新品種“美賜”半同胞子代種苗特性與優(yōu)株選擇
2023-08-11 09:01:08陳泉佐趙賢生焦玉鵬張棋淞楊新麗楊小軍付順華
特種經(jīng)濟(jì)動(dòng)植物 2023年8期
●陳泉佐 趙賢生 焦玉鵬 張棋淞 楊新麗 楊小軍 姜 磊 張 磊 付順華※
(1.浙江農(nóng)林大學(xué) 林業(yè)與生物技術(shù)學(xué)院 浙江 杭州 311300;2.富陽(yáng)區(qū)農(nóng)業(yè)技術(shù)推廣中心經(jīng)作站 浙江 杭州 311400;3.浦江縣俊果研究所 浙江 金華 322329;4.杭州富陽(yáng)恒浩家庭農(nóng)場(chǎng) 浙江 杭州 311402;5.浙江省桐鄉(xiāng)第一中學(xué) 浙江 嘉興 314500)
掌葉覆盆子(Rubus chingiiHu)是薔薇科懸鉤子屬的一種藤狀落葉灌木,又稱掌葉復(fù)盆子、華東覆盆子,主要分布在中國(guó)華東地區(qū)。其成熟果實(shí)鮮紅美艷,口感酸甜,風(fēng)味獨(dú)特。近成熟青果制干即為有悠久使用歷史的中藥材覆盆子。藥理和化學(xué)成分研究表明,掌葉覆盆子根、葉、果均具有藥用價(jià)值,有益腎、固精、縮尿、壯陽(yáng)及養(yǎng)肝明目等作用[1],可促進(jìn)消化、緩解腹瀉、補(bǔ)充鐵質(zhì)[2],可用于治療細(xì)胞的炎癥反應(yīng)[3]。掌葉覆盆子葉茶多酚含量接近于茶葉,且口感甘甜,有降血脂效果,制成茶劑或其他食品可滿足高血脂人群需求[4],并可使婦女子宮肌和骨盆肌肉保持正常,提高嬰兒母乳營(yíng)養(yǎng)價(jià)值,促進(jìn)產(chǎn)婦母乳分泌[5]。掌葉覆盆子葉的乙醇提取物有顯著的抗血栓作用,分離得到的山奈酚、槲皮素和椴樹(shù)苷能明顯延遲血液中血漿復(fù)鈣時(shí)間[6]。同時(shí)掌葉覆盆子果實(shí)含有豐富的氨基酸和維生素,有抗衰老、保健、美容功效[7-9],是一種亟待開(kāi)發(fā)的健康功能植物。
20世紀(jì)90年代,國(guó)內(nèi)懸鉤子屬植物受到人們關(guān)注,得到初級(jí)開(kāi)發(fā)利用。人們開(kāi)始對(duì)野生懸鉤子植物進(jìn)行馴化引種,并開(kāi)展生物學(xué)基礎(chǔ)、栽培性狀等研究,其中重點(diǎn)開(kāi)展對(duì)掌葉覆盆子的研究。從栽植環(huán)境、移栽時(shí)間和密度、苗期管理措施方面,探索掌葉覆盆子的栽培模式[1];開(kāi)展施肥用量和施肥時(shí)間等研究,了解合適施肥條件[10];開(kāi)展種苗繁殖和組培快繁研究,了解合適采種期、種子貯藏時(shí)間、催芽方式[11-13]和組織培養(yǎng)合適條件[7,14-15];從化學(xué)成分和藥理作用的深入研究中,發(fā)現(xiàn)果實(shí)、葉片中多種類的新型營(yíng)養(yǎng)物質(zhì),包括糖類、有機(jī)酸類、維生素類成分和揮發(fā)性成分[16-17],以及抑制原發(fā)性肝癌[18]、祛除黃褐斑等新功能[19]。
目前掌葉覆盆子良種選育已有一些初步研究[20-21],主要是通過(guò)比較不同植株個(gè)體生長(zhǎng)開(kāi)花結(jié)實(shí)特性的變異選擇優(yōu)株,但少有進(jìn)一步培育出生產(chǎn)上可利用的新品種的報(bào)道,涉及以皮刺為主要育種性狀的掌葉覆盆子良種選育研究更是未見(jiàn)報(bào)道。半同胞子代有一半的遺傳信息來(lái)源于母本,故能夠較好地繼承母本的優(yōu)良特性,同時(shí)又有來(lái)自不同父本的遺傳信息,會(huì)產(chǎn)生新的基因型和優(yōu)良表現(xiàn)型。在半同胞子代群體中開(kāi)展優(yōu)株選擇,可以有效選擇到綜合父母本雙親優(yōu)良特性的個(gè)體,進(jìn)而能培育性狀更優(yōu)良的品種或品系。本試驗(yàn)以在生產(chǎn)上開(kāi)始推廣應(yīng)用的少皮刺型新品種“美賜”半同胞子代(以下簡(jiǎn)稱“美賜”)的種子為材料,通過(guò)觀測(cè)“美賜”種子特性、種子發(fā)芽特性、苗期形態(tài)特征、分枝特征和皮刺分布,了解其種子種苗生長(zhǎng)特性,并開(kāi)展苗期少(無(wú))刺優(yōu)良單株選擇,為其后續(xù)品種升級(jí)換代、保純防衰和創(chuàng)制種質(zhì)資源提供參考依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
試驗(yàn)材料“美賜”種子從以掌葉覆盆子新品種“美賜”植株為母本的自由授粉成熟果實(shí)中獲得,普通種子從種植基地中的掌葉覆盆子栽培植株上獲得。
2020年5月和2021年5月兩次在金華市浦江縣花橋鄉(xiāng)掌葉覆盆子種植基地,選擇生長(zhǎng)良好的“美賜”植株和普通掌葉覆盆子植株分別取樣。摘取果型飽滿、顏色正常紅艷的成熟果實(shí),分別裝入保鮮袋,做好標(biāo)記,帶回實(shí)驗(yàn)室即時(shí)進(jìn)行稱重,同時(shí)記錄果實(shí)形態(tài)性狀。隨后對(duì)果實(shí)進(jìn)行破碎,用水洗法去除果肉,分離種子,將得到的種子放在陰涼處風(fēng)干。干燥后進(jìn)行千粒重測(cè)定,隨后將種子進(jìn)行貯藏。
1.2 研究方法
1.2.1 試驗(yàn)地整理 2021年4月,在浙江農(nóng)林大學(xué)平山試驗(yàn)基地的省級(jí)林木種質(zhì)保育基地大棚內(nèi)設(shè)置試驗(yàn)圃地。圃地土壤深耕,后將圃地雜草雜物清理,殺菌消毒,清除土壤病蟲(chóng)害。再在土壤上覆蓋一層2 cm厚的泥炭土,隨后噴水至圃地土壤完全濕潤(rùn)。
1.2.2 種子播前處理與播種 對(duì)2020年5月所采的貯藏后“美賜”種子,于播種前放入清水中浸泡4 h,使種子吸收足夠的水分,促進(jìn)種子萌發(fā)。圃地開(kāi)播種溝,以條播方式進(jìn)行播種。完成播種后,種子不覆土,輕度噴霧灑水濕潤(rùn),使種子充分接觸土壤。
1.2.3 苗期管理與病蟲(chóng)害防治 幼苗萌出后,觀察其生長(zhǎng)狀況的同時(shí),適時(shí)進(jìn)行水分補(bǔ)充。定期噴霧澆水,以土壤不干為度,進(jìn)行“美賜”苗期生長(zhǎng)性狀觀測(cè)和成苗期優(yōu)株選擇群體。
2021年5~6月,發(fā)現(xiàn)小苗葉片有被食葉昆蟲(chóng)啃食跡象(圖1),確認(rèn)是短額負(fù)蝗(Atractomorpha sinensisB.)成蟲(chóng)和菜粉蝶(Pieris rapaeL.)幼蟲(chóng),隨后使用蘇云金桿菌藥劑滅蟲(chóng),8~10 mL藥劑加水制成1.5 L混合溶液,葉面噴灑,數(shù)天后葉片被啃食現(xiàn)象消失。當(dāng)年8月,小苗葉面出現(xiàn)黃色病斑,使用鏈霉素藥劑進(jìn)行防治后,葉面黃色病斑逐漸減少。

圖1 苗期食葉昆蟲(chóng)及危害狀
1.2.4 性狀觀測(cè)
1.2.4.1 種子正常率與外觀形態(tài) 在2021年5月所采“美賜”種子、普通種子中分別隨機(jī)選取種子,用刀片小心切開(kāi)硬種皮,觀察種子內(nèi)部種胚結(jié)構(gòu)是否完整、正常,如果種胚白色、飽滿,認(rèn)為是正常種子;如果種胚黑色或棕色、干癟狀態(tài),則認(rèn)為是不正常種子。各隨機(jī)選取30粒種子作為一組,各重復(fù)3次,記錄觀測(cè)數(shù)據(jù)。種子正常率計(jì)算公式如下:
其中,n為正常種子數(shù),N為總解剖種子數(shù)。
使用電子游標(biāo)卡尺測(cè)量種子縱徑、橫徑和側(cè)徑。選擇“美賜”種子和普通種子若干,各從中隨機(jī)選取10粒種子,分別測(cè)定種子縱徑(縱軸方向)、橫徑(平行子葉且與縱軸垂直方向)和側(cè)徑(垂直子葉方向)[22]。
1.2.4.2 種子千粒重測(cè)定 參照《林木種子檢驗(yàn)規(guī)程》[23]進(jìn)行種子千粒重測(cè)定。從2021年5月所采“美賜”種子和普通種子中按檢驗(yàn)規(guī)程要求,各隨機(jī)選取3份1000粒種子,進(jìn)行稱重。
1.2.4.3 種子發(fā)芽率測(cè)定 從同期所采“美賜”種子和普通種子里,分別隨機(jī)選取相同數(shù)量的正常種子,使用無(wú)菌、透氣、柔軟的雙圈定性濾紙浸濕作發(fā)芽床,將種子按2 mm間隔排列在濾紙上,置透明玻璃培養(yǎng)皿中,觀測(cè)記錄發(fā)芽情況。種子發(fā)芽率計(jì)算公式如下:
其中,n為發(fā)芽種子總粒數(shù),N為試驗(yàn)種子總粒數(shù)。
1.2.4.4 1年生苗期生長(zhǎng)性狀與皮刺觀測(cè) 2021年9~11月,對(duì)株高、冠幅、地徑、皮刺、分枝高等性狀在所建立的苗期觀測(cè)選擇群體中進(jìn)行特性觀察測(cè)量。使用鋼卷尺測(cè)量種苗株高、冠幅、分枝高,使用電子游標(biāo)卡尺測(cè)量種苗地徑。從皮刺在莖稈上的分布數(shù)量、密度及起點(diǎn)終點(diǎn)位置等觀測(cè)植株皮刺生長(zhǎng)特性,從分枝數(shù)、分枝級(jí)數(shù)和分枝起點(diǎn)等觀測(cè)種苗分枝特性。
1.2.5 優(yōu)株選擇 2021年12月,在“美賜”1年生苗群體中,以植株皮刺數(shù)少為選擇目標(biāo),制定選優(yōu)標(biāo)準(zhǔn),進(jìn)行優(yōu)株選擇,以皮刺性狀遺傳力預(yù)期值為0.60(無(wú)性繁殖時(shí)預(yù)期重復(fù)力為0.90),計(jì)算選擇差、選擇強(qiáng)度、選擇響應(yīng)和遺傳增益等遺傳育種參數(shù)。并對(duì)初選優(yōu)株進(jìn)行詳細(xì)調(diào)查,綜合評(píng)價(jià)各優(yōu)株特性。計(jì)算各遺傳參數(shù)。參考《林木遺傳學(xué)基礎(chǔ)》[24]和《森林遺傳學(xué)》[25],公式如下:
式中x為入選群體平均值,μ為群體平均值,σp為標(biāo)準(zhǔn)差,h2為預(yù)期遺傳力(無(wú)性繁殖時(shí)為預(yù)期重復(fù)力)。
優(yōu)株入選率的計(jì)算公式如下:
其中y為入選優(yōu)株數(shù),Y為總株數(shù)估計(jì)值。
1.3 數(shù)據(jù)處理及分析
借助Excel 2018、SPSS 22.0、Origin29.0等軟件統(tǒng)計(jì)處理數(shù)據(jù)和做圖,進(jìn)行顯著性檢驗(yàn),計(jì)算平均值、標(biāo)準(zhǔn)差、相關(guān)系數(shù)及育種參數(shù)等。
2 結(jié)果與分析
2.1 種子特性分析
2.1.1 種子正常率與種子千粒重分析 從表1可知,“美賜”種子正常率平均值為82.80%,而普通種子正常率平均值為70.00%。獨(dú)立樣本t檢驗(yàn)顯示,“美賜”種子正常率與普通種子存在顯著性差異(p=0.013),表明“美賜”種子正常率顯著高于普通種子。

表1 “美賜”種子與普通種子特性比較
從表1可知,“美賜”種子千粒重平均值為1.00 g,普通種子千粒重平均值為1.29 g,表明美賜種子千粒重小于普通種子;兩者的標(biāo)準(zhǔn)差都較小,美賜種子千粒重標(biāo)準(zhǔn)差是0.05 g,普通種子為0.03 g,表明兩者的種子千粒重在個(gè)體間差異并不顯著。
2.1.2 種子外觀形態(tài)及種子個(gè)體間形態(tài)差異比較 種子外觀形態(tài)及種子個(gè)體間形態(tài)差異觀測(cè)結(jié)果,見(jiàn)表2。

表2 “美賜”種子與普通種子外觀性狀比較
由表2可知,“美賜”種子外觀形態(tài)比普通種子略小。“美賜”種子縱徑、橫徑、側(cè)徑平均值為2.092,1.270,0.834 mm,普通種子的縱徑、橫徑、側(cè)徑平均值為2.265,1.492,0.981 mm,美賜種子縱徑、橫徑、側(cè)徑分別比普通種子小7.51%,14.88%,14.98%,這與“2.1.1”節(jié)中種子千粒重的研究結(jié)果一致。
從種子外觀形態(tài)個(gè)體間的差異看,“美賜”種子的個(gè)體間變異比普通種子大。“美賜”種子的縱徑、橫徑和側(cè)徑3個(gè)外觀性狀值的方差(0.017,0.031,0.012 mm2)、標(biāo)準(zhǔn)差(0.133,0.181,0.110 mm)和變異系數(shù)(6.36%,14.25%,13.19% )均明顯大于普通種子的相應(yīng)外觀性狀值的方差(0.005,0.004,0.012 mm2)、標(biāo)準(zhǔn)差(0.074,0.069,0.115 mm)和變異系數(shù)(3.26%,4.62%,11.72%),尤其是“美賜”種子的橫徑變異更大,最大值/最小值為1.77(1.70/0.96),變異系數(shù)達(dá)14.25%,普通種子的橫徑變異較小,最大值/最小值為1.17(1.60/1.37),變異系數(shù)僅為4.62%。反映出“美賜”種子的個(gè)體間分化比普通種子更大。
2.2 種子發(fā)芽特性及發(fā)芽率分析
利用2021年5月所采的“美賜”種子和普通種子進(jìn)行發(fā)芽試驗(yàn),結(jié)果見(jiàn)圖2。由圖2可知,在9月至翌年1月的秋冬季節(jié),“美賜”種子和普通種子經(jīng)過(guò)長(zhǎng)達(dá)近半年的發(fā)芽過(guò)程中,前3個(gè)月的發(fā)芽率都極低,表明此階段掌葉覆盆子種子休眠特性仍未完全解除,不易發(fā)芽。100 d后,無(wú)論“美賜”種子還是普通種子的發(fā)芽率均開(kāi)始明顯上升,但“美賜”種子發(fā)芽率更高。曼-惠特尼U檢驗(yàn)結(jié)果(p=0.000)顯示,“美賜”種子和普通種子的發(fā)芽率存在顯著差異,試驗(yàn)結(jié)束時(shí)觀測(cè),“美賜”發(fā)芽率達(dá)14.48%,而普通種子的發(fā)芽率僅4.21%。“美賜”種子發(fā)芽率高于普通種子,這個(gè)結(jié)果與上述“2.1.1”種子正常率的研究結(jié)果一致。

圖2 “美賜”種子與普通種子秋冬季發(fā)芽率比較
2021年春,利用2020年5月所采集的“美賜”種子進(jìn)行發(fā)芽試驗(yàn)觀測(cè),其結(jié)果如圖3所示。試驗(yàn)處理種子320粒最終發(fā)芽種子數(shù)為193粒,發(fā)芽率為60.31%,開(kāi)始發(fā)芽時(shí)間為第26天。4月5日的發(fā)芽種子數(shù)最多,為39粒,4月9日、4月15日的發(fā)芽種子數(shù)最少,均僅4粒,這可能與氣溫的明顯升降變化有關(guān)。結(jié)合圖2“美賜”種子秋冬季經(jīng)過(guò)150 d發(fā)芽率才達(dá)到14.48%,顯示種子采收貯藏后的翌年春季是“美賜”種子的較合適發(fā)芽時(shí)期。除了“美賜”種子是否已解除休眠因素外,光照、溫度等因素可能都是它不同季節(jié)發(fā)芽率差異的重要原因。
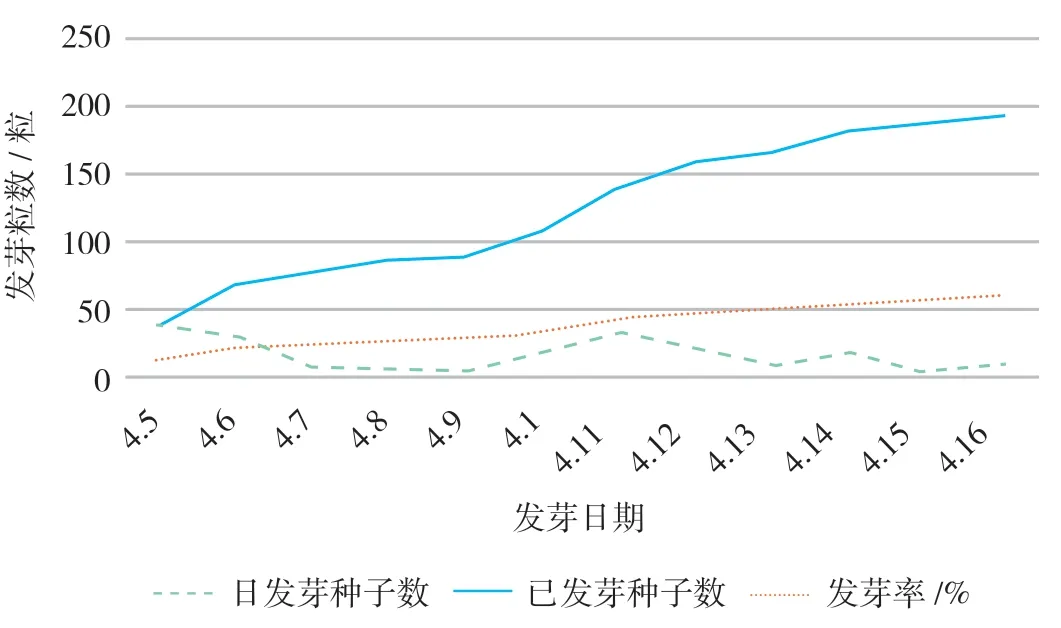
圖3 “美賜”種子春季發(fā)芽情況
2.3 “美賜”1年生苗期特性分析
2.3.1 “美賜”1年生苗生長(zhǎng)性狀及相關(guān)性 在“美賜”1年生苗的秋季生長(zhǎng)的時(shí)期Ⅰ(9月9日)、時(shí)期Ⅱ(10月14日),對(duì)株高、冠幅和地徑形態(tài)指標(biāo)分別進(jìn)行了觀測(cè),并分析了生長(zhǎng)性狀間的相關(guān)性,結(jié)果見(jiàn)表3。

表3 “美賜”1年生苗株高、冠幅和地徑生長(zhǎng)變化及性狀間相關(guān)性
從表3可知,在時(shí)期Ⅰ測(cè)定時(shí),“美賜”苗的株高、冠幅、地徑平均值分別為33.65 cm、21.48 cm、2.45 mm,到時(shí)期Ⅱ測(cè)定時(shí),“美賜”苗的株高、冠幅、地徑平均值分別為47.34 cm、41.10 cm、3.69 mm,時(shí)期Ⅱ“美賜”苗的株高、冠幅、地徑平均值分別比時(shí)期Ⅰ增加40.68%,91.34%,50.61%,表明在秋季這個(gè)階段,“美賜”1年生苗隨著溫度變涼適宜生長(zhǎng),形態(tài)建成較為快速。同時(shí),隨著個(gè)體形態(tài)建成,群體內(nèi)的個(gè)體分化也變得非常明顯,時(shí)期Ⅰ的標(biāo)準(zhǔn)差分別為10.22 cm、7.20 cm、0.83 mm,變化為時(shí)期Ⅱ的標(biāo)準(zhǔn)差13.68 cm、20.28 cm、1.41 mm,時(shí)期Ⅱ“美賜”苗的標(biāo)準(zhǔn)差分別比時(shí)期Ⅰ擴(kuò)大33.86%,181.67%,69.88%,其中以冠幅分化更大,地徑分化次之。
“美賜”1年生苗的生長(zhǎng)性狀株高、冠幅及地徑兩兩之間均存在極顯著正相關(guān)(表3)。其中株高與冠幅的相關(guān)系數(shù)為0.851(P<0.01),相關(guān)系數(shù)最大。株高與地徑的相關(guān)系數(shù)為0.473(P<0.01),冠幅與地徑的相關(guān)系數(shù)為0.451(P<0.01),相關(guān)系數(shù)中等。
2.3.2 “美賜”1年生苗分枝特征 隨著“美賜”1年生苗的生長(zhǎng)發(fā)育,其植株形態(tài)開(kāi)始發(fā)生劇烈變化(表4)。在較短的生長(zhǎng)階段內(nèi)由單莖分枝變?yōu)槎嗷患?jí)分枝莖并在一級(jí)分枝莖稈上再產(chǎn)生二級(jí)分枝。

表4 “美賜”1年生苗分枝特性
由表4可知,在1年生苗生長(zhǎng)的當(dāng)年,時(shí)期Ⅱ(10月4日)至?xí)r期Ⅲ(11月29日)時(shí)間段,“美賜”種苗出現(xiàn)分枝的植株比例由40.00%增加到了90.91%,其中,大多為基生一級(jí)分枝(即分生枝從近地面長(zhǎng)出),植株比例達(dá)到87.50%。同時(shí)植株個(gè)體分枝結(jié)構(gòu)也進(jìn)一步發(fā)生分化,在群體里表現(xiàn)不一,有部分植株出現(xiàn)一級(jí)分枝后開(kāi)始繼續(xù)出現(xiàn)二級(jí)分枝。
由表4可知,在產(chǎn)生分枝的“美賜”1年生種苗群體中一級(jí)分枝具1枝的植株占60.00%,具2枝的植株占33.33%,平均一級(jí)分枝數(shù)為1.47枝。同時(shí)該群體中,無(wú)二級(jí)分枝的植株占66.67%,具二級(jí)分枝1枝的植株占6.67%,具2枝的植株占26.67%,平均二級(jí)分枝數(shù)為0.60枝。表明“美賜”1年生苗的植株基本結(jié)構(gòu)是具1~2個(gè)一級(jí)分枝,較少二級(jí)分枝。
2.3.3 “美賜”苗期皮刺生長(zhǎng)特性分析
2.3.3.1 “美賜”苗期皮刺生長(zhǎng)的時(shí)間變化 “美賜”1年生種苗生長(zhǎng)期形態(tài)發(fā)生快速變化的同時(shí),皮刺生長(zhǎng)也有較大變化。在時(shí)期Ⅰ(9月9日)、時(shí)期Ⅲ(11月29日)兩個(gè)時(shí)間點(diǎn),對(duì)“美賜”種苗的皮刺分布情況進(jìn)行了觀測(cè),結(jié)果見(jiàn)表5。
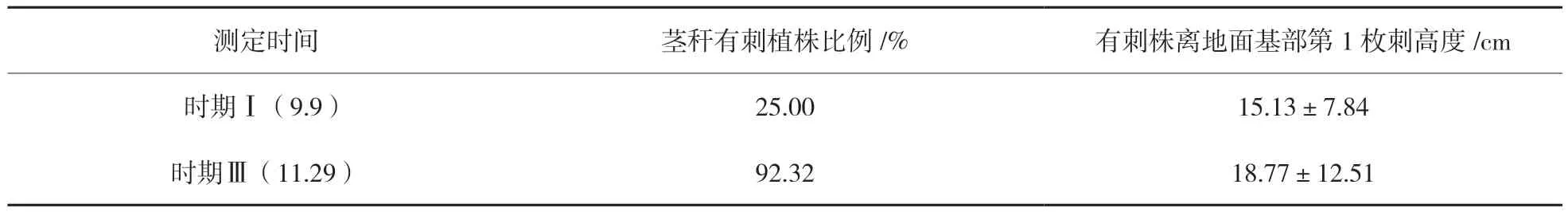
表5 “美賜”1年生苗莖稈皮刺分布變化
由表5可知,在此生長(zhǎng)過(guò)程中“美賜”種苗群體由少數(shù)植株有刺(25.00%)快速生長(zhǎng)成為了絕大部分植株有刺(92.32%)。“美賜”無(wú)刺植株逐漸變得很少,可能是反映了“美賜”控制皮刺生長(zhǎng)性狀的基因表達(dá)是在幼苗的后期階段。苗期出現(xiàn)皮刺可能也是“美賜”植株從幼苗轉(zhuǎn)化為成苗的形態(tài)上變化特征之一。
同時(shí),隨著植株的生長(zhǎng),樹(shù)高增加,植株上皮刺分布的最低位置也發(fā)生一定改變。由表5可知,隨著“美賜”植株1年生后期生長(zhǎng),基部離地面向上出現(xiàn)第1枚刺的位置有上移趨勢(shì),平均分布高度由15.13 cm變?yōu)?8.77 cm。原因可能是由于植株生長(zhǎng)的徑向拉伸導(dǎo)致植株整體伸長(zhǎng)所引起的,如竹子、楊樹(shù)等都有類似情況[26]。
2.3.3.2 “美賜”苗期皮刺數(shù)量分布特性與密度分布特征
“美賜”1年生苗期皮刺數(shù)量分布與密度分布測(cè)定結(jié)果,見(jiàn)表6。

表6 “美賜”1年生苗皮刺分布特性
從表6可知,“美賜”植株基部出現(xiàn)第1枚刺的平均高度為18.77 cm,莖稈上端出現(xiàn)最后1枚刺的高度平均為65.91 cm。皮刺分布的高低位置個(gè)體間差異較大,尤其是莖稈上端分布位置(標(biāo)準(zhǔn)差為16.95 cm)比基部分布位置(標(biāo)準(zhǔn)差為12.51 cm)波動(dòng)更大。
同時(shí),“美賜”植株上的皮刺分布密度不是均勻的,在植株離地面0~60 cm高度內(nèi)集中分布了95.24%的皮刺量。而在分生枝莖稈上的各段分布也不均勻。如圖4所示,在距離分枝起點(diǎn)10 cm以下莖段皮刺分布較少,平均只有0.92枚;在10~20 cm、20~30 cm、30~40 cm的皮刺分布平均值分別為3.75,4.33,2.17枚;在超過(guò)距離分枝起點(diǎn)60 cm以上部分,皮刺分布數(shù)量很少,分布平均值僅0.67枚。
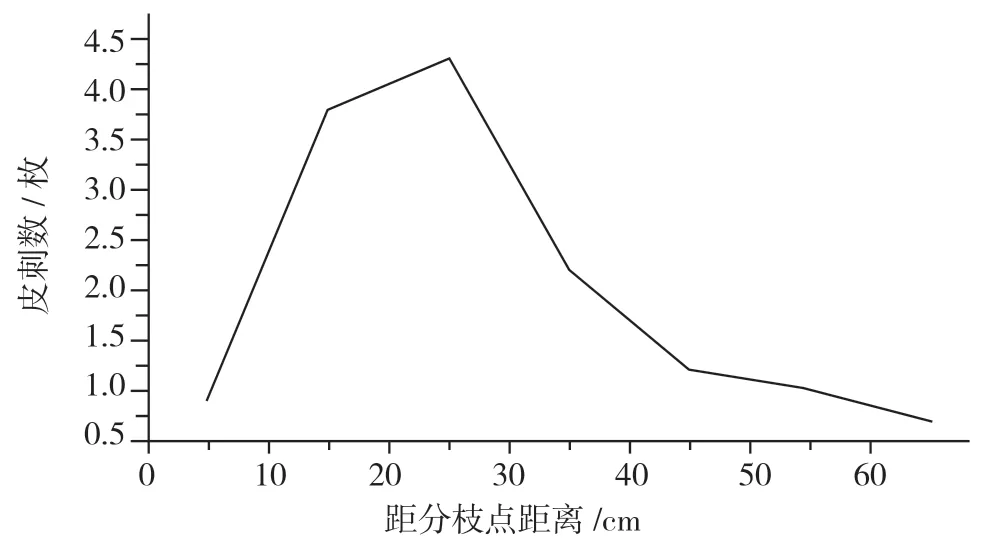
圖4 “美賜”1年生苗距離分枝起點(diǎn)不同高度莖稈上的皮刺數(shù)量
2.4 皮刺性狀的優(yōu)株選擇
對(duì)“美賜”1年生苗群體進(jìn)行皮刺數(shù)量分布進(jìn)行測(cè)定,植株個(gè)體平均皮刺數(shù)量值μ為24.80枚,個(gè)體間分化變異大,標(biāo)準(zhǔn)差σp為14.51枚,以X=μ-σp為入選標(biāo)準(zhǔn),確定皮刺性狀入選標(biāo)準(zhǔn)為數(shù)量≤10.29枚,植株皮刺數(shù)量少于此值為候選少(無(wú))刺優(yōu)株,綜合其他性狀比較,最后確定5個(gè)優(yōu)株,記為優(yōu)-1、優(yōu)-2、優(yōu)-3、優(yōu)-4、優(yōu)-5,最終優(yōu)株入選率為6.41%,各育種參數(shù)如表7所示。通過(guò)選擇,在有性繁殖情況下選擇可得理論遺傳增益?g1為57.10%,在無(wú)性繁殖情況下,通過(guò)選擇可得理論遺傳增益?g2為85.65%,皮刺性狀改良效果將極為明顯。

表7 “美賜”1年生苗期皮刺性狀選擇的育種參數(shù)
5個(gè)優(yōu)株詳細(xì)特征如下:
2.4.1 優(yōu)-1 皮刺數(shù)2,分布于距分枝點(diǎn)20~25 cm,向外著生;莖稈數(shù)2,綠;葉數(shù)17,綠,大小中等,掌狀;無(wú)病蟲(chóng)害。
2.4.2 優(yōu)-2 皮刺無(wú);莖稈數(shù)2,莖1:綠色(夾雜少許紫),莖2:上端綠,基部紫;葉數(shù)26,綠,小,掌狀;無(wú)病蟲(chóng)害。
2.4.3 優(yōu)-3 皮刺無(wú);莖稈數(shù)2,綠(少部分紫);葉數(shù)24,綠,小,掌狀;無(wú)病蟲(chóng)害。
2.4.4 優(yōu)-4 皮刺無(wú);莖稈數(shù)2,莖1:綠(少許紫),莖2:紫(少部分綠);葉數(shù)11,綠,小,掌狀;無(wú)病蟲(chóng)害。
2.4.5 優(yōu)-5 皮刺數(shù)4,分布于距分枝點(diǎn)14~25 cm,向外著生;莖稈數(shù)1,綠;葉數(shù)12,綠,小,掌狀;無(wú)病蟲(chóng)害。
3 討論與結(jié)論
本次試驗(yàn)中,測(cè)定的“美賜”種子千粒重為1.00 g,較普通掌葉覆盆子種子(1.29 g)輕,兩者之間存在顯著性差異。劉宇等[27]在研究種子千粒重方面發(fā)現(xiàn),多倍體家系種子千粒重普遍較重,其平均值較普通二倍體家系重22.42%;同時(shí)多倍體植物由于染色體組增加,器官往往變大,花期延后,而“美賜”萌芽、開(kāi)花均早于普通掌葉覆盆子。由于“美賜”是普通掌葉覆盆子中出現(xiàn)的變異類型,這是否可能表明“美賜”是由于發(fā)生了遺傳變異而遺傳物質(zhì)質(zhì)量變小有待后續(xù)試驗(yàn)驗(yàn)證。
本試驗(yàn)發(fā)現(xiàn),“美賜”1年生苗群體一級(jí)分枝數(shù)多為1枝,二級(jí)分枝數(shù)多為0枝。潘彬榮等[21]也發(fā)現(xiàn)掌葉覆盆子一般當(dāng)年抽發(fā)新枝,不具二級(jí)分枝,第2年抽發(fā)二級(jí)分枝。王碩等[28]認(rèn)為植物激素、遺傳因子對(duì)植株形態(tài)調(diào)控起主要作用,植物激素獨(dú)腳金內(nèi)酯會(huì)影響植株分枝數(shù)。胡盼盼等[29]探究獨(dú)腳金內(nèi)酯對(duì)草莓新莖分枝的調(diào)控機(jī)制時(shí)發(fā)現(xiàn),草莓品種“紅顏”在定植60 d后進(jìn)入分枝盛期、分枝數(shù)平均值為2.5枝,認(rèn)為獨(dú)腳金內(nèi)酯含量降低促進(jìn)草莓分枝,在分枝盛期新莖中積累了較多獨(dú)腳金內(nèi)酯而抑制了分枝數(shù)。美賜種苗分枝數(shù)目較少可能也是由于其體內(nèi)積累較多的獨(dú)腳金內(nèi)酯,而抑制其分枝數(shù)目增加。這有待后續(xù)試驗(yàn)證實(shí),同時(shí)也為美賜植株形態(tài)調(diào)控研究提供了思路借鑒。
試驗(yàn)中發(fā)現(xiàn)“美賜”1年生苗群體中無(wú)刺植株占比為3.80%,這與代漢萍等[30]在樹(shù)莓無(wú)刺性狀雜交試驗(yàn)中“無(wú)刺紅”與“費(fèi)爾圖德”雜交后代出現(xiàn)無(wú)刺實(shí)生苗比率2.5%相近。此外,朱報(bào)著等[31]研究美麗異木棉子代苗木皮刺時(shí),以多皮刺的WY05為母本、少皮刺的XS22無(wú)性系為父本,發(fā)現(xiàn)子代有無(wú)刺植株6株,占6.67%。這些研究結(jié)果進(jìn)一步表明,植株的皮刺性狀存在豐富的遺傳多樣性,掌葉覆盆子無(wú)刺型新品種選育可以通過(guò)更多的雜交組合試驗(yàn),在雜交子代群體中進(jìn)一步選擇優(yōu)株來(lái)實(shí)現(xiàn)。
通過(guò)本試驗(yàn)觀測(cè)“美賜”種子特性、發(fā)芽特性、苗期特性和苗期皮刺選擇,可得到如下結(jié)論:“美賜”種子正常率較高,種子形態(tài)縱徑、橫徑、側(cè)徑和種子千粒重均小于普通種,兩者有顯著性差異。在相同的發(fā)芽環(huán)境條件下,“美賜”種子發(fā)芽率高于普通種,兩者發(fā)芽特性差異明顯。“美賜”1年生種苗生長(zhǎng)性狀株高、冠幅、地徑之間均存在極顯著的正相關(guān)關(guān)系。“美賜”1年生種苗植株皮刺分布主要集中在0~60 cm高度范圍內(nèi)。“美賜”1年生種苗以基生一級(jí)分枝植株為主,一級(jí)分枝起點(diǎn)基本接近地面。對(duì)“美賜”1年生苗皮刺性狀進(jìn)行優(yōu)株再選擇,理論遺傳增益在有性繁殖情況下達(dá)57.10%,在無(wú)性繁殖情況下可達(dá)85.65%,植株少刺改良效果明顯,“美賜”品種優(yōu)良特性可取得進(jìn)一步提升,使更多更優(yōu)的新品種選育獲得豐富的育種材料和育種前景。
