長(zhǎng)枝木霉聚合根腫菌多元RNAi 靶標(biāo)防治榨菜根腫病的研究
2023-09-20 08:29:34蔡兆明王殿東
湖南農(nóng)業(yè)科學(xué) 2023年8期
劉 穎,蔡兆明,王殿東
(長(zhǎng)江師范學(xué)院現(xiàn)代農(nóng)業(yè)與生物工程學(xué)院,重慶 涪陵區(qū) 408100)
榨菜(Brassica juncea)又名莖瘤芥,是一種重要的十字花科蔬菜作物,廣泛栽培于中國(guó)南部地區(qū),具有較高的營(yíng)養(yǎng)價(jià)值和經(jīng)濟(jì)價(jià)值[1]。根腫病是榨菜栽培中的主要病害,對(duì)其產(chǎn)量和品質(zhì)造成極大影響[2]。根腫病是由蕓薹根腫病菌(Plasmodiophora brassicae)引起的一種土傳性病害,主要危害十字花科作物,植株染病后根部首先出現(xiàn)病斑,隨后根部腫大和腐爛,嚴(yán)重時(shí)會(huì)造成組織腐爛,植株死亡[3-6]。據(jù)統(tǒng)計(jì),全球已有60 多個(gè)國(guó)家受到根腫病的影響,一旦染病通常會(huì)造成10%~15%的產(chǎn)量損失[7-9],尤其是對(duì)榨菜產(chǎn)量的影響較大,嚴(yán)重時(shí)可造成36%以上的損失[8]。目前,根腫病的防治方法主要包括農(nóng)業(yè)防治[10]、化學(xué)防治[11]和生物防治[12]。農(nóng)業(yè)防治只能在一定程度上預(yù)防或緩解根腫病的癥狀,但是無(wú)法達(dá)到根治的目的;化學(xué)防治是現(xiàn)在運(yùn)用最為普遍、效果也最好的防治方法,但是長(zhǎng)期施藥會(huì)給農(nóng)產(chǎn)品安全以及生態(tài)環(huán)境帶來(lái)一定風(fēng)險(xiǎn);生物防治對(duì)農(nóng)產(chǎn)品和環(huán)境均較安全,是現(xiàn)階段防治根腫病的研究熱點(diǎn)[13]。
MAPKs 是一類絲裂原活化蛋白激酶,MAPK基因在細(xì)胞的生長(zhǎng)和繁殖中十分重要,根腫菌中的MAPK基因在根腫菌對(duì)植物進(jìn)行表層侵染的時(shí)候起作用[14-16]。小干擾RNA(Small interfering RNA,siRNA)又稱短干擾RNA(short interfering RNA)或沉默RNA(silencing RNA),是一個(gè)長(zhǎng)20 到25個(gè)核苷酸的雙股RNA,在生物學(xué)上有許多不同的用途。目前,已知siRNA 主要參與RNA 干擾(RNAi)現(xiàn)象,以帶有專一性的方式調(diào)節(jié)基因的表達(dá),是一種常用的遺傳學(xué)研究手段[17]。通過(guò)siRNA 技術(shù)對(duì)MAPK基因進(jìn)行靶向沉默,從而干擾MAPK 信號(hào)通路,在植物抗病研究中取得了顯著的成果[18]。王鵬程[19]通過(guò)沉默擬南芥(Arabidopsis thaliana)中MAPK2基因發(fā)現(xiàn),突變體植株生長(zhǎng)更加旺盛,并表現(xiàn)出與野生型不同的形態(tài)特征,例如蓮座葉增多、或分層,花芽萌發(fā)時(shí)缺乏頂端優(yōu)勢(shì),次級(jí)花序生長(zhǎng)增強(qiáng)等。這些差異可能暗示了MAPK2 在發(fā)育過(guò)程中的作用,同時(shí)也預(yù)示著這種差異與生長(zhǎng)素的信號(hào)轉(zhuǎn)導(dǎo)途徑的關(guān)系。然而,關(guān)于MAPK基因在根腫病發(fā)病過(guò)程中的作用報(bào)道較少。該研究旨在通過(guò)RNAi 技術(shù)對(duì)生防菌長(zhǎng)枝木霉的MAPK1、MAPK2、MAPK3基因進(jìn)行沉默,以研究MAPK基因在榨菜根腫病防治過(guò)程中的作用。
1 材料與方法
1.1 試驗(yàn)材料
供試生防菌為長(zhǎng)枝木霉,供試蕓薹根腫菌采自重慶涪陵榨菜種植地,保存于長(zhǎng)江師范學(xué)院榨菜病蟲害防治實(shí)驗(yàn)室,供試種子為涪雜2 號(hào)榨菜種子。
1.2 構(gòu)建表達(dá)載體
從發(fā)病榨菜腫根中提取根腫菌孢子,提取總RNA 反轉(zhuǎn)錄為cDNA,以cDNA 為模板分別以MAPK1-360F/MAPK1-673R、MAPK2-776F/MAPK2-1126、MAPK3-560F/MAPK3- 885R 為引物擴(kuò)增根腫菌MAPK1、MAPK2、MAPK3基因的cDNA 片段后,以MAPK1、MAPK2基因片段為模板,以MAPK1-360F、MAPK1-JR、MAPK2-JR、MAPK2-1126R 為引物擴(kuò)增MAPK1和MAPK2基因融合片段,以MAPK1-360F、MAPK2-JR 、MAPK3-JF、MAPK3-885R 為引物,以MAPK1和MAPK2基因融合片段和MAPK3基因片段為模板擴(kuò)增MAPK1、MAPK2、MAPK3基因融合片段;以融合片段為模板,分別以引物MAPK1-Hind3-360F/MAPK3-Xba1- 885R 和MAPK1-Bgl2-360F/MAPK3-Spe1-885R 擴(kuò)增含有酶切位點(diǎn)的正向序列和反向序列,在HindⅢ、XbaI酶切位點(diǎn)處加入正向片段,在BglⅡ、SpeI酶切位點(diǎn)處加入反向片段到載體pDH-RH 上,構(gòu)建pDH-RHMAPK 載體(圖1)。所用引物序列如表1 所示,由生工生物工程(上海)股份有限公司合成。

表1 載體構(gòu)建所用引物序列
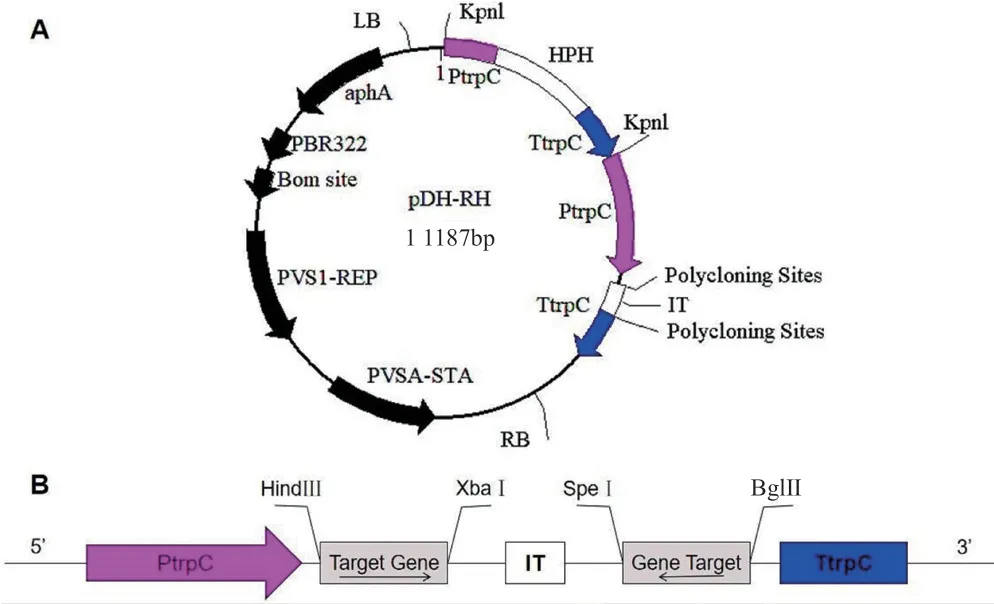
圖1 絲狀真菌pDH-RH-MAPK 載體示意圖
1.3 pDH-RH-MAPK 載體轉(zhuǎn)化長(zhǎng)枝木霉
供試生防菌長(zhǎng)枝木霉首先采用液體培養(yǎng)獲得菌絲。收集菌絲用0.7 mol/L 的氯化鈉溶液沖洗至團(tuán)狀,按1 g 菌絲加入20 mg 崩潰酶(20 mg/mL)的量向菌絲中加入崩潰酶,在28℃、150 r/min 的條件下酶解6 h,然后用0.7 mol/L 氯化鈉溶液反復(fù)沖洗后收集原生質(zhì)體,再用5 mL STC 溶液(1.2 mol/L 山梨醇,50 mmol/L 氯化鈣,10 mmol/L Tris-HCl)重懸。采用PEG 介導(dǎo)的原生質(zhì)體轉(zhuǎn)化方法進(jìn)行轉(zhuǎn)化,用含有60 mg/L 潮霉素B 的PDA 培養(yǎng)基篩選陽(yáng)性轉(zhuǎn)化子。
1.4 轉(zhuǎn)基因菌株鑒定
經(jīng)潮霉素B 篩選后,提取陽(yáng)性轉(zhuǎn)化子(即轉(zhuǎn)基因的長(zhǎng)枝木霉菌)的基因組DNA,以未轉(zhuǎn)化的長(zhǎng)枝木霉菌基因組DNA 和ddH2O 為對(duì)照,以潮霉素引物(Hyg-F:5'- AAGCCTGAACTCACCGCGAC-3';Hyg-R:5'- CTATTCCTTTGCCCTCGGAC-3')進(jìn)行PCR 擴(kuò)增,檢測(cè)是否轉(zhuǎn)化成功。
1.5 轉(zhuǎn)基因長(zhǎng)枝木霉菌對(duì)根腫病防效分析
將轉(zhuǎn)基因的長(zhǎng)枝木霉菌配置成濃度為1×107個(gè)/mL的懸浮液,取30 mL 轉(zhuǎn)基因長(zhǎng)枝木霉菌懸液浸根處理榨菜幼苗30 min,然后將幼苗移栽至含有根腫菌的土壤(含8×107個(gè)根腫菌)中,以無(wú)菌水和未轉(zhuǎn)基因長(zhǎng)枝木霉菌為對(duì)照,接種后40 d 取樣,測(cè)量根鮮重,并統(tǒng)計(jì)發(fā)病株數(shù)及發(fā)病情況。根腫病病害分級(jí)標(biāo)準(zhǔn)如下:0 級(jí),根部無(wú)腫瘤;1 級(jí),僅側(cè)根有小腫瘤;2 級(jí),側(cè)根有大腫瘤,主根有小腫瘤;3 級(jí),主根有大腫瘤。每個(gè)處理調(diào)查20株,計(jì)算發(fā)病率和病情指數(shù)。
發(fā)病率(%)=(發(fā)病株數(shù)/總株數(shù))×100
病情指數(shù)=[(各級(jí)病株數(shù)×相對(duì)病級(jí)數(shù)值)/(調(diào)查總株數(shù)×最高病級(jí)數(shù)值)]×100
1.6 轉(zhuǎn)基因長(zhǎng)枝木霉菌處理后根腫菌中靶標(biāo)基因表達(dá)分析
收集轉(zhuǎn)基因長(zhǎng)枝木霉菌、未轉(zhuǎn)基因長(zhǎng)枝木霉菌和無(wú)菌水處理40 d 后的根腫菌(來(lái)自于發(fā)病的榨菜腫根),提取總RNA,反轉(zhuǎn)錄為cDNA,以根腫菌Actin2基因?yàn)閮?nèi)參進(jìn)行熒光定量PCR(qRT-PCR),檢測(cè)根腫菌中MAPK1、MAPK2、MAPK3基因的表達(dá)量,qRT-PCR 的引物序列見表2,由生工生物工程(上海)股份有限公司合成。

表2 qRT-PCR 的引物序列
2 結(jié)果與分析
2.1 融合MAPK1、MAPK2、MAPK3 基因的dsRNA 表達(dá)載體構(gòu)建
PCR 擴(kuò)增后分別得到根腫菌MAPK1、MAPK2、MAPK3基因的cDNA 片段,大小分別為314、351、326 bp(圖2A),與預(yù)測(cè)大小一致,測(cè)序后基因組序列也一致,可用于后續(xù)試驗(yàn)。隨后按照1.2 中的步驟依次將3 個(gè)基因融合,其中,MAPK1和MAPK2基因的融合片段大小為660 bp(圖2B),與預(yù)測(cè)大小一致;3 個(gè)基因的融合片段大小為1 000 bp 左右(圖2C),符合連接片段對(duì)應(yīng)大小。
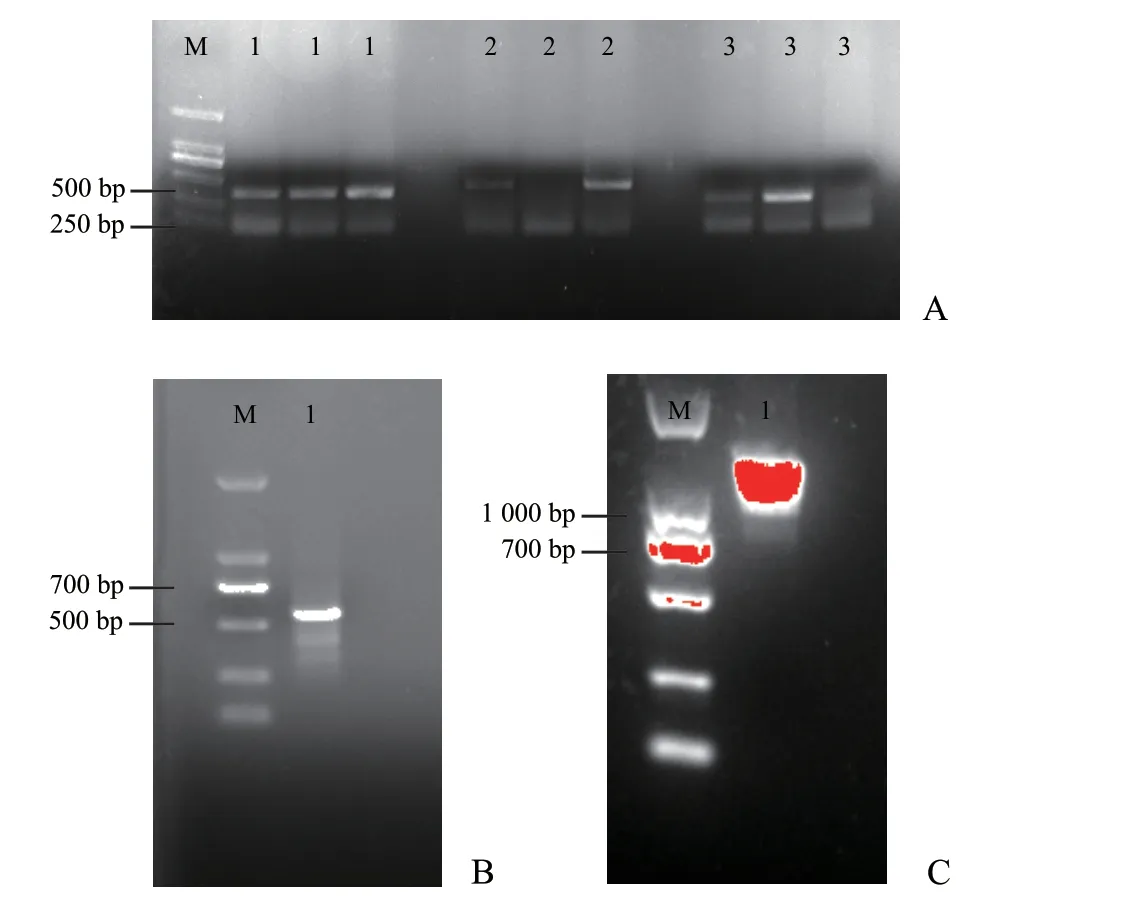
圖2 3 個(gè)MAPK 基因克隆及融合片段的電泳檢測(cè)結(jié)果
將根腫菌MAPK1、MAPK2、MAPK3基因融合片段分別在HindⅢ、XbaI和BglⅡ、SpeI酶切位點(diǎn)處正向和反向連接到絲狀真菌dsRNA 表達(dá)載體上,構(gòu)建木霉菌表達(dá)MAPK1、MAPK2、MAPK3基因融合片段的 dsRNA 載體,結(jié)果如圖3 所示。
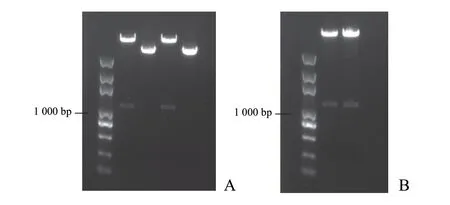
圖3 pDH-RH-MAPK 載體正向連接和反向連接的酶切電泳檢測(cè)結(jié)果
2.2 pDH-RH-MAPK 載體轉(zhuǎn)化長(zhǎng)枝木霉的結(jié)果
利用原生質(zhì)體轉(zhuǎn)化法,將表達(dá)根腫菌3 個(gè)MAPK基因的dsRNA 載體轉(zhuǎn)化到長(zhǎng)枝木霉菌株,挑取陽(yáng)性轉(zhuǎn)化子,利用潮霉素抗性基因引物進(jìn)行菌落PCR 鑒定,獲得陽(yáng)性轉(zhuǎn)化子(圖4)。利用根腫菌中MAPK1、MAPK2、MAPK3基因特異引物檢測(cè)3 個(gè)基因片段在轉(zhuǎn)基因長(zhǎng)枝木霉中的表達(dá)情況,發(fā)現(xiàn)3個(gè)基因的RNA 均在長(zhǎng)枝木霉中進(jìn)行表達(dá)(圖5)。
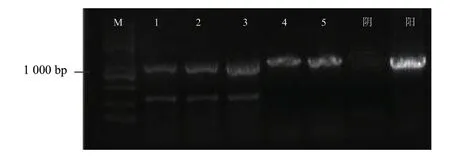
圖4 潮霉素抗性引物 PCR 檢測(cè)結(jié)果
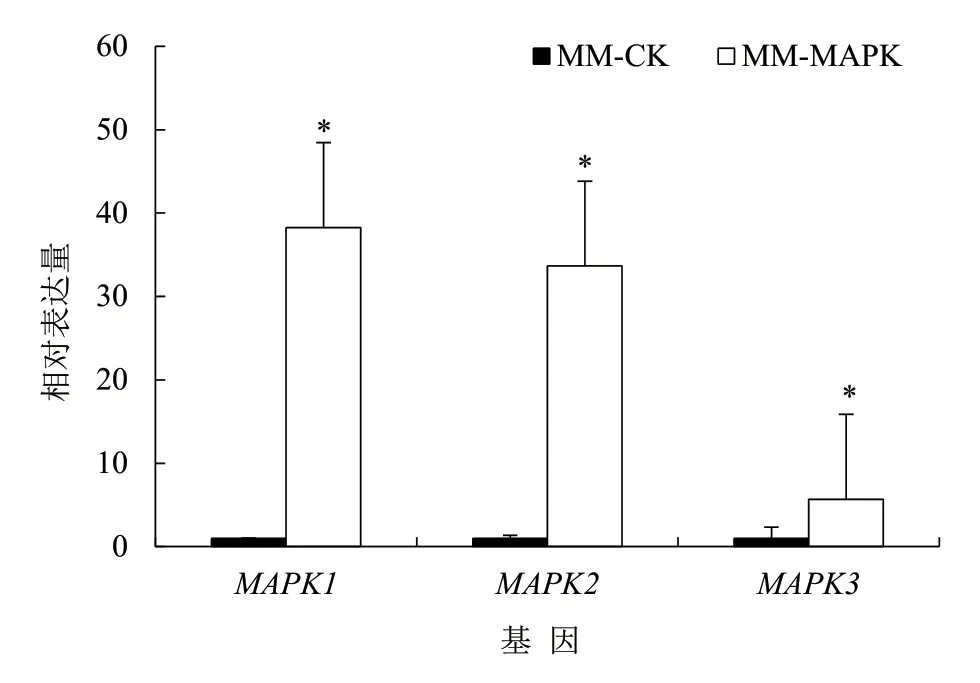
圖5 熒光定量PCR 檢測(cè)3 個(gè)MAPK 基因在轉(zhuǎn)基因長(zhǎng)枝木霉中的表達(dá)情況
2.3 轉(zhuǎn)基因長(zhǎng)枝木霉對(duì)根腫菌MAPK 基因表達(dá)水平的影響
采集接種轉(zhuǎn)基因長(zhǎng)枝木霉菌和對(duì)照處理的榨菜根腫病樣本,分離根腫菌并提取總RNA,反轉(zhuǎn)錄為cDNA,通過(guò)熒光定量PCR 檢測(cè)靶標(biāo)基因MAPK1、MAPK2、MAPK3基因表達(dá)量,結(jié)果如圖6 所示,轉(zhuǎn)基因長(zhǎng)枝木霉菌處理榨菜后可顯著降低根腫菌中MAPK1、MAPK2、MAPK3基因的表達(dá)量。
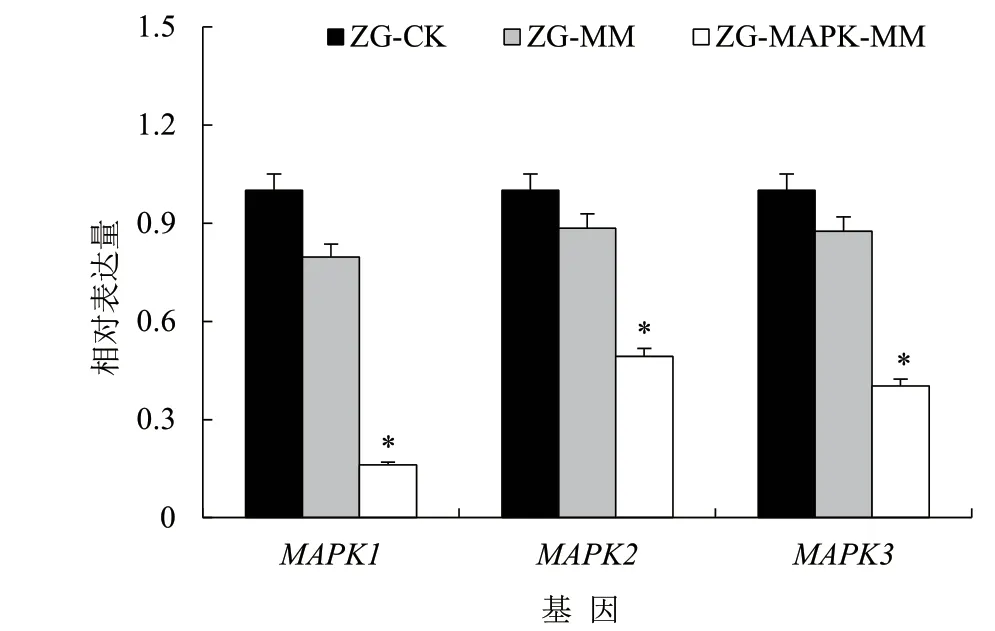
圖6 各處理根腫菌MAPK 基因的表達(dá)情況
2.4 接種轉(zhuǎn)基因長(zhǎng)枝木霉后對(duì)榨菜根腫病的防效
由圖7 可知,各處理榨菜根腫病發(fā)病率為100%,其中空白對(duì)照組(CK)的平均鮮重為 1.37 g,病情指數(shù)為 62;接種未轉(zhuǎn)基因長(zhǎng)枝木霉處理(MM-CK)的平均鮮重為 1.30 g,病情指數(shù)為 63;接種轉(zhuǎn)基因長(zhǎng)枝木霉處理(MM-MAPK)的平均鮮重為 1.17 g,病情指數(shù)為 55,MM-MAPK 處理的榨菜根鮮重和病情指數(shù)均顯著低于CK 和MM-CK 處理,而MM-CK處理的榨菜根鮮重顯著低于CK,但病情指數(shù)與CK無(wú)顯著差異。
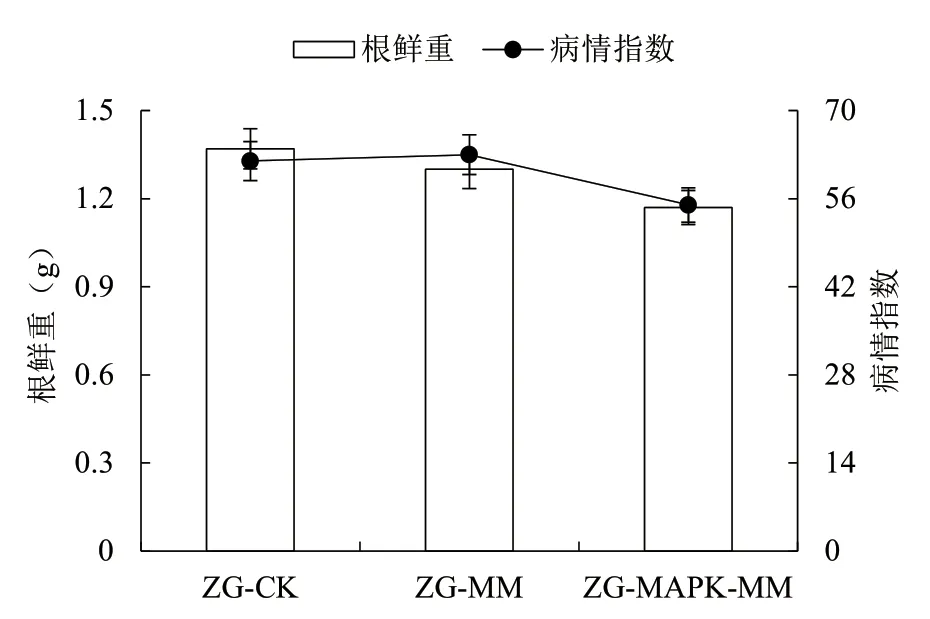
圖7 各處理榨菜發(fā)病腫根的平均根鮮重和病情指數(shù)
3 結(jié)論與討論
根腫病是十字花科蔬菜栽培中危害最為嚴(yán)重的病害,榨菜不同于其他十字花科蔬菜,其本身是野生四倍體,這給抗病育種帶來(lái)了極大的障礙。該研究所采用的長(zhǎng)枝木霉分離自榨菜根腫病發(fā)病田,是一株對(duì)根腫病具有生防作用的菌株。通過(guò)RNAi 技術(shù)對(duì)該長(zhǎng)枝木霉菌株中多個(gè)MAPK基因進(jìn)行聯(lián)合干擾,可增強(qiáng)長(zhǎng)枝木霉對(duì)根腫病病原菌的抗性,顯著抑制根腫菌中3 個(gè)MAPK基因的表達(dá),從而提高長(zhǎng)枝木霉對(duì)榨菜根腫病的防效,降低榨菜根腫病的病情指數(shù)。MAPK 蛋白通路是病原菌和植物發(fā)育和致病過(guò)程中極為重要的信號(hào)通路,參與了多種生物學(xué)過(guò)程,包括細(xì)胞分裂、生長(zhǎng)發(fā)育、應(yīng)對(duì)生物和非生物脅迫等,在植物對(duì)病原體侵染和逆境脅迫的響應(yīng)中發(fā)揮了重要作用[20-22]。該研究中采取的長(zhǎng)枝木霉聚合siRNA 策略可以同時(shí)靶向多個(gè)MAPK基因,從而達(dá)到更好的防治效果。同時(shí),采用生防菌防治病害的這種手段專一性強(qiáng)、效率高、整體危害較輕,可有效減輕根腫菌對(duì)榨菜產(chǎn)業(yè)的危害。
