小麥分蘗數目遺傳研究進展與展望
2023-10-11 08:46:40申雪懿李東升曲朝喜姚維成劉家俊溫明星
麥類作物學報 2023年10期
申雪懿,李東升,陳 琛,曲朝喜,郭 瑞,姚維成,劉家俊,鄧 垚,溫明星
(江蘇丘陵地區鎮江農業科學研究所,江蘇句容 212400)
小麥是中國三大糧食作物之一。近年來由于產業結構供給側調整,小麥播種面積不斷下降,尤以春麥區、北部冬麥區和西南冬麥區下降最為嚴重[1]。為保障國家糧食安全以及滿足居民糧食需求,提高單位面積產量成為小麥育種工作的重中之重。
小麥分蘗數目影響單位面積小穗數的形成,是構成小麥產量的重要因素之一。作為一個復雜的數量性狀,分蘗數目同時由多個主效基因和微效基因控制。挖掘控制分蘗數目的關鍵基因,解析影響分蘗數目的分子調控網絡,有利于揭示分蘗發育的遺傳機制和分子機理,為小麥高產育種提供堅實的理論基礎和有效的基因資源。隨著基因組測序拼裝技術和生物信息學算法的快速進步,為小麥分蘗數目相關數量位點(quantitative trait locus, QTL)的定位及基因克隆奠定了重要基礎。本文對小麥分蘗發育過程、分蘗數目相關基因/QTL的定位以及相關基因的克隆三方面進行綜述,以期為小麥分蘗數目QTL進一步精細定位、優異等位基因挖掘和分子標記輔助選擇高產育種提供理論依據。
1 小麥分蘗的發育過程
不同作物之間由于馴化選擇方向的不同,分蘗的生長發育也隨之不同。在小麥中,多分蘗性狀被保留了下來;而在玉米中,多分蘗性狀在馴化和選擇中丟失,分蘗變少,使營養主要集中在主莖上[2]。分蘗起源于腋生分生組織,其產生和發育主要經歷兩個階段:(1)腋生分生組織發育,即在莖尖和葉原基之間形成邊界后,腋生分生組織開始在葉腋近軸端的分生組織細胞中形成;(2)腋芽伸長形成分蘗[3]。邊界的形成和分生組織的發育受遺傳背景、激素和環境的多重影響[4-7]。此外,腋芽的伸長也受多種因素的控制,除自身遺傳因素外,也受腋生分生組織活性、分蘗芽生長位置、營養利用情況、光照以及溫度等各種外在因素的影響[8-11]。
小麥分蘗的生長發育通常遵循一定的規律(圖1)。在成熟小麥胚中,胚芽鞘的腋部產生一個腋芽,此腋芽通常處于休眠狀態,稱為T0;此外,第一葉原基的腋部也產生另一個腋芽,稱為T1;當第三葉充分伸長,第四葉出現時,T1開始發育;從T1葉腋產生的次級分蘗稱為T1.1,從莖基部先出葉產生的次級分蘗則稱為T1.0和T2.0,并以此類推[12]。但在實際生產中,由于各種外在因素的影響,小麥不會無限分蘗,只有在條件良好的情況下,才可以生成三級分蘗和四級分蘗。在前期營養生長階段,主莖和次級分蘗的產生決定了小麥總分蘗數目。進入生殖生長階段后,植物體內的營養物質主要集中在主莖,而其他分蘗獲得的營養物質較少。不同分蘗獲得的營養物質也不同,其中擁有獨立營養供給系統的四葉分蘗和大分蘗能夠獲得足夠的營養進行抽穗結實,成為有效分蘗,而沒有獨立營養供給系統的小分蘗會逐漸死亡,成為無效分蘗[13-15]。小麥除無效分蘗和有效分蘗之外,還存在動搖分蘗,即三葉分蘗。外界生長環境、耕作栽培方式等不確定因素使三葉分蘗無法獲得穩定的營養供給,常常在無效分蘗和有效分蘗之間轉換[16]。

C:胚芽鞘;L:葉;T:分蘗.C:Coleoptile; L:Leaf; T:Tiller.圖1 小麥分蘗結構示意圖[12]Fig.1 Diagram of wheat tillering structure[12]
2 控制小麥分蘗數目的遺傳位點
分蘗抑制突變體及其近源屬種為小麥分蘗發育機制的研究提供了理想材料。迄今為止,采用分蘗抑制突變系及其近源屬種已定位到4個分蘗抑制基因tin1、tin2、tin3和ftin[17-20]。
tin1來自于Alpha和一個北非陸地種雜交衍生的單莖小麥系,位于1AS染色體上,為隱性基因,與SSR標記Xgwm136共分離[17]。tin2來自于面包小麥品系88F2185,為顯性基因,位于2A染色體上[18]。tin3來自于一個分蘗力受損的二倍體小麥(T.monococcumsubsp.)單莖突變體,位于3AL染色體上,與共顯性標記Xpsr1205緊密連鎖[19]。ftin來自于普通小麥與冰草雜交獲得的可育分蘗抑制系Pubing 3558,位于1AS染色體上,與標記Xcfa2153緊密相連[20]。
連鎖分析是進行作物復雜數量性狀遺傳解析的重要方法。前人利用雙親構建的RIL群體,在除1D、3A和5B之外的其他18條染色體上均定位到了控制小麥分蘗數目的QTL,可解釋表型變異0.74%~55.40%,其中在兩個及以上環境下能被檢測到的基因/QTL信息如表1所示。
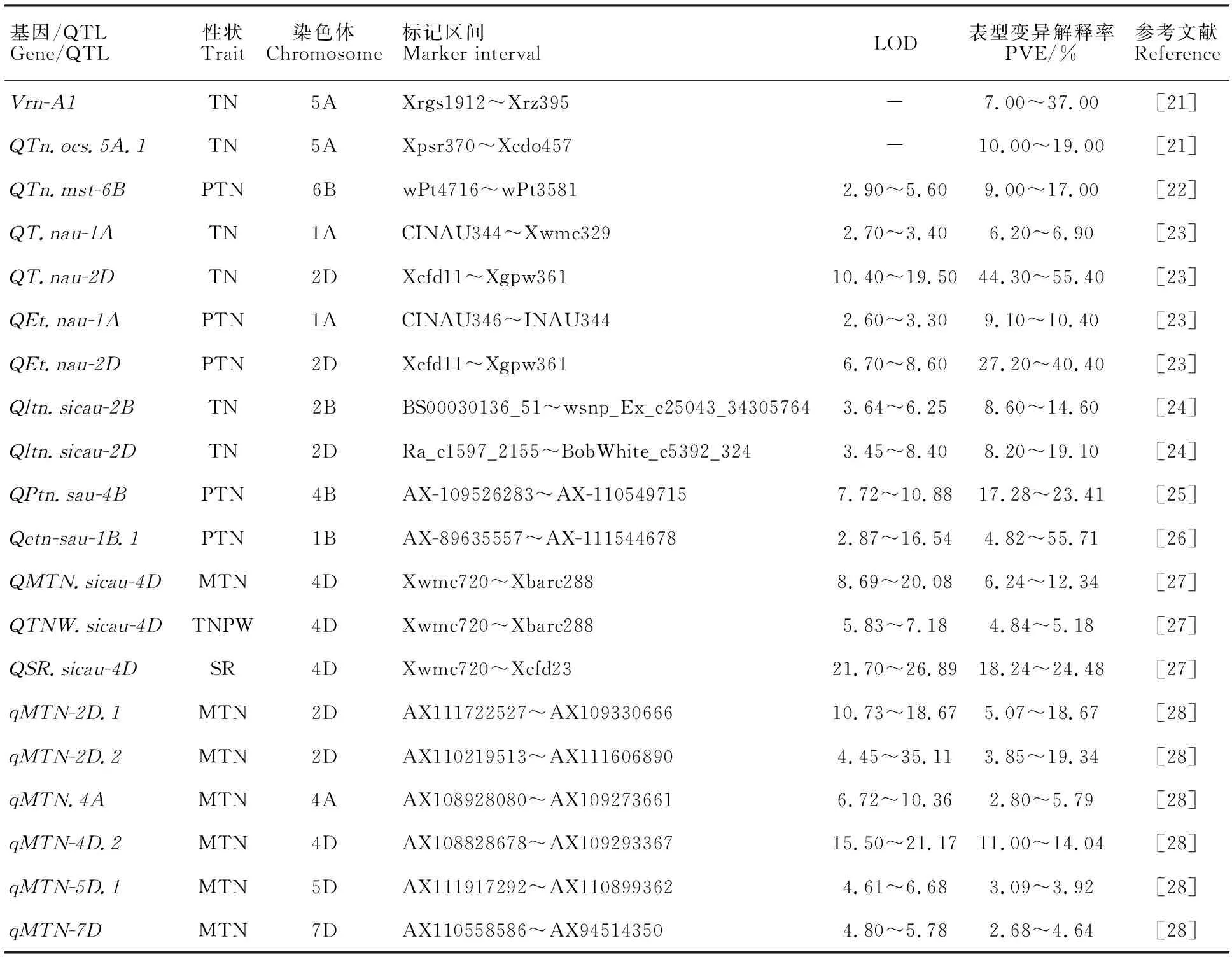
表1 已知控制小麥分蘗數目的遺傳位點Table 1 Genetic loci controlling tiller number in wheat
Kato等[21]利用Cappelle-Desprez和中國春雜交衍生的RIL群體為材料,在5A染色體上鑒定到兩個控制單株分蘗數目的穩定遺傳位點,其中QTn.ocs.5A.1位于分子標記Xpsr370和Xcdo457之間,Vrn-A1處于分子標記Xrgs1912和Xrz395之間。Naruoka等[22]利用Reeder和Conan雜交構建的RIL群體為材料,在6B染色體上鑒定到一個控制有效分蘗數目的穩定QTL,位于標記wPt4716和wPt3581之間,可解釋9%~17%的表型變異;該QTL同時在RIL群體McNeal/Thatcher和McNeal/Reeder以及近等基因系中均得到了驗證。Xu等[23]利用EMS誘變望水白得到的多分蘗突變株系NAUH167,將其與蘇麥3號進行雜交,對其后代衍生的RIL群體進行遺傳定位,在2D染色體上定位到一個同時控制單株分蘗數目和單株有效分蘗數目的穩定QTL,位于分子標記Xcfd11和Xgpw361之間。Wang等[24]利用H461和川農16雜交構建的RIL群體為材料,在2B和2D染色體上分別定位到兩個控制單株分蘗數目的穩定QTL,在多個遺傳背景下均具有顯著效應,其中Qltn.sicau-2B位于SNP標記BS00030136_51和wsnp_Ex_c25043_34305764之間;Qltn.sicau-2D位于SNP標記Ra_c1597_2155和BobWhite_c5392_324之間。Liu等[25]基于小麥55K芯片構建的全基因組遺傳連鎖圖譜,利用20828和川農16雜交構建的RIL群體為材料,在4B染色體上定位到一個在多環境下穩定表達的有效分蘗數目QTL,位于標記AX-109526283和AX-110549715之間。Liu等[26]基于小麥55K芯片、SSR標記和KASP標記構建的高密度遺傳連鎖圖譜,利用20828和SY95-71雜交構建的RIL群體為材料,在1BL染色體上定位到一個在多環境下穩定表達的有效分蘗數目QTL,位于標記AX-89635557和AX-111544678之間。Hu等[27]利用川農18和T1208構建的RIL群體,在4D染色體上發現一個同時控制單位面積最大分蘗數目和冬前單位面積分蘗數目的遺傳位點,在該位點附近還存在一個與成穗率有關的QTL;Ren等[28]則在該RIL群體的多條染色體上定位到控制早期單位面積分蘗數目、冬前單位面積分蘗數目、單位面積最大分蘗數目和單位面積有效分蘗數目的遺傳位點,其中控制單位面積最大分蘗數目的QTL在多個環境下能被檢測到。
3 小麥分蘗數目相關基因的克隆
目前,通過圖位克隆的方法還未在小麥中克隆到與分蘗數目相關的基因,但通過同源基因克隆的方法,已經克隆到8個與小麥分蘗數目相關的基因(TaMOC1、TaTB1、TaPAY1、TaPIN、TaD27、TaPIL1、TaSPL和TaD53)。這8個基因作用于小麥生長的不同生育階段,通過與不同遺傳因子或外源激素相互作用參與調控小麥分蘗。
3.1 TaMOC1基因
TaMOC1編碼一種GRAS家族核蛋白,可促使小麥腋芽形成并促進其生長[5],另外在小麥葉原基的表皮細胞、莖尖分生組織、小穗原基和幼葉中也有表達[29-30]。涂田莉[30]研究發現,TaMOC1基因可能參與無效分蘗的形成,在TaMOC1-RNAi轉基因T3代陽性株系中,越冬前期和拔節期的分蘗數目顯著少于野生型,但抽穗期的有效分蘗數目與野生型無顯著差異。杜麗君[31]通過對TaMOC1基因啟動子區域順式作用元件進行分析,發現TaMOC1基因可能通過與分生組織表達有關的順式調控元件參與調控小麥分蘗。
3.2 TaTB1基因
TB1是控制小穗發育和調節株型的主要基因,在不同作物中功能非常保守。TB1基因最先在玉米中被克隆,可抑制玉米側枝的發生,使玉米由祖先大芻草的多分枝性狀變成單莖性狀[32-34]。Lewis等[35]將玉米TB1基因轉化到小麥品種Bobwhite中,發現過表達TB1基因抑制了小麥分蘗的形成。Dixon等[36]研究表明,TaTB-B1和TaTB-D1基因可抑制小麥腋芽的形成進而減少分蘗數目,同時可促進小穗的發育。
3.3 TaPAY1基因
PAY1基因可調節作物株型和產量[37]。研究表明,TaPAY1基因在小麥莖尖分生組織、葉原基、小穗原基和幼葉中均有表達,其表達受脫落酸、赤霉素、茉莉酸、生長素以及干旱、低溫脅迫的調控,且拔節期和抽穗期TaPAY1-RNAi轉基因T2代株系的分蘗數目較野生型均有所增加[16]。
3.4 TaPIN基因
生長素是一種具有極性運輸特性的激素,在調控植物分蘗方面起重要的作用[38-41]。PIN蛋白是生長素運輸的特異性蛋白,是生長素極性運輸的限制因子[42-44]。目前在小麥中已鑒定出9個PIN1同源基因,其中TaPIN1-6基因在小麥莖尖和嫩葉中高表達[45]。在TaPIN1-RNAi轉基因小麥株系中,TaPIN1s基因下調表達,使植株的分蘗數目增加,表明生長素可能介導小麥腋芽的產生,且TaPIN1-RNAi株系的小穗數、每穗粒數和千粒重較野生型均有所下降[45]。
3.5 TaD27基因
D27基因是獨腳金內酯生物合成和信號轉導過程中的關鍵基因,在調控分蘗數目方面具有重要作用。獨腳金內酯是除生長素外調節分蘗數目的另一重要激素,其可通過抑制腋芽的伸長負調控分蘗數目[46]。TaD27基因編碼一種獨腳金內酯合成酶,進而影響小麥分蘗數目。TaD27-RNAi株系表現為獨腳金內酯含量降低和分蘗增多,而TaD27-B過表達株系表現為獨腳金內酯含量增多和分蘗減少[47]。此外,在水培條件下,GR24(合成獨腳金內酯模擬物)和TIS108(獨腳金內酯合成抑制劑)影響TaD27-RNAi株系和TaD27-B過表達株系的腋芽生長,表明TaD27-B通過參與獨腳金內酯的生物合成來調控小麥分蘗[47]。
3.6 TaPIL1基因
Zhang等[48]研究表明,TaPIL1可作為一種新的分蘗抑制因子,激活TaTB1基因的轉錄,進而抑制小麥分蘗;過表達TaPIL1基因可降低小麥分蘗數目,而過表達TaPIL1-SUPERMAN抑制結構域則可增加小麥分蘗數目。
3.7 TaSPL基因
SPL轉錄因子參與植物生長發育過程,在作物株型調控方面具有重要作用,其典型特征是含有一個可以被miR156識別的SBP-box結構域。小麥miR156通過對TaSPL基因進行轉錄切割來調控分蘗數目,miR156的過表達可抑制TaSPL3/17基因的表達,進而增加小麥分蘗數目[49]。
3.8 TaD53基因
TaD53是小麥獨腳金內酯信號通路的抑制因子,與轉錄共抑制因子TaTPL相互作用發揮功能。同時TaD53基因還可直接與miR156控制的TaSPL3/17蛋白相互作用,抑制TaTB1基因的表達,導致分蘗數目增加[49]。
4 研究展望
4.1 多角度多維度解析分蘗數目性狀
為全面解析小麥分蘗數目遺傳調控網絡,對分蘗數目的遺傳研究需要進一步細化和深入。在以往的研究中,單株分蘗數目和單株有效分蘗數目是研究的熱點[21-26],不同時期的單位面積分蘗數目在近幾年也有所涉及[27-28],而單位面積有效分蘗數目和成穗率則鮮有研究。在小麥生產中,單位面積有效分蘗數目和成穗率與最終產量顯著相關,增加單位面積有效分蘗數目在一定程度上可以提升單位面積的產量水平;降低無效分蘗比率可提高小麥分蘗成穗率,有利于減少資源消耗,進而增強小麥的個體競爭力。在后續研究中,對單位面積有效分蘗數目和分蘗成穗率相關基因/QTL的挖掘和克隆有待加強,為精準改良分蘗性狀和提高小麥單產水平提供參考。
4.2 利用基因組學技術促進小麥分蘗數目相關基因的克隆
前人利用同源基因克隆方法在小麥中克隆到多個與小麥分蘗數目相關的基因,但采用經典圖位克隆的方法還未克隆到相關基因,原因可能是小麥龐大的基因組阻礙了功能基因組研究的步伐。隨著高通量測序技術和基因組拼裝技術的不斷更新和發展,普通小麥[50]、祖先種[51-53]及其近緣屬種[54-56]的高質量參考基因組已相繼公布。小麥基因組測序技術的快速發展,加快了高密度SNP芯片的開發,為分子標記的開發和遺傳圖譜的構建提供了大量的遺傳多態性信息。此外,小麥突變體數據庫[57]、小麥表達譜數據庫[58]和小麥族同源基因數據庫[59]也已建立。基于上述豐富的基因組數據和轉錄組數據,在采用雙親群體進行遺傳連鎖定位的同時,可利用全基因組關聯分析(genome-wide association study, GWAS)和轉錄組關聯分析(transcriptome-wide association study, TWAS)進一步識別和確定候選基因,加快分蘗數目相關基因的挖掘進程。
4.3 利用基因編輯技術加快分蘗基因功能研究
普通小麥作為一種異源六倍體作物,大多數基因有三個拷貝,且同源基因之間可能存在功能冗余,這為基因功能驗證帶來了困難。利用RNA干擾可有效地沉默同源基因。近年來隨著小麥遺傳轉化效率的極大提高,基于CRISPR/Cas9系統的基因編輯技術在小麥基因功能研究及品種改良中取得了重要進展[60-62]。基因編輯技術不僅可以針對單個基因,還可以靶向三個等位基因或串聯基因簇,更加高效地進行基因功能分析,為揭示小麥分蘗的遺傳調控網絡和進行品種遺傳改良提供幫助。
