度明顯上升,3~9 h內酸度呈緩慢上升趨勢,在 9 h后酸度劇烈上升,pH的變化也與此現象相互吻合。在第12 h,除Lac/Cel-Pul/Rhi發酵外,其余發酵pH均在5.0以下,其中Lac發酵產酸能力最強。4種發酵方式pH在3 h內迅速下降,這是因為在初始發酵階段生長環境適宜且營養豐富,使得植物乳桿菌和米根霉快速生長繁殖,產生大量乳酸。3~9 h內Lac/Cel-Pul、Rhi、Lac/Cel-Pul/Rhi發酵進入平穩期,pH緩慢下降,這可能是因為酶的添加促使紫甘薯中緩沖物質的釋放,從而保證溶液體系的穩定。9 h后pH迅速下降是由于乳酸不斷積累,逐漸破壞體系穩定性。不同發酵產酸能力 Lac/Cel-Pul>Lac>Lac/Cel-Pul/Rhi>Rhi,通過對比可以明顯發現植物乳桿菌在發酵初期就成為優勢主導菌,并且有酶參與的發酵組酸度下降更慢[20]。
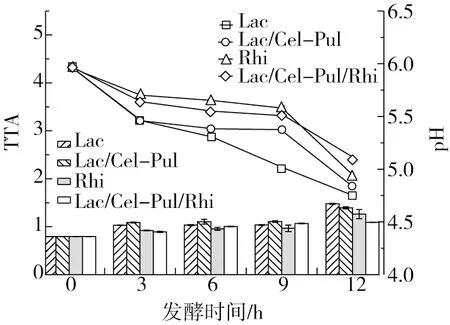
圖2 發酵對紫甘薯生全粉pH和酸度的影響
2.3 發酵對紫甘薯生全粉直鏈淀粉含量的影響
由表2可知,不同發酵方式使紫甘薯生全粉直鏈淀粉含量下降,但均無顯著性差異(P>0.05),結合淀粉含量變化情況分析,雖然直鏈淀粉含量在下降,但直鏈淀粉與支鏈淀粉的比率在不斷上升。有研究報道發酵過程中的微生物產生的酶和酸會水解支鏈淀粉,提高直鏈淀粉的比例,多種菌種混合發酵時優勢微生物越多,產生的酶和酸就越多,直鏈淀粉增加的更明顯[23]。
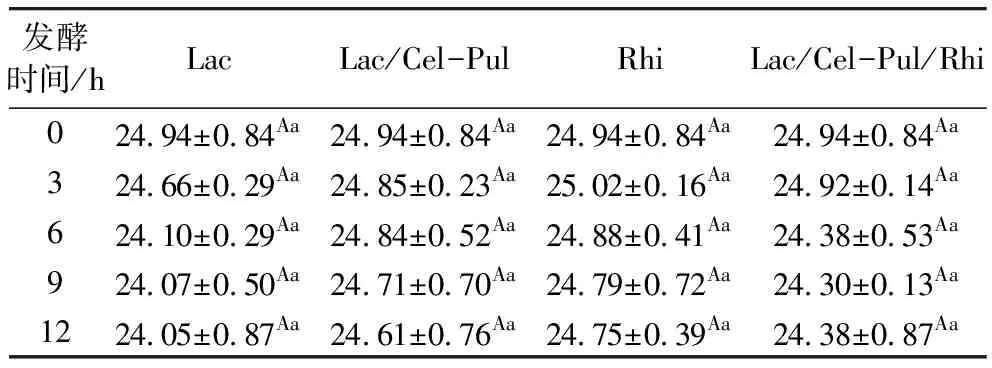
表2 發酵前后紫甘薯生全粉中直鏈淀粉質量分數(干基)/%
2.4 發酵對紫甘薯生全粉總酚含量和DPPH自由基清除率的影響
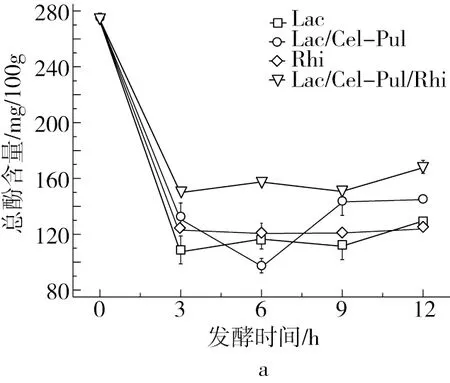
由圖3可知,紫甘薯生全粉具有較高總酚含量,發酵使樣品的總酚含量降低,Hector等[24]研究指出乳酸菌發酵降解了某些酚類物質,這與本研究結果一致。3 h后總酚含量有上升的趨勢,可能是在發酵時產生的一些酶使細胞結構結合的多酚物質得以釋放,從而提高總酚含量[25]。植物乳桿菌和米根霉單獨發酵在12 h后總酚含量較為接近,但略低于混合菌發酵。不同發酵方式均會提高紫甘薯生全粉的抗氧化活性,發酵12 h時DPPH自由基清除能力大小為Lac/Cel-Pul>Lac>Lac/Cel-Pul/Rhi>Rhi,但差異不顯著。有研究報道乳酸菌發酵會使一些酚類物質降解為抗氧化性更強的物質[7],從而提高其抗氧化能力。此外,可以明顯看出米根霉參與的發酵DPPH自由基清除能力要略低一些,可能是因為米根霉的參與抑制了部分酚類物質降解。
注:不同大寫字母表示同一發酵時間不同發酵方式的差異顯著(P<0.05),不同小寫字母表示同一發酵方式不同發酵時間的差異顯著(P<0.05),余同。
2.5 發酵對紫甘薯生全粉花青素含量的影響
如圖4所示,不同發酵方式對花青素含量的影響顯著。研究發現乳酸菌發酵對花青素有一定的降解作用[7],在本實驗中的降解作用比較明顯,0~3 h內所有發酵花青素含量劇烈下降。與總酚含量變化情況相同,添加米根霉的發酵對花青素的有影響較小,初步判斷可能是米根霉的加入使植物乳桿菌分泌的一些微生物受到抑制,因此對花青素含量影響較小,但這需要進一步去研究論證。此外,隨著發酵的進行,體系的pH降低,酸度變大,花青素的穩定性提高[26],因此幾種發酵的花青素含量下降趨勢變緩。Pretorius等[27]指出在發酵時一些微生物的次級代謝活動的產物可以與花青素結合成一種復合物,進而提高花青素物質的穩定性,這或許是花青素含量在發酵后期提高的原因。
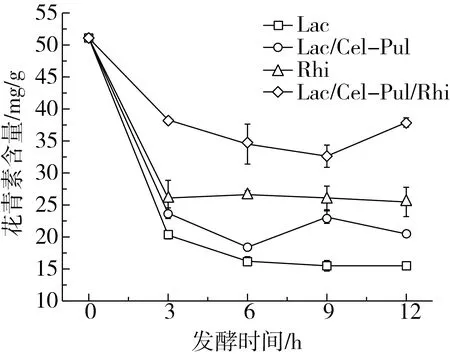
圖4 發酵對紫甘薯生全粉花青素含量的影響
2.6 發酵對紫甘薯生全粉色澤的影響
色差值越小證明與原樣品顏色越接近,當色差值大于3時可以用肉眼分辨出來。由表3可見,所有發酵方式對紫甘薯生全粉的色澤影響較大,明顯可以用肉眼分辨。隨著發酵時間的延長,色差值有一個先上升后下降的過程,其中Lac發酵對紫甘薯生全粉的色澤影響最明顯,發酵6 h時色差值達到最大。色澤變化情況可能與酸度變化有關,12 h時色差值變化情況為Lac>Lac/Cel-Pul>Lac/Cel-Pul/Rhi>Rhi,與pH變化情況一致(圖2)。王宏茲等[7]指出主要花青素的顯色會根據溶液pH的變化而變化,pH較低時為紅色,隨著pH的升高溶液逐漸變為淺紫色。
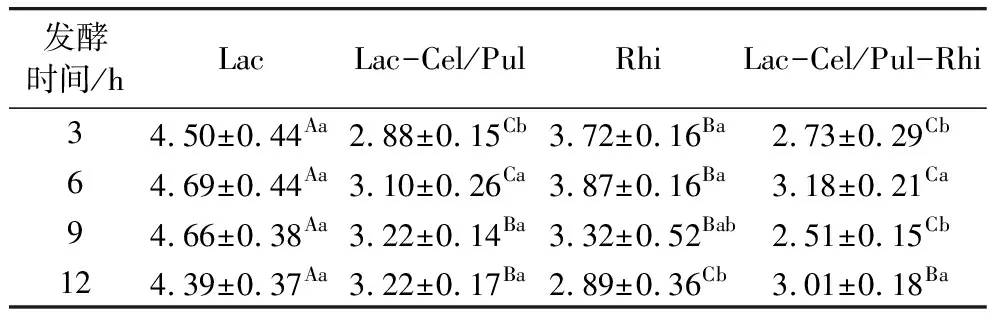
表3 發酵對紫甘薯生全粉色澤的影響
2.7 發酵對紫甘薯生全粉水合特性的影響
由圖5可見,發酵后紫甘薯生全粉的持水率和吸水膨脹性顯著下降,尤其是米根霉參與的發酵下降的更為明顯,持油率隨發酵時間的延長而增大,混合菌種的效果更明顯。胡畔等[19]研究發現發酵使一些復合大分子在某些連接處斷裂生成小分子,小分子的含量逐漸增大,而持水率的測定需要經過離心等操作,離心時小分子物質無法沉淀在底部而隨上清液倒出,從而使發酵后玉米粉的持水率降低。混合菌的發酵加劇了此類小分子的生成,因此混合發酵的持水率、吸水膨脹性下降的更為嚴重。徐忠等[9]使用植物乳桿菌液態發酵玉米粉,發現持水率的降低和蛋白質、淀粉分子結構被破壞有關。持油率的上升可能是SDF含量的增加導致,Li等[28]研究發現發酵使SDF的比表面積增加,形成松散空洞的結構,更多非極性基團的暴露也可能使持油率上升。
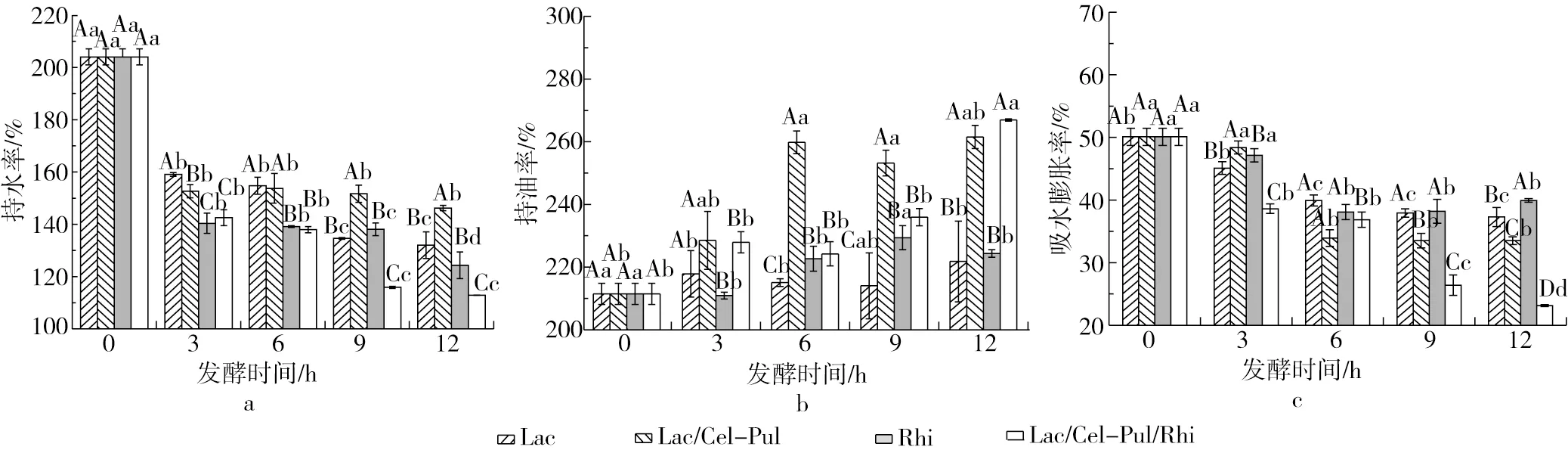
圖5 發酵對紫甘薯生全粉持水、持油、吸水膨脹性的影響
2.8 發酵對紫甘薯生全粉熱力學特性的影響
由表4可見,與未發酵組相比,發酵組紫甘薯生全粉ΔH、Tc顯著提高,To顯著降低,Tp變化不顯著,但有明顯下降的趨勢,特別是植物乳桿菌參與的發酵效果明顯優于米根霉發酵,混合菌種的發酵效果顯著優于單一菌種發酵。王東坤等[29]指出發酵過程產生的酶和酸能夠水解或破壞淀粉的無定型區,水分子更易進入淀粉顆粒內部,從而使淀粉更易糊化,To降低。此外,由于淀粉的無定形區被破壞,其比例有所降低,結晶區比例相對增加,造成ΔH升高,Lac/Cel-Pul/Rhi發酵12 h時ΔH顯著高于其余發酵。莫琰等[23]指出普魯蘭酶對淀粉無定形區和不完美結晶酶解后,剩余的完美結晶需要更高的溫度才能熔融。此外,溶出的直鏈淀粉與脂類物質形成更為復雜的雙螺旋結構,糊化所需的熱量隨之上升。直鏈淀粉相對含量上升可能會減弱淀粉結晶結構在糊化過程中受到的破壞,因此直鏈淀粉含量的增加可能也可能是造成ΔH升高的原因。
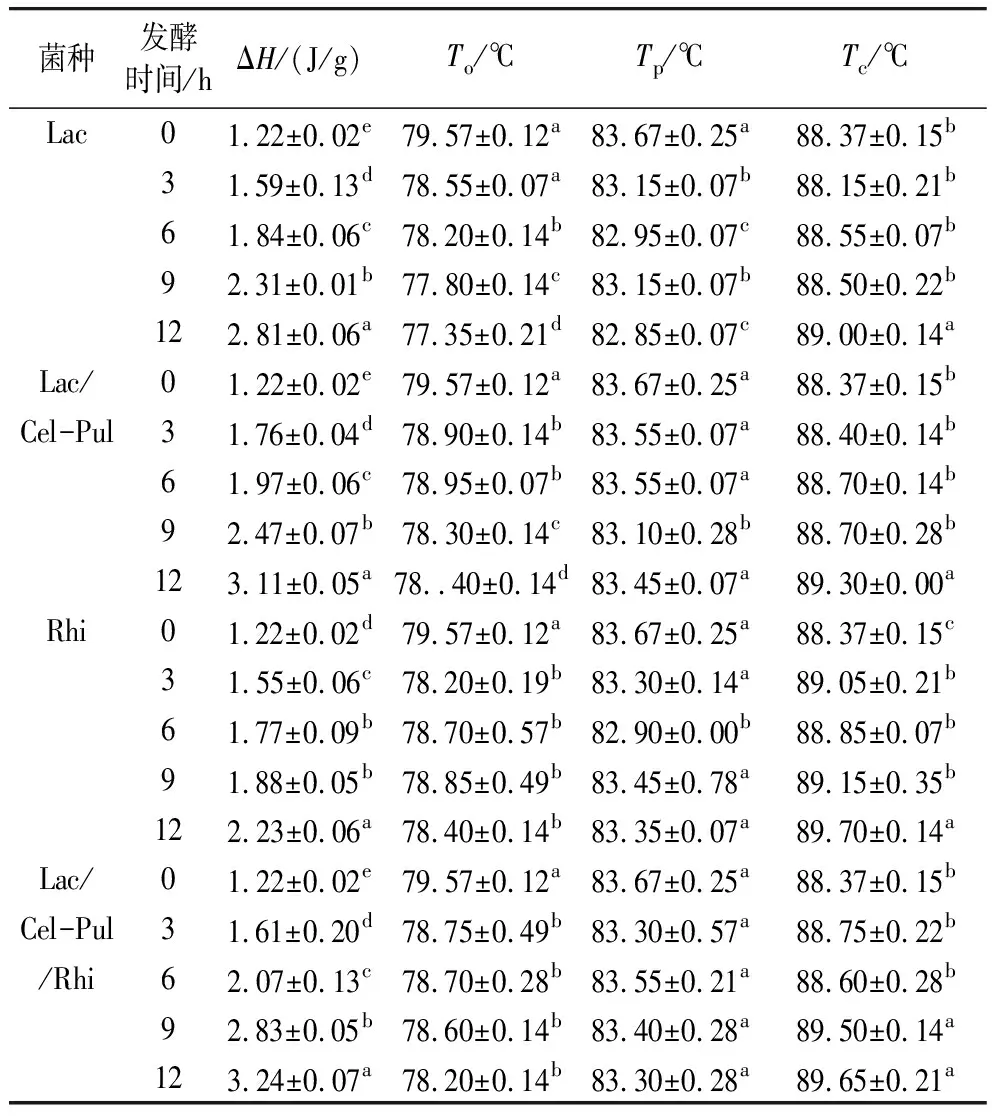
表4 發酵對紫甘薯生全粉熱力學特性的影響
3 結論
研究不同生物酶協同植物乳桿菌發酵對紫甘薯生全粉基本成分、理化特性、色澤和熱特性的影響。結果表明,Lac/Cel-Pul/Rhi發酵品質最好,SDF、可溶性糖、總酚、花青素、持油性明顯高于其他發酵組,但持水性、吸水膨脹性較差,熱特性結果顯示其ΔH最高,To提前,Tc推遲,具有良好的熱特性。紫甘薯生全粉顏色變化與產酸能力變化情況一致,由強到弱依次為Lac/Cel-Pul>Lac>Lac/Cel-Pul/Rhi>Rhi,這是因為花青素在不同pH條件下顯色不同造成紫甘薯生全粉色澤的差異。此外,不同發酵方式均使紫甘薯生全粉抗氧化能力升高。不同生物酶協同植物乳桿菌發酵得到的紫甘薯生全粉品質較好,有利于發酵在紫甘薯生全粉工業的應用和高品質紫甘薯生全粉產品的開發。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
