苦蕎對發酵豆乳納豆激酶活力、風味及抗氧化活性的影響
2023-08-22 08:04:38王淼霜仝艷軍蔣雨橋楊瑞金
食品與生物技術學報 2023年7期
王淼霜, 仝艷軍, 蔣雨橋, 楊瑞金*1,
(1. 江南大學 食品科學與資源挖掘全國重點實驗室, 江蘇 無錫 214122;2. 江南大學 食品學院, 江蘇 無錫 214122)
心腦血管疾病(cardiovascular disease,CVD)是造成我國居民死亡的首要病因之一[1]。 從病理上來看, 形成CVD 的主要原因是纖維蛋白在循環系統的累積。 外源性纖維蛋白溶解酶可有效溶解纖維蛋白并改善血流情況,如尿激酶、鏈激酶、納豆激酶(nattokinase,NK)等[2]。 其中,NK 是近幾十年來的研究熱點,與其他纖維蛋白溶解酶相比,NK 不僅具有成本低、療效好、有效時間長、安全性高的特點,還具有多種CVD 的預防和緩解作用,如抗血栓、抗高血壓、抗動脈粥樣硬化、降脂和神經保護等[3]。
目前市場上納豆激酶的主要來源是納豆、納豆激酶膠囊、納豆激酶片劑,但這些產品存在成本高、有異味、吞咽困難等缺點。 關于納豆的制作分為固態發酵和液態發酵兩大類。 相比固態發酵,液態發酵更適用于納豆激酶的大量工業化生產及提取純化[4-5]。 發酵豆乳是在液態豆乳中接種納豆芽孢桿菌,經發酵后獲得的營養豐富的發酵豆制品, 但發酵豆乳產品刺鼻的“氨臭味”常常使消費者望而卻步。
有研究表明“氨臭味”的主要來源為揮發性鹽基氮和含硫化合物[6],納豆產品的揮發性鹽基氮未有國家標準規定,但存在相關文獻報道。 鄭丹妮利用多菌種復合發酵低“氨臭味”的納豆,最終產品中揮發性鹽基氮質量分數為198.66 mg/hg, 酶活力為341.25 U/g[7]。 為了改善納豆的“氨臭味”,許多研究人員從發酵底物入手進行創新。 陳樂樂等在以高蛋白質大豆為原料制備納豆過程中加入超甜玉米,結果發現隨著超甜玉米質量分數的提高,納豆激酶活力呈現先上升后下降的趨勢,揮發性鹽基氮呈現持續降低的趨勢;高蛋白質大豆和超甜玉米質量比為8∶2 時,納豆的揮發性鹽基氮質量分數為32.87 mg/g,納豆激酶活力達到1 134.76 U/g[8]。 張杰等通過糯米與小黃豆復合發酵獲得品質較好的納豆,結果表明“氨臭味” 顯著降低, 揮發性鹽基氮質量分數為15.41 mg/hg,納豆激酶活力為382.32 U/g[9]。 趙謀明等在納豆菌液態基礎培養基中添加不同谷物(糙薏仁、玉米、蕎麥、糙米)并采用不同的谷物處理方式探究對納豆激酶活力的影響,結果表明采用浸泡蒸煮的方式處理谷物并添加糙米可顯著促進納豆菌產納豆激酶[10]。此外,有文獻報道以糙米作為發酵底物,采用納豆芽孢桿菌發酵,總必需氨基酸和脂肪質量分數增加, 粗蛋白質質量分數減少,2,2-二苯基-1-苦基肼(DPPH)自由基清除能力顯著增加[11]。由上述報道可見谷物對納豆的納豆激酶活力、揮發性鹽基氮質量分數等方面存在積極影響,在納豆制作方面具有應用價值。
谷物通常具有良好的生物活性[12-16],比如:苦蕎具有降血脂、抗氧化、增強人體免疫力的作用;燕麥經枯草芽孢桿菌發酵后可提升其蛋白質含量。 作者以實驗室前期研究中篩選出的一株高產納豆激酶且產“氨臭味”少的納豆芽孢桿菌JNFE0127 作為發酵菌株。 通過添加不同谷物(苦蕎、糙米、薏米、藜麥、燕麥粉)探究納豆芽孢桿菌JNFE 0127 產納豆激酶的能力,確定使其產酶量高的谷物種類;然后研究該谷物不同添加量對納豆芽孢桿菌JNFE 0127發酵豆乳的納豆激酶活力、風味及抗氧化活性的影響。 該研究提高了液態發酵豆乳制品的酶活力并改善其風味,為產業發展提供理論依據。
1 材料與方法
1.1 材料與試劑
納豆芽孢桿菌JNFE 0127 (Bacillus natto JNFE 0127,簡稱為JNFE 0127):作者所在實驗室前期篩選并保存;大豆、苦蕎、糙米、薏米、藜麥、燕麥:購自京東商城;牛凝血酶(1 000 U/g)、牛纖維蛋白原:沈陽拜英生物技術有限公司產品;2,4,6-三吡啶基三嗪(純度98%)、2,2-二苯基-1-苦基肼(純度98%)、5,6-二苯基-3-(2-吡啶基)-1,2,4-三嗪-4,4’-二磺酸單鈉鹽(純度97%):北京沃凱生物科技有限公司產品;其余試劑均購自國藥集團化學試劑有限公司。
1.2 儀器與設備
LE2002E 型分析天平、FE28 型pH 計:梅特勒-托利多儀器(上海)有限公司產品;LDZX-50KBS 型立式壓力蒸汽滅菌器: 上海申安醫療器械廠產品;DRP-9082 型電熱恒溫培養箱: 上海森信實驗儀器有限公司產品;MQD-S3R 型振蕩培養箱:上海旻泉儀器有限公司產品;HH-4 型數顯恒溫水浴鍋:常州國華電器有限公司產品;UV-1800PC 型紫外可見分光光度計: 上海美譜達儀器有限公司產品;Super Mini Dancer 桌面型迷你離心機:生工生物工程(上海)股份有限公司產品;高速冷凍離心機:賽默飛世爾科技(上海)有限公司產品;KQ52000DE 型數控超聲波清洗機:昆山市超聲儀器有限公司產品;破壁機: 九陽股份有限公司產品;4500A 型多功能粉碎機:永昌市展帆工貿有限公司產品;快速氣相色譜電子鼻:法國Alpha MOS S.A.有限公司產品。
1.3 培養基的配制
LB 固體培養基:胰蛋白胨質量分數1.0%、酵母提取物質量分數0.5%、氯化鈉質量分數1.0%、瓊脂質量分數2.0%。
活化培養基:無水葡萄糖質量分數0.5%、大豆蛋白胨質量分數1.0%、氯化鈉質量分數0.5%。
基礎發酵培養基:酵母粉質量分數0.50%、葡萄糖質量分數1.50%、CaCl2質量分數0.02%、NaCl 質量分數0.20%、MgSO4質量分數0.05%,0.1 mol/L K2HPO4/KH2PO4緩沖液調pH 至7.2。
發酵豆乳培養基:大豆粉質量分數11.1%、水質量分數89.9%。
1.4 方法
1.4.1 種子發酵液的制備 將保藏于-80 ℃甘油管中的JNFE 0127 菌液劃線接種到LB 固體培養基上,37 ℃恒溫培養10 h; 分別挑取JNFE 0127 單菌落接種于活化培養基中,37 ℃、200 r/min 培養12 h,即為種子發酵液。
1.4.2 樣品的制備
1)基礎發酵液的制備 取活化的JNFE 0127種子發酵液按照體積分數2%接種于基礎發酵培養基中,37 ℃、200 r/min 分別培養24、36、48、60、72、96 h。
2)不同谷物發酵乳的制備 在基礎發酵培養基中分別添加苦蕎粉、薏米粉、糙米粉、藜麥粉、燕麥粉,添加質量分數分別為1%、3%、6%,裝液量50 mL,115 ℃滅菌30 min,冷卻后按照體積分數2%接入活化的種子發酵液, 于39 ℃、200 r/min 培養24、36、48、60、72、96 h。
3)不同質量比的發酵豆乳制備 將大豆清洗并浸泡12 h,瀝干后加入濕豆質量8 倍的水用破壁機打漿5 min, 再添加體積分數9%的42 型果葡糖漿, 使用高速剪切分散機以10 000 r/min 剪切15 min。采用相同方法處理谷物。將不同質量比的大豆漿和谷物漿(3∶1、2∶1、1∶1、1∶2、1∶3)以及純豆乳和純谷物漿裝入250 mL 三角瓶中, 裝液量100 mL,105 ℃滅菌25 min,冷卻后接入體積分數2%的種子發酵液,39 ℃、200 r/min 搖瓶發酵12 h。
1.4.3 NK 活力的測定 參考Astrup 等的方法測定NK 活力[17]。 將滅菌生理鹽水置于37 ℃水浴鍋中保溫, 取0.15 g 纖維蛋白原溶于15 mL 生理鹽水,37 ℃水浴溶解10 min, 制成纖維蛋白原溶液。 將100 μL 凝血酶(10 U/mL)溶于400 μL 生理鹽水中,制成2 U/mL 的凝血酶溶液。 將1.25 g 瓊脂粉加入125 mL 去離子水中,加熱至完全溶解。 待瓊脂溶液溫度降低至55 ℃左右時, 與配好的纖維蛋白原溶液和凝血酶溶液快速混勻并立即倒平板, 每個板25~27 mL。 待完全凝固后打孔。 在纖維蛋白原平板孔內加入10 μL 發酵液,37 ℃靜置培養18 h, 用游標卡尺測量透明圈直徑。1 min 轉化1 μmoL 底物所需酶量為1 個酶活力單位(U)。
1.4.4 pH 及酸度的測定
1)pH 的測定 使用pH 計測定發酵液pH。
2)酸度的測定 配置0.1 mol/L 氫氧化鈉溶液并用鄰苯二甲酸氫鉀(KHP)進行標定,取10 mL 發酵液加20 mL 滅菌水,混勻,用氫氧化鈉標準溶液滴定直至終點(pH 8.3)[18]。 記錄消耗氫氧化鈉標準溶液的體積V1,按公式(1) 計算:
式中:X 為樣品的酸度,°T;c 為氫氧化鈉標準溶液的濃度,mol/L;V1為滴定所消耗氫氧化鈉標準溶液的體積,mL;V2為空白組消耗氫氧化鈉標準溶液的體積,mL;m 為樣品的質量,g。
1.4.5 揮發性鹽基氮的測定 參考GB 5009.228—2016《食品中揮發性鹽基氮的測定》[19]。 取10 mL 樣品于消化管中,加入75 mL 水,振蕩均勻,浸漬30 min。待浸漬結束后,加入1 g 氧化鎂,立刻連接到凱氏定氮儀上,設置加堿體積和加水體積均為0,蒸餾時間180 s。 將1 份甲基紅乙醇溶液與5 份溴甲酚綠乙醇溶液混合作為指示劑,量取30 mL 硼酸接收液,滴加10 滴混合指示劑,以指示劑顏色的變化來判斷終點。 揮發性鹽基氮質量濃度的計算見公式(2):
式中:ρ 為樣品揮發性鹽基氮質量濃度,mg/dL;V1為消耗鹽酸的體積,mL;V2為空白組消耗鹽酸的體積,mL;c1為鹽酸滴定液的濃度,mol/L;V 為樣品的體積,mL。
1.4.6 抗氧化活性的測定
1)總抗氧化能力的測定 采用文獻[20]的方法, 將0.3 mol/L 醋酸緩沖液 (pH 3.6)、10 mmol/L TPTZ 溶 液、20 mmol/L FeCl3溶液按體 積比10∶1∶1混合得到FRAP 工作液。 取發酵液1 mL,5 000 g 離心10 min,取離心后的上清液100 μL,加入2.4 mL FRAP 工作液, 空白組取去離子水100 μL, 加入2.4 mL FRAP 工作液, 分別混勻后37 ℃水浴10 min,在593 nm 下用分光光度計測吸光度。 依照FeSO4·7H2O 標準曲線計算總抗氧化能力。
2)DPPH 自由基清除能力的測定 參考Ai 等的方法[21]稍加改動。 在1 mL 發酵液中加入4 mL 去離子水混勻,5 000 g 離心10 min, 取上清液500 μL,加入500 μL 0.2 mmol/L DPPH 溶液(用體積分數95%乙醇定容),空白組取100 μL 體積分數95%乙醇,加入500 μL 0.2 mmol/L DPPH 溶液,分別混勻后25 ℃避光水浴30 min,設定檢測波長為517 nm。DPPH 自由基清除能力按公式(3)計算:
式中:Y 為樣品的DPPH 自由基清除能力,%;As為樣品組的吸光度;Ac為空白組的吸光度。
3)亞鐵離子螯合能力的測定 參照文獻[22]的方法測定亞鐵離子螯合能力。 量取10 mL 發酵液,5 000 g 離心10 min, 準確取3 mL 上清液加入50 μL 2 mmol/L FeCl3溶液、200 μL 5 mmol/L 菲洛嗪溶液和750 μL 去離子水,空白組取3.75 mL 去離子水 加 入50 μL 2 mmol/L FeCl3溶 液 和200 μL 5 mmol/L 菲洛嗪溶液,分別混勻后25 ℃孵育30 min,在562 nm 波長下測定吸光度。 亞鐵離子螯合能力按公式(4)計算:
式中:Z 為樣品的亞鐵離子螯合能力,%;A1為樣品組的吸光度;A0為空白組的吸光度。
1.4.7 電子鼻測定 取10 mL 發酵液于20 mL 氣相樣品瓶中,旋緊瓶蓋,按大豆含量遞減的順序擺放,重復兩次,首尾各放一個空瓶作為空白樣,采用HeraclesII 系統進行自動測定,分析軟件為該儀器配套軟件。
1.4.8 數據處理 利用SPSS 20.0 進行方差分析及ANOVA 差異顯著性分析(P<0.05 為差異顯著)。 利用Origin 9.0 軟件處理數據并繪圖。
2 結果與討論
2.1 谷物對JNFE 0127 發酵產NK 能力及pH 的影響
2.1.1 谷物對JNFE 0127 發酵產NK 能力的影響近年來液態發酵制備NK 產品已經成為趨勢。 選擇特色的發酵底物,并對發酵條件進行優化可以實現底物的高效利用。 趙謀明等以糙米為底物,通過優化納豆芽孢桿菌液態發酵條件, 制備具有高NK 活力和富含谷物多酚的發酵產物[23]。 作者選擇消費者普遍認可的苦蕎、燕麥、藜麥等5 種富含膳食纖維、蛋白質、多酚、礦物質等成分[12-16]的健康谷物作為添加物展開研究。 如圖1 所示,將JNFE 0127 在不添加谷物的基礎發酵培養基中發酵,48 h 時產酶量最高,可達到6 000 U/mL 以上。 在基礎發酵培養基中添加不同谷物后,產酶量顯著提高。 值得注意的是,在發酵時間和谷物添加質量分數一致的條件下,不同谷物對JNFE 0127 發酵產NK 能力的影響不同。用同一種谷物進行發酵時,添加質量分數影響產酶量的大小, 添加質量分數為3%時, 產酶量普遍更高。 添加質量分數3%的苦蕎粉發酵96 h 時,其產酶量為23 326.23 U/mL,與基礎發酵培養基相比提高301.96%。 目前文獻報道的納豆芽孢桿菌產酶量為300~15 000 U/mL[24-27]。 比如,王艷平等利用納豆芽孢桿菌固態發酵花生粕制備NK, 經正交優化后產酶量為3 162 U/mL[28]。 根據以上信息可以說明添加苦蕎粉可以明顯提升產酶能力。 此外,當谷物添加質量分數一定時,隨著發酵時間的延長,產酶量顯示出達到峰值后回落的變化規律。 因此發酵時間是產酶能力的重要影響因素。 在發酵過程中,應及時終止發酵預防酶回落現象,劉文濤等也發現了該現象[29]。

圖1 谷物對JNFE 0127 發酵產NK 的影響Fig. 1 Effect of grains on NK production by fermentation of JNFE 0127
2.1.2 谷物對JNFE 0127 發酵過程pH 的影響pH是評價微生物生長過程的一項重要指標。 依據pH可以了解菌株的生長周期、對培養基的適應能力以及NK 的穩定性等情況。 由圖2 可知, 在發酵過程中, 發酵體系的pH 呈現先減小后增加的趨勢,在24~60 h 發酵階段,pH 基本逐漸降低, 在發酵60 h時pH 為5.6 左右; 發酵60~96 h, 體系pH 逐漸升高, 在96 h 發酵結束后, 體系pH 基本均在6.0 以上。 當JNFE 0127 處于對數生長期時,快速消耗培養基中的碳源和氮源等營養物質,同時會分泌酸性代謝產物,如奎尼酸、琥珀酸等[30],使pH 逐漸降低;在進入穩定期后,培養基中可供直接利用的碳源被逐步消耗完畢,JNFE 0127 會將有機酸作為碳源,氮源中的碳元素也會被利用,并以氨的形式釋放氮元素,導致培養基pH 升高;進入衰亡期后,由于菌體自溶的原因,培養基的pH 會進一步升高。此處培養基pH 的變化規律與上述JNFE 0127 的生長周期一致。

圖2 谷物對JNFE 0127 發酵中pH 的影響Fig. 2 Effect of grains on pH in JNFE 0127 fermentation
2.2 大豆和苦蕎質量比對發酵豆乳NK 活力的影響
大豆與苦蕎按不同質量比復配后進行發酵,NK活力的結果見圖3。隨著苦蕎添加量的增加,發酵液中的酶活力出現峰值。 當大豆與苦蕎質量比為2∶1時,發酵液中NK 活力最高,可達到4 460.28 U/mL;與純大豆發酵液相比,酶活力提高了61.42%。 由此得出,復配谷物有利于提高發酵豆乳酶活力,與其他研究者的結論一致[16]。 然而隨著苦蕎添加量繼續增加,發酵體系中酶活力逐漸降低,在純苦蕎發酵體系中NK 活力只有2 018.10 U/mL。根據前人的研究可知,苦蕎本身含有單寧和蘆丁成分[31],單寧和蘆丁具有一定的抗菌性, 可抑制納豆芽孢桿菌的生長,進而使產酶量降低[32]。 此外,酸性環境以及缺乏某些生長補充劑也會限制其生長速度[33]。

圖3 谷物質量比對發酵豆乳酶活力的影響Fig. 3 Effects of grain mass ratios on enzyme activity of fermented soybean milk
2.3 大豆和苦蕎質量比對發酵豆乳pH 及酸度的影響
由于發酵體系中氮源含量的不同以及JNFE 0127 生長狀態的差異,發酵前后的pH 和酸度也會有所差異。如圖4(a)所示,發酵前各體系的pH 無差異, 均為6.5~6.6, 發酵后各體系的pH 降至5.6 左右,并且隨著苦蕎添加量的增加,發酵后體系的pH呈現下降的趨勢, 純大豆和純苦蕎體系發酵后pH分別為5.74 和5.52。 另外,由圖4(b)可知,純苦蕎發酵液的酸度最大(25.3 °T),與純大豆的酸度相比較,提高了27.78%。 一方面,苦蕎含有較多氨基酸類、多酚類等酸性物質,會使酸度升高[34];另一方面,在發酵過程中微生物產生的酶(纖維素酶、蛋白酶等)能打破多酚和其他取代基之間的聯系以釋放可溶性及游離性酚類化合物,也會使發酵液酸度升高[35]。

圖4 谷物質量比對發酵豆乳pH 及酸度的影響Fig. 4 Effects of grain mass ratios on pH and acidity of fermented soybean milk
2.4 大豆和苦蕎質量比對發酵豆乳風味物質的影響
2.4.1 大豆和苦蕎質量比對發酵豆乳揮發性鹽基氮質量濃度的影響 揮發性鹽基氮包含氨類、甲胺、二甲胺、三甲胺等化合物,通常是判斷納豆芽孢桿菌發酵豆乳不良風味的一項重要指標。 在JNFE 0127 發酵過程中,產生的蛋白酶或微生物分解體系含有的蛋白質及發酵過程中產生的游離氨基酸,會釋放揮發性氨類物質,產生不良氣味[36-37]。 結果如圖5 所示, 發酵液中揮發性鹽基氮質量濃度隨苦蕎添加量增加先降低后穩定。 在大豆與苦蕎質量比為1∶3 時,發酵體系中揮發性鹽基氮質量濃度最低,為2.63 mg/dL, 與純大豆發酵體系相比, 降低了39.12%;另外,大豆與苦蕎質量比為2∶1 的復配體系中,揮發性鹽基氮質量濃度為3.45 mg/dL,相比純大豆發酵降低了20.14%。 這與董岳峰的研究結果類似[12],其研究表明苦蕎中淀粉糖含量較高,在納豆芽孢桿菌發酵過程中添加苦蕎,可以提高復配發酵體系中的碳氮比,從而減少游離氨形成,改善風味[12]。
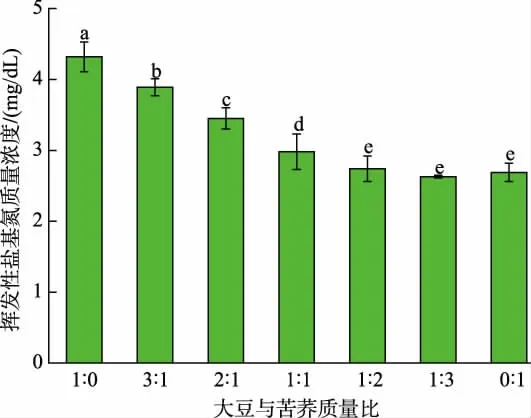
圖5 谷物質量比對發酵豆乳揮發性鹽基氮質量濃度的影響Fig. 5 Effects of grain mass ratios on volatile base nitrogen mass concentration in fermented soybean milk
2.4.2 發酵豆乳的電子鼻分析結果 納豆芽孢桿菌發酵的不良風味主要來自大豆本身的豆腥味和其發酵過程中產生的揮發性鹽基氮、生物胺等不良風味物質,電子鼻可以快捷地測定其不良風味的主要來源,有助于進行針對性改良。 結果如圖6 所示,大豆與苦蕎不同復配發酵體系的電子鼻測定數據主要有4 個峰位, 保留時間-色譜柱號分別為16.54-1、50.67-1、19.99-2、61.02-2。 經過Alphasoft軟件分析,16.54-1、19.99-2 對應氨氮類化合物,是發酵豆乳中不良風味的主要來源;50.67-1、61.02-2對應己醛等豆腥味的主要來源物質。 隨著苦蕎添加量的增加,4 種主要物質的峰面積逐漸降低,即含量逐漸降低。 該結果表明大豆與苦蕎復配可以有效改善發酵豆乳的不良風味,與上述測定的揮發性鹽基氮結果一致。

圖6 不同復配發酵體系所產豆乳的電子鼻分析Fig. 6 Electronic nose analysis of fermented soybean milk with different compound fermentation system
2.4.3 發酵豆乳的主成分分析結果 由圖7 可知,主成分1 和主成分2 累積貢獻率高達99.690%。 另外,如果兩樣品之間的距離越近,表示其整體的氣味信息越接近,差異越小,反之則越大。 各組在PCA分布圖中空間相對獨立,可以很好地區分不同復配發酵體系,也進一步說明不同發酵體系中風味特征存在差異。 其中,純大豆發酵體系和大豆與苦蕎質量比1∶3 的復配發酵體系風味信息較為相近, 大豆與苦蕎質量比為2∶1 和3∶1 的復配發酵體系風味信息較為相近,純苦蕎發酵體系與其他發酵體系整體風味信息差異較大。 此外,沿PC1 軸自左向右分布的樣品中,苦蕎添加量依次增加。 結合電子鼻及揮發性鹽基氮測定結果可知,第一主成分為可接受的風味,第二主成分為異味。
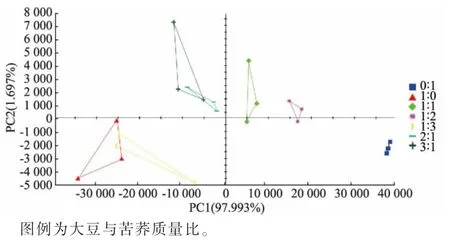
圖7 不同組別發酵豆乳的信號強度主成分分析Fig. 7 Principal component analysis of signal strength of fermented soybean milk in different groups
2.5 大豆和苦蕎質量比對發酵豆乳抗氧化活性的影響
通過測定發酵液總抗氧化能力、DPPH 自由基清除能力以及亞鐵離子螯合能力來評價不同組的抗氧化水平。 結果如表1 所示,大豆與苦蕎質量比為2∶1 時,發酵豆乳的總抗氧化能力(16.34 mmol/L)、DPPH 自由基清除能力(93.02%)以及亞鐵離子螯合能力(56.43%)均顯著高于純大豆及純苦蕎發酵體系, 與純大豆發酵體系相比分別提高96.39%、14.51%、34.17%。苦蕎中含有大量的酚類物質,隨著發酵體系中苦蕎的增加,酚類物質含量增加[38]。 Liu等發現結合酚不能被人體內的酶系統直接消化,經過微生物發酵后可發揮更高的生物活性[39]。 Caizhi等進一步研究表明酚類物質是植物性食品中的主要抗氧化劑,主要以與纖維素、胺和脂質部分結合的共軛形式存在[40];發酵過程能夠將共軛酚類化合物轉化為游離形式,從而增強抗氧化活性[29]。

表1 不同復配發酵體系所得豆乳的抗氧化活性Table 1 Antioxidant properties of soybean milk with different compound fermentation system
3 結 語
通過添加苦蕎、糙米、薏米、藜麥、燕麥粉可明顯提高JNFE 0127 液體發酵的產酶能力,其中苦蕎效果最好,當添加質量分數為3%時,產酶量由原來的5 803.12 U/mL 提高到23 326.23 U/mL。 在發酵豆乳制備中,隨著苦蕎添加量的增加,酶活力出現峰值,揮發性鹽基氮質量濃度呈現先降低后維持穩定的趨勢, 抗氧化活性呈現先升高再降低的趨勢。當大豆與苦蕎質量比為2∶1 時, 揮發性鹽基氮質量濃度可降低到3.45 mg/dL, 比純大豆發酵得到的發酵豆乳降低20.14%; 總抗氧化能力、DPPH 自由基清除能力以及亞鐵離子螯合能力分別提高到16.34 mmol/L、93.02%和56.43%, 也均高于純大豆發酵得到的發酵豆乳。 該研究在確保NK 活力足夠的條件下,為開發產品風味更易被消費者接受、食用更方便的納豆食品提供借鑒。
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
青年歌聲(2018年5期)2018-10-29 03:18:40
廣東飼料(2016年3期)2016-12-01 03:43:12
汽車觀察(2016年3期)2016-02-28 13:16:26
林業與生態(2016年2期)2016-02-27 14:24:01
