梵凈山自然保護區珙桐萌蘗繁殖在種群數量中的作用
2023-08-14 06:25:50盧志宏譚廷鴻李海波張曉東楊傳東
西北植物學報 2023年6期
盧志宏,漆 江,代 松,譚廷鴻,李海波,張曉東,楊傳東*
(1 廣西農業職業技術大學,南寧 530001;2 廣西生態工程職業技術學院,廣西柳州 545004;3 銅仁市林業科學院,貴州銅仁 554300;4 銅仁學院 貴州銅仁 554300;5 貴州省梵凈山自然保護區管理局,貴州江口 554400)
珙桐(Davidiainvolucrata)是中國特有的第三紀孑遺物種,有活化石之稱,為中國一級重點保護珍稀瀕危植物[1-4],目前僅零星分布于中國西南地區[5-9]。珙桐頭狀花序,無花瓣,而基部兩片大型苞片呈花瓣狀,遠觀如飛翔的鴿子,故稱作鴿子花;珙桐種子外殼堅硬,透水透氣性差,導致自然更新困難[1,4,10]。
珙桐具有種子和萌蘗兩種繁殖策略,二者之間存在著重要的權衡關系,萌蘗植物種子質量低、繁殖能力差、種子庫小,導致產生的實生苗少,因此萌蘗繁殖就顯得尤為重要,是一種重要補充策略[11]。萌蘗植物具有在原地快速恢復和更新的能力,在種子更新失敗時,萌蘗繁殖對維持種群穩定性具有十分重要的意義,表現為“駐留—定居”的權衡關系[12],成年個體的萌蘗行為則暗示了其潛在的持續性[13]。
劇烈干擾破壞種群時,萌蘗繁殖可以迅速使種群得到更新,較短時間恢復原先狀態[12,14],稱為“駐留生態位”效應[13],這種效應能夠維持種群競爭優勢,提高抗擾動能力[15-17],有利于在干擾發生后占據資源和空間[18],對維持種群穩定具有重要的生活史意義[13,19-20]。珙桐種群主要以萌蘗更新為主,實生苗(種子繁殖)極少[7-9]。珙桐依靠種子的自然更新十分困難,多數為萌蘗苗,可能和生境覆蓋物、自身特性有關[21-22]。梵凈山珙桐種群中萌蘗苗多發生在倒木、損傷的大樹根部,無損傷植株很少,而實生幼苗稀缺[9],幼苗、幼樹死亡率較高,幼苗和中齡級個體的不足是珙桐瀕危的重要原因[23-24],萌蘗繁殖是一種重要的更新方式[25]。
從現有的關于珙桐種群數量動態方面的研究來看,主要以萌蘗苗或實生苗(以基株為單位)數據進行種群動態分析,而忽略了兩種繁殖方式的區別與聯系,本研究主要探討兩種繁殖方式的徑級(齡級)結構、存活曲線及生存狀況,闡述萌蘗繁殖和種子繁殖(以基株為單位)在維持珙桐種群數量變化中的作用。
1 研究地概況
梵凈山國家級自然保護區位于貴州省東北部,地處亞熱帶,具有亞熱帶季風山地濕潤氣候特征,地理位置為108°45′5″-108°48′30″E,27°49′50″-28°01′30″N,境內最高點為鳳凰頂,海拔2 572 m,為武陵山脈的主峰。梵凈山自然保護區年均氣溫5.0~17.0 ℃,年均無霜期225~256 d,年均降水量2 500 mm,相對濕度年均在80%以上,保護區內植被覆蓋度在80%以上,海拔由低至高的植被景象有4個較為明顯的垂直帶譜,海拔700~1 300 m為常綠闊葉林帶,常見喬木樹種以白花樹(Styraxhypoglauca)、缺萼楓香樹(Liquidambaracalycina)、白辛樹(Pterostyraxpsilophylla)、中華槭(Acersinense)、野核桃(Juglanscathayensis)、水青樹(Tetracentronsinensis)、水青岡(Faguslongipetiolata)、厚皮栲(Castanopsischunii)和石櫟(Lithocarpusglaber)等為主。
2 研究方法
2.1 樣地調查
2019年7月13-21日在梵凈山自然保護區標水巖山谷開展野外實地調查,調查范圍:108°40′22″-108°40′26″E,27°57′38″-27°57′41″N,海拔963 m,進行每木調查,記錄高度、數量、胸徑(DBH)、冠幅等指標,共記錄實生苗57株(萌蘗苗400株),其胸徑1~56 cm,樹高0.5~20 m,冠幅0.2~10 m。
2.2 徑級結構劃分
珙桐為國家一級重點保護植物,因不能測定每株的具體年齡,所以采用常用的“空間推時間”的方法,用立木的胸徑徑級結構代替齡級研究珙桐種群動態[26]。每木調查以基株為單位進行統計,為更深入了解萌蘗繁殖和種子繁殖在珙桐種群數量中的作用,有萌蘗枝條的則同時記錄每個枝條的直徑和高度。
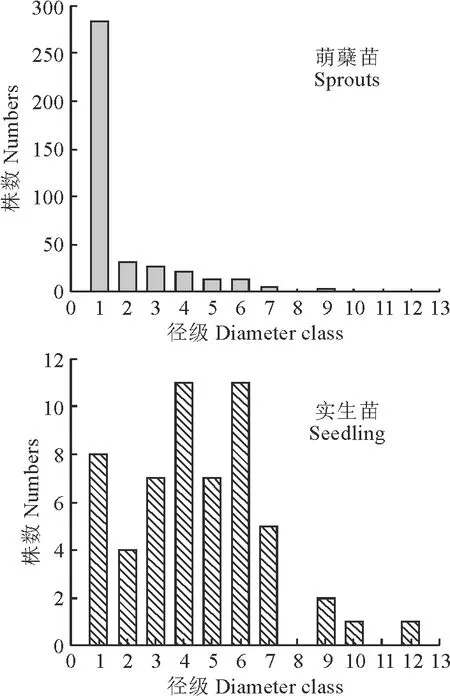
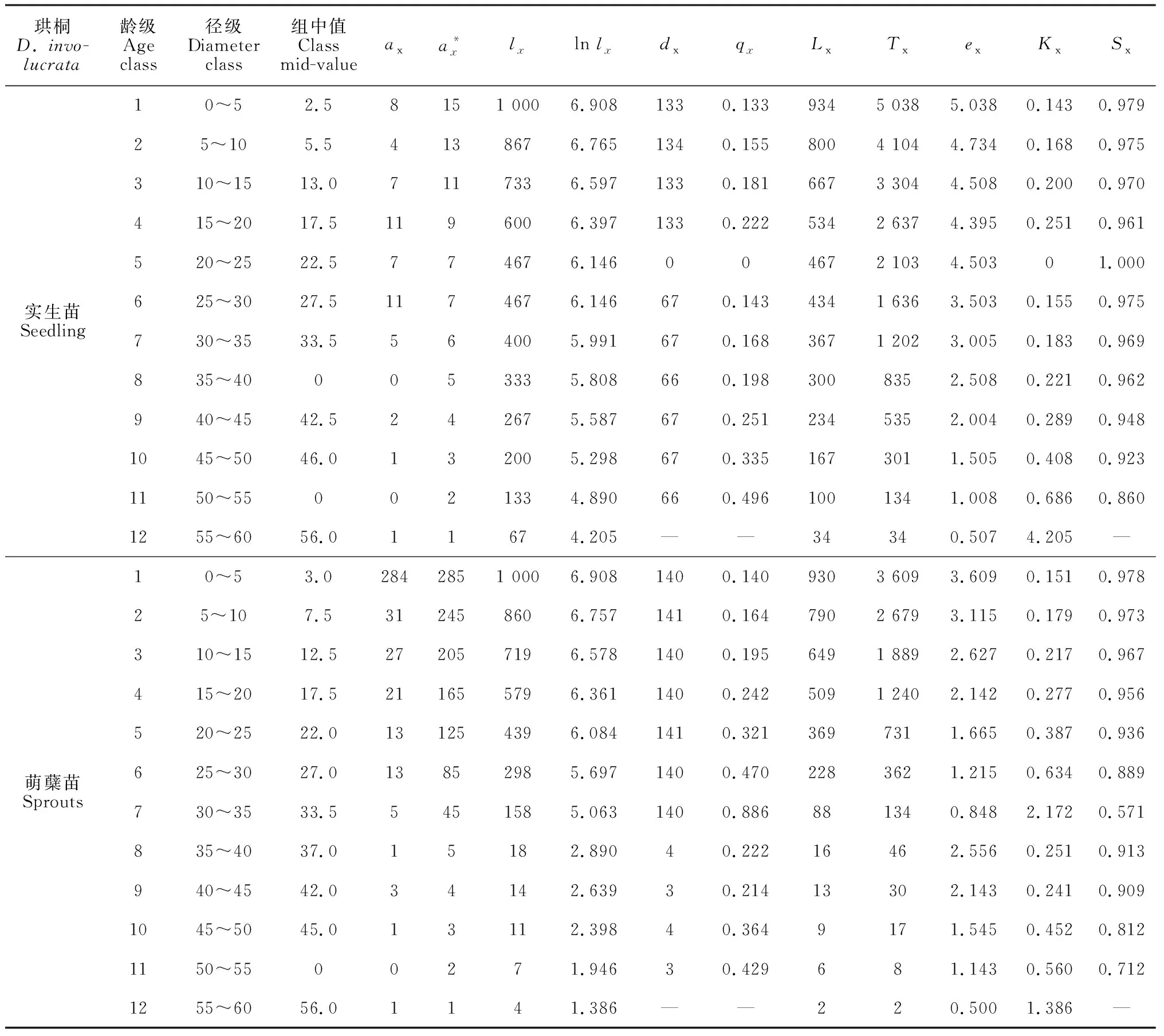
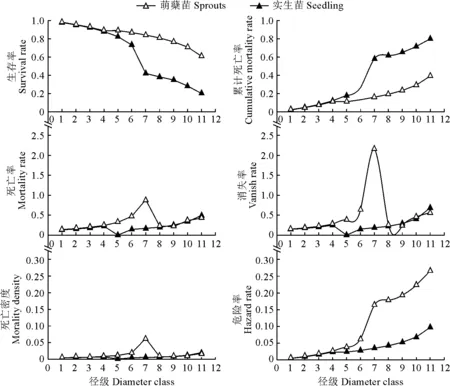
根據調查研究所獲得的數據以及珙桐的生活史,將珙桐種群分為12個胸徑徑級。0 采用種群徑級結構的動態指數對梵凈山自然保護區珙桐種群年齡結構進行定量的分析[29]。公式為: (1) (2) 式中:Vn為種群從n(n=1,2,…,k,k=11)到n+1徑級(齡級)的個體數量變化動態;Vpi為種群徑級(齡級)結構的數量變化動態;Sn為第n個徑級(齡級)的個體數;Sn+1為第n+1個徑級(齡級)的個體數;max(……)為取括號中數列的最大值;-1≤Vn≤1,Vn分別取正、0、負值時分別表示種群內兩相鄰的徑級(齡級)個體數的增長、穩定、衰退的動態關系。-1≤Vpi≤1,Vpi分別取正、0、負值時分別表示整個種群徑級(齡級)結構的數量增長、穩定、衰退的動態關系。Vsepi為萌蘗苗(以基株為單位);Vspi為萌蘗苗種群數量動態。 當將未來的外部干擾考慮進來時, (3) 式中:k為種群的徑級數;Vn、Vpi、Sn代表的意義均與上述相同。 標水巖珙桐靜態生命表編制,由于靜態生命表反映的是種群多個世代重疊的年齡動態,并不是對這個種群全部生活史的追蹤,因此經常會在實際應用中出現死亡率為負數的情況,鑒于此,常采用勻滑技術對各個徑級(齡級)的個體數目進行勻滑修正[30],然后使用勻滑后的數據編制種群靜態生命表,本研究也采用這種勻滑技術。 lx=(ax/a0)×1000 (4) Sx=lx+1/lx (5) dx=lx-lx+1 (6) qx=(dx/lx)×100% (7) Lx=(lx+lx+1)/2 (8) (9) ex=Tx/lx (10) Kx=lnlx-lnlx+1 (11) 根據以上各項數據處理,繪制珙桐種群的靜態生命表。 采樣種群生存率函數S(i)、累計死亡率函數F(i)、死亡密度函數f(ti)和危險率函數λ(ti)等4個函數進行珙桐種群生存分析。4個函數計算公式分別為: S(i)=S1×S2…×Si (12) F(i)=1-S(i) (13) f(ti)=(S1-Si-1)/hi (14) λ(ti)=2(1-Si)/[hi(1+Si)] (15) 式中:Si為存活率;hi為徑級(齡級)寬度。 根據上述值,繪制出珙桐種群的生存率曲線、累計死亡率曲線、死亡密度曲線以及危險率曲線[31-32]。 由圖1可以看出,統計萌蘗枝條后的種群數量徑級(齡級)結構大體上呈現不斷下降的趨勢,第1徑級的個體數量共有284株,占總數的71.00%;1~2徑級(齡級)為幼齡階段,個體數共有315株,占總數的78.75%,3~5徑級(齡級)為中齡階段,個體數共有61株,占總數的15.25%,6~12徑級(齡級)為成齡個體,僅有24株,占總數的6.00%。實生苗(以基株為單位)徑級(齡級)結構大體上呈對稱分布,1~2徑級(齡級)為幼齡階段,個體數共有12株,占總數的21.45,3~5徑級(齡級)為中齡階段,個體數共有25株,占總數的43.86%,6~12徑級(齡級)為成齡階段,個體數共有20株,占總數的23.09%,8和10徑級(齡級)沒有個體。這種現象說明幼齡個體能穩定成長,成齡個體成長不穩定。萌蘗繁殖是對種子繁殖的一種補充,珙桐的萌生枝條較多,且在野外調查過程中,個別個體枝干上半部分死亡,或有樹洞,中空的現象,而萌蘗枝條可以維持該個體的生存。 圖1 珙桐種群徑級結構分析Fig.1 Diameter structure of Davidia involucrata population 根據陳曉德[27]推導的動態指數,計算得到珙桐種群實生苗(以基株為單位計算)和萌蘗苗種群各個相鄰徑級間的數量動態變化。 由表1可知,實生苗(以基株為單位計算)種群的第2,3,5,8,11徑級(齡級)的Vn都是負數,表明珙桐種群在這5個徑級(齡級)更新比較差,個體數量呈衰退的趨勢。但整個種群徑級(齡級)結構指數Vpi=25.34%,將未來的外部干擾考慮進來時Vpi=2.30%>0,因此現階段珙桐種群在整體上是增長的。 表1 珙桐種群的數量動態Table 1 Quantitative dynamics of D. involucrata populations 計算萌蘗苗種群的第8和11徑級(齡級)的Vn是負數,第5徑級(齡級)的Vn是0,表明這3個徑級(齡級)的更新較差,呈衰退趨勢。整個種群徑級(齡級)結構指數Vpi=71.51%,將未來的外部干擾考慮進來時Vpi=6.50%>0,因此現階段珙桐種群在整體上是增長的。通過對實生苗和萌蘗苗的對比分析,幼齡和中齡階段的更新明顯不同,甚至是相反的,而在成齡階段6~11徑級(齡級)幾乎相同,可能是中齡階段的珙桐受到外界干擾,如地上部分死亡,倒伏,樹洞,中空等現象發生,通過萌蘗繁殖維持個體存活,直到恢復原先狀態。現階段的珙桐種群表現為增長型種群,萌蘗繁殖策略在維持種群穩定方面具有重要的作用。 根據標水巖珙桐胸徑數據經分析編制萌蘗苗和實生苗靜態生命表(表2)。由表2可以看出,實生苗的幼齡和成年個體較少,中齡個體較多。ex反映x徑級內個體生命期望的壽命,第1徑級的數值最高,為5.038,隨著徑級(齡級)的增加而降低;可能是因為珙桐種子萌發困難,僅在適合的環境下容易萌發并建成個體,而隨著個體的生長,容易受到環境干擾。存活曲線能夠反映種群個體在各個徑級(齡級)的存活狀況,經SPSS 25分析,實生苗的冪函數和指數方程式分別為: 表2 珙桐種群靜態生命表Table 2 Static lifetable of D. involucrata populations y=7.601x-0.158,R2=0.684,F=21.619 y=7.464e-0.038x,R2=0.886,F=77.557 指數方程的R2和F值均大于冪函數的,實生苗生存曲線趨向于DeeveyⅡ型曲線。 萌蘗苗的幼齡個體數目相對較多,隨著徑級(齡級)的增大個體數不斷降低,第1徑級的個體生命期望最高,為3.609,在第1~7和8~12徑級(齡級)內隨著徑級(齡級)增加而降低,第7徑級(齡級)為拐點。萌蘗苗的冪函數和指數方程式分別為: y=11.022x-0.608,R2=0.650,F=18.581 y=10.56e-0.149x,R2=0.891,F=81.690 指數方程的R2和F值均大于冪函數的,萌蘗苗存活曲線趨向于DeeveyⅡ型曲線。因此,標水巖珙桐種群存活曲線屬于DeeveyⅡ型曲線,各個徑級(齡級)具有相同的死亡率,成齡生長趨于穩定。 由圖2可以看出:實生苗和萌蘗苗的生存率曲線單調下降,累計死亡率單調上升,死亡率和消失率曲線走向一致,消失率曲線波動大一些;消失率曲線最大值均出現在第7徑級(齡級),死亡率曲線最大值均出現在第11徑級(齡級)。死亡密度曲線較為平緩,危險率曲線波動較大,尤其是萌蘗苗的變化幅度更大,而前期的危險率幾乎相同,后期差異增大,說明萌蘗苗的作用主要在5~7徑級(齡級)維持種群數量,而后期對種群穩定的作用較小。 圖2 珙桐種群的生存分析曲線Fig.2 Survival analysis curves of D. involucrata populations 從生存率函數、死亡密度函數、死亡率密度和危險率函數分析種群結構和數量動態,實生苗各個徑級(齡級)死亡率相同,種群比較穩定;萌蘗苗前期下降、中后期穩定的特點;珙桐在中齡和成齡階段受擾動后,萌蘗繁殖大量增加幼齡(第1,2徑級)個體數量,在原地維持和恢復,保持種群數量的穩定。 一般認為萌蘗繁殖可以維持種群的穩定性[12,33-34],成齡階段萌蘗繁殖暗示著種群具有潛在的可持續性[20],而多數物種在成齡階段喪失了萌蘗繁殖能力。在實地調查中,珙桐的萌蘗現象主要集中在成齡個體,實生苗幼齡階段未發現萌蘗現象,成齡個體受到干擾時,萌蘗繁殖可以就地恢復,維持種群數量,有利于種群的穩定性。 一般認為萌蘗植物的種子繁殖能力有限,種子庫較小和種子質量低,實生苗少,萌蘗更新是一種補充策略[35]。珙桐種子外殼堅硬,自然萌發率極低,實生苗極少,因此其萌蘗更新成為其一種重要的繁殖策略。 從珙桐種群結構來看,羅柏青等分析滇東北珙桐群落時指出,珙桐更新形式以萌蘗為主,極少見實生苗,具有衰退跡象[36]。珙桐有性繁殖能力很弱,幼樹幾乎都是靠萌蘗產生的萌蘗苗[23],在不適于種子萌發的環境下,萌蘗維持了珙桐種群的繁衍[8]。 吳明開等在研究梵凈山珙桐種群時指出,珙桐大樹根部萌條居多,實生幼苗稀缺,為衰退型種群[9]。并指出前期銳減、中期穩定和后期衰退,認為幼苗和中齡級個體不足是珙桐瀕危的重要原因,本調查也呈現出前期減少,中期穩定的規律。對比徑級,可以發現前者調查的徑級Ⅸ為40~45 cm,本研究最大徑級Ⅻ為55~60 cm,超過前者3個徑級,因此在相似的研究區域內,珙桐種群數量動態規律與吳明開等的結論差異較大,為增長型種群。造成相同研究區域內差異較大的原因可能是因調查方式不同引起的,本研究在標水巖珙桐分布區域內進行了每木調查,而前者的研究采取隨機大樣方進行調查,二者的差異可能由樣本抽取方法導致。 雖然,光葉珙桐萌生力較強,但萌條多見于倒木、樹樁、根際等,生長狀況良好,無損傷的植株鮮見萌生苗[25]。在實地調查過程中,發現了2個極端案例,位于溝谷底部一株胸徑40 cm的珙桐根部受水流沖刷,直徑幾毫米的細小的萌蘗枝條極為密集,而在地勢較平坦的地方另一株在1.5 m位置有3個直徑20 cm左右的分支,基部沒有萌蘗現象,可能是由于萌蘗繁殖恢復了原有個體的空間資源,即有利于維持種群數量穩定。 陳光平等在分析寬闊水自然保護區珙桐種群結構時指出成年及老年珙桐比重大,實生苗和幼樹嚴重缺乏,萌蘗苗多而成活率很低,總體屬于衰退型種群[37]。萌蘗苗成活率低,維持了干擾前個體的競爭優勢,有利于恢復原有的生存空間,對種群穩定具有重要意義。喻理飛認為過度干擾是造成珙桐林衰退的主要原因,可通過自身更新恢復種群穩定[25],同時也說明珙桐的萌蘗繁殖策略是進化選擇的必然結果。 通過分析珙桐2種繁殖策略下現存的種群結構,研究表明萌蘗繁殖和種子繁殖在幼齡階段存在明顯差異,萌蘗繁殖是種子繁殖的重要補充策略,萌蘗繁殖有利于維持種群數量,而在成齡階段兩種策略作用相似。2.3 種群徑級結構的數量變化動態分析方法
2.4 靜態生命表編制
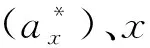
2.5 生存分析
3 結果與分析
3.1 徑級結構分析
3.2 種群年齡結構的數量動態變化

3.3 靜態生命表的編制與存活曲線分析
3.4 生存分析
4 討 論
5 結 論
