miR-891a-5p對結腸癌細胞腫瘤生物學行為和血管生成潛能的作用
2023-10-08 09:21:04康麗麗陳昭偉
河北醫學 2023年9期
康麗麗, 陳昭偉, 宋 健
(海南省腫瘤醫院, 海南 海口 570123)
美國癌癥協會公布的癌癥統計數據顯示,人類結腸癌的發病率為10.2%,死亡率已達到9.2%,從第四位上升到第二位[1],是消化系統中最常見的癌癥之一。通常,腫瘤切除適用于結腸癌的早期階段,化療藥物的組合通常用于不同階段的結腸癌患者,特別是在晚期[2]。隨著癌癥治療的發展,特別是免疫治療和靶向治療癌癥患者的總生存率有所提高[3]。然而,并非所有癌癥患者都能從這些治療中受益,為了解決這個問題,許多研究還探索了一系列分子特征,如基因組特征[4]和生物標志物[5],用于預測對治療的反應,從而指導個性化治療。MicroRNA是一類由19-22個核苷酸組成的小非編碼RNA,作為mRNA翻譯效率的負介質參與幾乎所有細胞過程,轉錄后的MicroRNA通過結合靶mRNA的3,非翻譯區(3,-UTR)中的序列來降低靶基因表達[6]。miRNA的調控功能已在細胞增殖、遷移、分化和侵襲等各種細胞過程中被發現,新興研究報道,功能性miRNA可直接作為抑癌基因或促癌基因,從而調節腫瘤進展[7]。腫瘤的生長、侵襲和遷移取決于血管生成,血管生成為腫瘤細胞提供足夠的氧氣和營養。因此,血管生成抑制劑開啟了癌癥治療的新時代,近年來,已被廣泛接受為一種臨床治療策略。本研究探討了miR-891a-5p作為潛在血管生成因子在結腸癌轉移中的作用。本研究考察了miR-891a-5p在結腸癌患者組織樣本以及結腸癌細胞系中的表達水平,并確定了miR-891a-5p的診斷和預后價值。此外,利用體外實驗研究了miR-891a-5p對結腸癌細胞增殖、遷移和侵襲及血管生成的影響,并通過分析潛在的靶基因初步確定了miR-891a-5p在結腸癌發展中的潛在機制。
1 資料與方法
1.1臨床樣本采集:患者從海南省腫瘤醫院2021年5月至2022年4月就診的36例患者中收集了結腸癌組織和鄰近正常組織(距離腫瘤邊緣至少3cm),所有收集的組織均儲存在-80℃以進行后續實驗。事先簽署患者知情同意書,并取得海南省腫瘤醫院研究所研究倫理委員會的批準。
1.2細胞培養和轉染:結腸癌細胞系(SW480,HCT8,HCT116和SW620)和正常人結腸上皮細胞(FHC)購自中國科學院細胞庫。所有細胞均在Dulbecco的改良Eagles培養基中孵育,并補充有10%胎牛血清(FBS)在37℃的含5% CO2的細胞培養箱中。使用Lipofectamine2000檢測試劑盒進行轉染細胞,僅用轉染試劑轉染的細胞用作空白對照組。細胞轉染在37℃下進行6h,然后更換培養基,并在轉染后48h將細胞用于后續分析。
1.3CCK-8:將細胞接種到96孔板(2×103細胞/孔)并孵育0h、24h、48h和72h。然后,向每個孔中加入10μL lCCK-8試劑,并進一步孵育2h。使用酶標儀在450nm處測量光密度。
1.4管形成測定:將50μL基質膠接種在96孔培養板中,然后在37℃下孵育。人臍靜脈內皮細胞(HUVECs)在具有不同外泌體或其他處理的基質膠上培養。在37℃孵育4h后,在顯微鏡下觀察管的形成,并通過測量累積管長度進行分析。
1.5RNA提取和逆轉錄定量PCR(RT-qPCR):使用PrimeScript逆轉錄酶試劑盒將RNA逆轉錄成單鏈cDNA。使用qPCR和SYBR-GreenI預混液試劑盒在7500實時熒光定量PCR系統。U6用作miR-891a-5p的內源性對照,GAPDH用作NLRP7的內源性對照。熱循環條件如下:初始變性在95℃下變性10min,然后在95℃下變性40個循環30s,在60℃退火20s,在72℃下延伸15s,最終表達式值是使用2-ΔΔCt方法。見表1。

表1 引物序列表
1.6傷口愈合試驗:細胞(1×105細胞/mL)在處理24h后接種在24孔板中,并孵育至70%-80%匯合。使用無菌移液器吸頭,通過刮擦細胞單層產生劃痕傷口。加入新鮮培養基,將細胞再孵育24h或不孵育,細胞在倒置顯微鏡下成像。
1.7Transwell:Transwell室在37℃下預包被基質膠1h用于侵襲測定,而沒有基質膠涂層的室用于遷移測定。轉染的細胞密度為2×105細胞/腔室用無血清培養基接種到上室中,同時將補充有10%FBS的培養基添加到下室中,作為化學引誘劑。在37℃孵育48h后,將下室中的細胞在室溫下用0.1%結晶紫染色20min,并在倒置光學顯微鏡下計數。
1.8靶基因預測和雙熒光素酶報告基因檢測:TargetScan在線工具(http://www.targetscan.org/vert_72)預測miR-891a-5p潛在的靶基因。為了確認miR-891a-5p和NLRP7在SW480細胞中的相互作用,將SW480細胞按照4×104細胞/孔接種到六孔板中,并在37℃下培養24h。隨后,將SW480細胞的野生型(WT)和突變型(MUT)3'-UTR克隆到熒光素酶報告載體PsiCheck2中。MUT序列是使用定點誘變試劑盒生成的。根據制造商的方案,將WT和MUT SW480細胞熒光素酶質粒(100ng)與miR-891a-5p mimic(50nM)和相應的NC共轉染到SW480細胞中。在37℃孵育48h后,使用雙熒光素酶報告檢測系統測量相對熒光素酶活性,螢火蟲熒光素酶活性被標準化為雷尼拉螢光素酶活性。
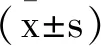
2 結 果
2.1miR-891a-5p在結腸癌中的表達水平:miR-891-5a在結腸癌組織樣本中的表達水平顯著高于健康對照組或鄰近正常對照組,如圖1A所示,還發現miR-891a-5p在結腸癌細胞系(SW480,HCT8,HCT116和SW620)中的表達水平顯著高于正常人結腸上皮細胞(FHC)(F=25.29,P<0.01),miR-891a-5p在SW480中表達最高,因此挑選SW480為后續研究,如圖1B所示,在測定miR-891a-5p在SW480細胞的細胞質和細胞核中的含量時,發現miR-891a-5p在細胞質中的表達高于細胞核中的表達量,如圖1C所示。

圖1 miR-891a-5p在結腸癌組織與細胞中的表達
2.2miR-891a-5p對SW480細胞增殖、侵襲及遷移作用:miR-891a-5p mimic和miR-891a-5p inhibitor轉染后,miR-891a-5p的mRNA表達水平分別在SW480細胞系中成功過表達和沉默(F=26.30,P<0.01),如圖2A所示,之后,使用CCK-8測定,轉染miR-891a-5p mimic的細胞增殖、侵襲及遷移能力顯著增加,但與對照組相比,轉染miR-891a-5p inhibitor的細胞增殖(F=28.93,P<0.01)、侵襲(F=21.24,P<0.01)及遷移(F=22.68,P<0.01)能力顯著降低。
2.3miR-891a-5p對SW480細胞血管生成的影響:圖3A結果表明,與Control比較miR-891a-5p mimic的血管生成數量明顯增加,而miR-891a-5p inhibitor的血管生成數量明顯降低(F=25.67,P<0.01),為驗證HUVEC管形成實驗的可靠性,使用Western blot對血管生成因子的相關蛋白VEGF和MMP-9進行檢驗,結果發現,VEGF和MMP-9在miR-891a-5p mimic組蛋白表達水平顯著增加,蛋白表達水平在miR-891a-5p inhibitor明顯降低(F=20.36,P<0.01)。

圖3 miR-891a-5p對SW480細胞血管生成的影響

圖4 NLRP7是miR-891a-5p下游靶點基因
2.4NLRP7是SW480細胞中miR-891a-5p的直接靶標:根據starBase v2.0網站(http://starbase.sysu.edu.cn/starbase2/)在線分析的結果,預測NLRP7是miR-891a-5p的靶標,在其3′-UTR,在NLRP7基因中發現了miR-891a-5p的結合位點,并利用熒光素酶報告基因檢測miR-891a-5p與NLRP7之間的相互作用。雙熒光素酶報告基因檢測結果表明,NLRP7-WT組中的熒光素酶活性因miR-891a-5p的表達水平上調而降低,然而,MUT組的熒光素酶活性沒有顯著變化(t=1.31,P=0.26)。NLRP7的表達水平在結腸癌患者腫瘤組織中明顯低于鄰近正常組織(t=10.08,P<0.001),與miR-891a-5p的表達水平呈負相關,上述結果表明,NLRP7可能是結腸癌中miR-891a-5p的直接靶標。
2.5NLRP7過表達對SW480細胞增殖、侵襲及遷移的作用:NLRP7作為miR-891a-5p的靶向目標,與OE-NC組相比,OE-NLRP7組中NLRP7表達水平顯著增加(t=18.12,P<0.01),表明細胞轉染成功,如圖5A所示,而且OE-NLRP7組的細胞增殖(t=20.17,P<0.01)、侵襲(t=23.15,P<0.01)及遷移(t=19.64,P<0.01)能力明顯下降。

圖5 NLRP7過表達對結腸癌增殖、侵襲及遷移的作用
2.6NLRP7過表達對SW480細胞血管生成的影響:Western blot實驗結果表明VEGF(t=13.88,P<0.01)和MMP-9(t=15.67,P<0.01)蛋白表達水平在OE-NLRP7顯著降低。圖6B結果表明,與OE-NC比較OE-NLRP7的血管生成數量明顯降低(t=20.17,P<0.01)。

圖6 NLRP7過表達對SW480細胞血管生成的影響
3 討 論
結腸癌發展的基礎機制為各種基因的轉錄和多種信號轉導相關的協同過程。盡管已有許多研究探索了結腸癌發展的分子機制,但還遠遠不夠。研究表明miRNA的異常表達已被確定與多種惡性腫瘤的發生密切相關,據報道,它們通過改變轉錄后基因表達參與細胞凋亡,增殖,細胞周期和轉移[8]。miRNA已被鑒定為腫瘤發生中的癌基因或抑癌基因,侵襲和遷移是腫瘤細胞的重要生物學行為,與預后不良密切相關,嚴重威脅患者的健康和生命。因此,深入分析遷移機制對提高結腸癌患者的生存率和預后具有重要意義。據報道,miR-891a-5p在許多癌癥中具有促進癌癥的功能[9],本研究使用36份結腸癌組織和鄰近正常組織檢測miR-891a-5p表達水平,我們的結果表明,結腸癌患者標本中的miR-891a-5p顯著增加。基質金屬蛋白酶-9(MMP-9)是侵襲和破壞細胞外基質所必需的,可促進癌細胞向附近健康組織的侵入性進展[10],在惡性腫瘤的侵襲性,遷移性和血管生成進展中具有不可或缺的地位。血管內皮生長因子(VEGF)是重要的促血管生成的生長因子,對內皮細胞具有有絲分裂和抗凋亡作用,增加血管通透性,促進細胞遷移等[11],遷移的開始不僅僅是細胞自主事件,而是受到復雜組織微環境的影響,腫瘤相關巨噬細胞可以被癌細胞分泌的乳酸刺激以促進血管生成,微環境對癌細胞侵襲和遷移的影響既是細胞自主的,也是組織環境依賴性的,這進一步增加了這一過程的復雜性。總而言之,miR-891a-5p的下調導致腫瘤細胞遷移、增殖和侵襲受到抑制,但miR-891a-5p的上調導致腫瘤細胞生物學行為增強。
本研究中,在NLRP7的3'UTR中發現了miR-891a-5p的互補序列,我們使用熒光素酶活性測定證明了miR-891a-5p與NLRP7的相互作用。此外,結腸癌患者miR-891a-5p與NLRP7 mRNA表達呈負相關。先前報道的關于NLRP7在結腸癌中的功能作用的研究,結合我們的數據啟發我們推斷miR-891a-5p可能通過靶向NLRP7參與結腸癌進展的調節。miR-891a-5p在結腸癌進展中的確切分子機制尚未深入探索,這是本研究的局限性,值得進一步研究。
NLR家族成員的重要作用是直接或間接促成癌癥發展,包括炎癥、細胞死亡、腫瘤生長、血管生成、侵襲和轉移[12]。在功能方面,一些成員可能是炎癥小體途徑的啟動者,其規范激活半胱天冬酶-1,此后激活IL-1β和IL-18[13],在NLR家族的所有成員中,NLRP7及其功能知之甚少。最近報道了NLRP7基因與潰瘍性結腸炎(UC)之間的相關性,并且發現NLRP7表達在炎癥性腸病患者的腸黏膜中上調[14]。考慮到炎癥性腸病患者發生結直腸癌的風險過高,且NLRP7在癌癥中的功能尚不清楚,我們結果證實,NLRP7可能在結腸癌進展中起到抑制作用,這與其他人的研究結果并不一致,因此其具體機制有待進一步探索。
但是本研究有幾個局限性,例如樣本量有限,缺乏動物實驗。此外,未研究甲基化等其他因素對NLRP7表達水平的影響,這可能干擾miR-891a-5p對NLRP7的調節。因此,在未來的研究中,可利用動物癌癥模型進一步研究miR-891a-5p在結腸癌發生中的功能作用和機制,以及miRNA 891a-5p/NLRP7軸的具體調控機制。
綜上所述,本研究結果表明,miR-891a-5p在結腸癌組織的表達增加,可作為結腸癌診斷和預后生物標志物。miR-891a-5p過表達可促進結腸癌細胞增殖、遷移、侵襲及血管生成能力,表明miR-891a-5p可能是結腸癌進展中的癌基因,對結腸癌的治療具有靶向潛力。NLRP7也被確定為miR-891a-5p的靶基因,可能介導miR-891a-5p在結腸癌進展中的生物學功能。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
西南國防醫藥(2016年7期)2016-12-01 06:01:15
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
