白藜蘆醇通過調(diào)節(jié)鐵死亡通路抑制小鼠潰瘍性結(jié)腸炎相關(guān)性結(jié)腸癌實(shí)驗(yàn)研究
2023-06-21 02:15:24張一楠任彩佩吳亞俐張文琴崔香麗
陜西醫(yī)學(xué)雜志 2023年6期
張一楠,任彩佩,吳亞俐,張文琴,崔香麗
(山西醫(yī)科大學(xué)生理學(xué)系 山西醫(yī)科大學(xué)細(xì)胞生理學(xué)教育部重點(diǎn)實(shí)驗(yàn)室,山西 太原 030001)
炎癥性腸病(Inflammatory bowel disease,IBD)是一種慢性特發(fā)性腸道疾病,包括潰瘍性結(jié)腸炎(Ulcerative colitis,UC)和克羅恩病(Crohn’s disease,CD)。其中,UC的好發(fā)部位是結(jié)腸和直腸,而CD的好發(fā)部位在小腸和結(jié)腸。潰瘍性結(jié)腸炎相關(guān)性結(jié)腸癌(Ulcerative colitis-associated colorectal cancer,UCACC)約占整個(gè)腸癌病死的1%~2%,占UC患者病死的10%~15%,是導(dǎo)致UC患者死亡的主要原因[1]。白藜蘆醇(Resveratrol,Res)是一種從植物中提取的非黃酮類多酚有機(jī)化合物,已被證明有多種生物活性,包括抗炎[2]、抗氧化[3]、抗衰老[4]、抗癌[5]等。前期研究[6]證明白藜蘆醇對結(jié)腸炎和腸炎相關(guān)腸癌有預(yù)防和治療作用,其機(jī)制的闡明對于腸炎和腸癌的治療有臨床意義。鐵死亡是一種新型的細(xì)胞程序性死亡方式,主要特點(diǎn)是細(xì)胞內(nèi)鐵代謝失衡導(dǎo)致的鐵積累和脂質(zhì)過氧化,主要表現(xiàn)為在細(xì)胞內(nèi)鐵的存在下催化細(xì)胞內(nèi)的脂質(zhì)發(fā)生脂質(zhì)過氧化,從而誘導(dǎo)細(xì)胞死亡。發(fā)生鐵死亡時(shí),細(xì)胞膜上高表達(dá)的多不飽和脂肪酸脫氫,觸發(fā)過氧化鏈?zhǔn)椒磻?yīng),產(chǎn)生大量氧自由基,導(dǎo)致細(xì)胞死亡。細(xì)胞內(nèi)鐵超載[7]和脂質(zhì)過氧化[8]是細(xì)胞發(fā)生鐵死亡的先決條件,同時(shí)伴有抗氧化體系胱氨酸谷氨酸轉(zhuǎn)運(yùn)受體(System Xc-)的抑制和谷胱甘肽過氧化物4(Glutathione peroxidase 4,GPX4)的表達(dá)降低。白藜蘆醇抗UCACC的作用與鐵死亡是否有關(guān)鮮有報(bào)道。因此,本實(shí)驗(yàn)探討Res是否可通過調(diào)節(jié)鐵死亡通路抑制UCACC。
1 材料與方法
1.1 實(shí)驗(yàn)動(dòng)物與細(xì)胞 SPF級雄性C57BL/6小鼠28只,6~8周齡,體重21~25 g,購自北京斯貝福生物技術(shù)有限公司。人結(jié)腸癌HCT 116細(xì)胞購自中國科學(xué)院細(xì)胞庫。
1.2 主要試劑 葡聚糖硫酸鈉(DSS,貨號:160110)購自美國MP公司;偶氮甲烷(AOM,貨號:25843-45-2)購自Sigma公司;白藜蘆醇(貨號:501-36-0)和二甲基亞砜(DMSO,貨號:67-68-5)購自Sigma公司;鼠、兔二步法免疫組化檢測試劑盒(貨號:PV-9000)購自北京中杉金橋生物公司;GPX4兔單克隆抗體(貨號:ab125066)、長鏈脂酰輔酶A合成酶4(ASCL4)兔單克隆抗體(貨號:ab155282)、鐵蛋白重鏈(FTH)兔單克隆抗體(貨號:ab183781)、P53小鼠單克隆抗體(貨號:ab26)均購自美國Abcam公司;FTH兔多克隆抗體(貨號:D220253-0025)購自上海生工公司;辣根過氧化物酶(HRP)標(biāo)記山羊抗兔IgG二抗(貨號:HS101)購自北京全式金公司;β-肌動(dòng)蛋白(β-actin)鼠單克隆抗體(貨號:CL594-66009)、HRP標(biāo)記山羊抗鼠IgG二抗(貨號:SA00001-1)購自武漢Proteintech公司;杜爾貝科改良伊格爾高糖培養(yǎng)基(DMEM,貨號:C11965500BT)購自Gibco公司;青霉素鏈霉素雙抗(貨號:V900929)購自Sigma公司。
1.3 小鼠UCACC模型制備 小鼠使用普通飼料適應(yīng)性喂養(yǎng)1周后,隨機(jī)分為四組,即Control組、AOM組、AOM+DSS組、AOM+DSS+Res組,每組7只。造模期間,各組小鼠喂食高鐵飼料。造模第1天,AOM組、AOM+DSS組和AOM+DSS+Res組以10 mg/kg的劑量腹腔注射AOM,Control組等劑量注射0.9%氯化鈉溶液。第1周各組小鼠均飲用滅菌水;第2周開始AOM+DSS組和AOM+DSS+Res組飲用含1% DSS水;第3周所有組飲用滅菌水;第4周AOM+DSS組和AOM+DSS+Res組飲用含1% DSS水,其余組飲用滅菌水。以此類推,直到70 d造模周期結(jié)束。造模第2周開始,AOM+DSS+Res組給予0.1 ml 12 mg/ml的Res灌胃,每周灌胃5次。70 d造模結(jié)束后,安樂死小鼠,取出小鼠結(jié)腸,部分結(jié)腸用于提取蛋白,另一部分用4%多聚甲醛固定結(jié)腸24 h后對結(jié)腸組織進(jìn)行包埋,切片。
1.4 細(xì)胞培養(yǎng) 使用高糖DMEM+10%胎牛血清+1%青霉素鏈霉素雙抗配置培養(yǎng)基,于37 ℃、5% CO2細(xì)胞培養(yǎng)箱中培養(yǎng)細(xì)胞。細(xì)胞傳至3~4代后,按8×105個(gè)/孔細(xì)胞接種于6孔板。接種后的細(xì)胞放入培養(yǎng)箱中培養(yǎng),24 h后分別在每孔中加入2 μl濃度為4、8、16 mg/ml的白藜蘆醇,空白對照組加入2 μl DMSO,藥物終濃度調(diào)整為0、4、8、16 μg/ml。培養(yǎng)箱中孵育48 h,收集細(xì)胞進(jìn)行下一步實(shí)驗(yàn)。
1.5 小鼠結(jié)腸組織HE染色 染色之前將小鼠結(jié)腸組織石蠟切片放于烘箱中,60 ℃烤片1 h。脫蠟復(fù)水,切片依次浸泡于二甲苯、不同濃度的無水酒精中,蒸餾水沖洗。切片上滴加蘇木素染液染色,蒸餾水沖洗,伊紅染色,去離子水沖洗。染色后的切片依次經(jīng)過不同濃度的無水乙醇和二甲苯,切片脫水后晾干,中性樹脂封片后顯微鏡下觀察并拍照,記錄小鼠結(jié)腸組織的形態(tài)學(xué)改變。
1.6 免疫組織化學(xué)(IHC)檢測小鼠結(jié)腸組織FTH、ACSL4、GPX4、P53蛋白的表達(dá) 石蠟切片60 ℃烤片1 h。脫蠟復(fù)水,依次將切片放入二甲苯和不同濃度的乙醇中脫蠟復(fù)水,磷酸鹽緩沖液沖洗。之后把切片放入抗原修復(fù)液中,待水浴鍋溫度升高到98 ℃之后將切片放入,于98 ℃進(jìn)行抗原修復(fù)。抗原修復(fù)結(jié)束后,待切片冷卻至室溫,PBS沖洗。避光,用免疫組化筆圈住組織,滴加一滴內(nèi)源性過氧化酶阻斷劑(封閉液),去除內(nèi)源性過氧化物酶。封閉結(jié)束后,PBS沖洗,分別在組織上滴加FTH、GPX4、ACSL4、P53一抗,一抗完全覆蓋組織。切片放于濕盒中4 ℃過夜。第2天,取出濕盒復(fù)溫,復(fù)溫結(jié)束后避光滴加二抗反應(yīng)增強(qiáng)液,反應(yīng)結(jié)束后PBS沖洗。滴加酶標(biāo)山羊抗兔/小鼠IgG聚合物,二抗孵育結(jié)束后PBS沖洗。DAB染液顯色,顯色結(jié)束后PBS沖洗,之后蘇木素復(fù)染、分化液分化。切片依次浸泡于不同濃度的乙醇和二甲苯中,脫水透明,待切片自然晾干后中性樹脂封片,顯微鏡下觀察并拍照,用Image J軟件分析結(jié)果。
1.7 CCK-8實(shí)驗(yàn)檢測白藜蘆醇抑制HCT 116細(xì)胞增殖 將HCT 116細(xì)胞分為空白對照組及4、8、16 μg/ml白藜蘆醇組。當(dāng)細(xì)胞生長至對數(shù)生長期后,接種于96孔板上,孵育24 h貼壁后采用4、8、16 μg/ml白藜蘆醇處理HCT 116細(xì)胞,空白對照組加入等量DMSO。48 h后,棄去原培養(yǎng)基。加入含10% CCK-8檢測液的無血清無雙抗培養(yǎng)基,2 h后酶標(biāo)儀于450 nm波長處檢測吸光度值。
1.8 蛋白質(zhì)印跡(Western blot)檢測小鼠結(jié)腸組織和人HCT 116細(xì)胞FTH、ACSL4、GPX4、P53蛋白的表達(dá) 取適量小鼠結(jié)腸組織或收集的HCT 116細(xì)胞,加入裂解液(增強(qiáng)型RIPA+PMSF),超聲1 min破碎細(xì)胞,離心取上清,BCA法測蛋白濃度。測量完畢后,以蛋白質(zhì)終濃度為2 mg/ml配置上樣體系,在上樣體系中加入RIPA、上樣緩沖液、蛋白質(zhì)上清。配置好后100 ℃煮蛋白15 min使蛋白變性。煮蛋白結(jié)束后冷卻至室溫,進(jìn)行SDS-PAGE凝膠電泳。每孔上樣量10 μl,電泳條件為100 V、25 min/150 V、50 min。電泳結(jié)束后使用半干轉(zhuǎn)將蛋白由凝膠上轉(zhuǎn)移到PVDF膜上。轉(zhuǎn)膜結(jié)束后,洗膜5次,每次7 min。之后用封閉液封閉2 h,洗膜。之后分別加入FTH、ACSL4、GPX4、P53、β-actin一抗,搖床4 ℃孵育過夜。第2天TBST洗膜后加入HRP標(biāo)記山羊抗兔IgG或山羊抗小鼠IgG,室溫孵育1 h。二抗孵育結(jié)束后洗膜,滴加發(fā)光液,使用Chemi Doc MP Imaging System系統(tǒng)進(jìn)行顯影,使用Image Lab軟件對條帶進(jìn)行分析。
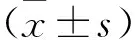
2 結(jié) 果
2.1 小鼠結(jié)腸組織HE染色結(jié)果 見圖 1。與Control組比較,AOM+DSS組小鼠腸壁增厚,上皮細(xì)胞損傷明顯,隱窩結(jié)構(gòu)被破壞,癌細(xì)胞排列紊亂,核大深染,腫瘤細(xì)胞形成類橢圓形結(jié)構(gòu),中央可見空泡。與AOM+DSS組比較,AOM+DSS+Res組小鼠腸壁變薄,未見腫瘤結(jié)構(gòu),僅在黏膜層可見炎細(xì)胞浸潤,腺細(xì)胞排列較為規(guī)則,腸上皮基本完整。HE染色結(jié)果顯示小鼠UCACC模型建模成功,Res可明顯抑制AOM+DSS誘導(dǎo)的小鼠UCACC形成。
2.2 四組小鼠結(jié)腸組織相關(guān)蛋白IHC染色結(jié)果比較 見表1(圖2)。與Control組比較,AOM組FTH蛋白表達(dá)下降;AOM+DSS組GPX4和FTH蛋白表達(dá)明顯上升,ACSL4和P53蛋白表達(dá)明顯下降(均P<0.05)。與AOM+DSS組比較,AOM+DSS+Res組GPX4和FTH蛋白表達(dá)下降,ACSL4和P53蛋白表達(dá)明顯上升(均P<0.05)。
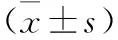
表1 四組小鼠結(jié)腸組織相關(guān)蛋白IHC染色結(jié)果比較

圖1 四組小鼠結(jié)腸組織HE染色結(jié)果(左×50,中×100,右×200)

圖2 四組小鼠結(jié)腸組織GPX4、ACSL4、FTH、P53蛋白IHC染色結(jié)果
2.3 四組小鼠結(jié)腸組織相關(guān)蛋白Western blot檢測結(jié)果比較 見表2。與Control組比較,AOM組GPX4、FTH蛋白表達(dá)下降,ACSL4表達(dá)上升;AOM+DSS組GPX4和FTH蛋白表達(dá)上升,ACSL4和P53蛋白表達(dá)下降(均P<0.05)。與AOM+DSS組比較,AOM+DSS+Res組GPX4和FTH蛋白表達(dá)下降,ACSL4和P53蛋白表達(dá)上升(均P<0.05)。
表2 四組小鼠結(jié)腸組織相關(guān)蛋白Western blot檢測結(jié)果比較
2.4 四組HCT 116細(xì)胞存活率比較 CCK-8實(shí)驗(yàn)結(jié)果顯示,與空白對照組比較,用Res處理HCT 116細(xì)胞后,鏡下觀察可見細(xì)胞數(shù)量減少,細(xì)胞不貼壁,漂浮在培養(yǎng)基上。空白對照組及4、8、16 μg/ml Res組細(xì)胞存活率依次為0.907±0.060、0.861±0.014、0.813±0.048和0.720±0.095,8、16 μg/ml Res組細(xì)胞存活率低于空白對照組(均P<0.05)。
2.5 四組HCT 116細(xì)胞GPX4、ACSL4、FTH蛋白表達(dá)水平比較 見表3。Western blot檢測結(jié)果顯示,與空白對照組比較,4、8、16 μg/ml Res組細(xì)胞GPX4和FTH蛋白表達(dá)下降,ACSL4蛋白表達(dá)上升(均P<0.05)。
表3 四組HCT 116細(xì)胞GPX4、ACSL4、FTH蛋白表達(dá)水平比較
3 討 論
Res是最初從毛葉藜蘆中分離出來的一種酚類物質(zhì),大量存在于葡萄、葡萄酒、花生、大豆和漿果中,是一種植物在緊急狀況下分泌出的抗毒素[9]。大量實(shí)驗(yàn)證明Res可以調(diào)控多種信號通路,包括凋亡、壞死、焦亡、細(xì)胞周期、氧化應(yīng)激、炎癥、脂代謝[10]。
結(jié)腸癌是一種發(fā)生部位在結(jié)腸的惡性腫瘤,已經(jīng)成為病死率第三高的癌癥[11]。結(jié)腸癌的治療手段包括手術(shù)切除、放療、聯(lián)合化療、免疫治療、生物靶向治療等。有實(shí)驗(yàn)[6,12-13]證實(shí),Res可以抑制結(jié)腸癌的發(fā)生。本次實(shí)驗(yàn)結(jié)果表明,Res作用于鐵死亡相關(guān)通路蛋白,升高了鐵死亡相關(guān)蛋白ACSL4、P53的表達(dá),降低了GPX4、FTH蛋白的表達(dá),證明Res可以通過促進(jìn)鐵死亡預(yù)防和治療結(jié)腸癌。
鐵死亡是一種鐵依賴的非凋亡細(xì)胞死亡形式,是由于在細(xì)胞內(nèi)鐵的存在下催化產(chǎn)生了大量的活性氧(ROS)加速脂質(zhì)過氧化導(dǎo)致細(xì)胞死亡的過程[14]。機(jī)體內(nèi)的鐵(Fe3+)與轉(zhuǎn)鐵蛋白(TF)形成復(fù)合物后與轉(zhuǎn)鐵蛋白受體(TFR)結(jié)合運(yùn)送至細(xì)胞內(nèi)還原成Fe2+,維持正常細(xì)胞內(nèi)Fe2+含量動(dòng)態(tài)平衡。而細(xì)胞內(nèi)游離Fe2+含量升高可以通過芬頓反應(yīng)產(chǎn)生大量ROS損傷細(xì)胞[15]。鐵死亡也在腸道疾病的發(fā)病中起重要作用,研究[16]證實(shí)鐵攝入超標(biāo)會增加患癌風(fēng)險(xiǎn)。鐵死亡對腸道的作用是雙向的,與結(jié)腸癌的增殖和侵襲有關(guān)。在小鼠結(jié)腸組織缺血-再灌注損傷時(shí),小鼠結(jié)腸組織鐵死亡促進(jìn)因子ACSL4含量上升,抑制因子GPX4、FTH表達(dá)水平降低,而且通過藥物抑制鐵死亡可以減輕結(jié)腸組織上皮的損傷,避免結(jié)腸癌的發(fā)生[17]。還有報(bào)道[16]稱,可以通過在結(jié)腸癌細(xì)胞中誘導(dǎo)鐵代謝失衡和細(xì)胞脂質(zhì)過氧化促進(jìn)鐵死亡,從而抑制結(jié)腸癌。
ACSL4、GPX4、FTH是鐵死亡的標(biāo)記蛋白。ACSL4是體內(nèi)脂肪酸代謝的關(guān)鍵酶,主要功能是催化長鏈脂肪酸形成長鏈酰基輔酶A,參與磷脂酰乙醇胺或磷脂酰肌醇等帶負(fù)電膜磷脂的合成[18-19],膜磷脂過氧化是誘發(fā)細(xì)胞鐵死亡的關(guān)鍵步驟。GPX4是細(xì)胞內(nèi)重要的抗氧化酶[20],在谷胱甘肽存在的情況下催化降解細(xì)胞內(nèi)脂質(zhì)過氧化物而使其失去氧化活性,細(xì)胞內(nèi)GPX4上升可以抑制鐵死亡。FTH可以結(jié)合多余的游離Fe2+維持細(xì)胞內(nèi)鐵穩(wěn)態(tài),溶酶體攝取并降解FTH釋放出大量的游離鐵促進(jìn)鐵死亡的發(fā)生,發(fā)生鐵死亡時(shí)FTH的表達(dá)下降。P53是經(jīng)典的抑癌基因,參與細(xì)胞凋亡、細(xì)胞周期調(diào)節(jié)、DNA錯(cuò)配修復(fù)和抑制腫瘤血管生成等過程,有實(shí)驗(yàn)證明P53通過抑制細(xì)胞內(nèi)System Xc-的作用抑制細(xì)胞對胱氨酸的攝取,降低GPX4的活性,增強(qiáng)細(xì)胞對鐵死亡的敏感性。本實(shí)驗(yàn)結(jié)果顯示,UCACC模型小鼠結(jié)腸組織和人結(jié)腸癌HCT 116細(xì)胞中ACSL4、P53蛋白表達(dá)下降,GPX4、FTH蛋白表達(dá)上升,用Res處理的小鼠和細(xì)胞中ACSL4、P53蛋白表達(dá)上升,GPX4、FTH蛋白表達(dá)下降,證明Res可以通過促進(jìn)鐵死亡抑制小鼠UCACC。
綜上所述,Res可以通過調(diào)控細(xì)胞鐵死亡通路抑制UCACC,為預(yù)防和治療UCACC的機(jī)制提供了實(shí)驗(yàn)依據(jù)。下一步將探討Res影響UCACC細(xì)胞鐵死亡的靶點(diǎn),為UCACC治療機(jī)制提供更詳細(xì)的資料。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
西南國防醫(yī)藥(2016年7期)2016-12-01 06:01:15
中國衛(wèi)生標(biāo)準(zhǔn)管理(2015年1期)2016-01-14 03:41:26
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
河南醫(yī)學(xué)研究(2014年3期)2014-02-27 14:51:48
沈陽醫(yī)學(xué)院學(xué)報(bào)(2014年1期)2014-02-16 06:19:24
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
