桂花OfMYB1R47轉錄因子在芳香揮發物形成過程中的功能分析
2023-06-07 06:39:52岳遠征胡宏敏劉家偉申慧敏施婷婷楊秀蓮王良桂
浙江農林大學學報 2023年3期
岳遠征,胡宏敏,劉家偉,申慧敏,施婷婷,楊秀蓮,王良桂
(1.南京林業大學 風景園林學院,江蘇 南京 210037;2.南京林業大學 江蘇省風景園林重點實驗室,江蘇 南京 210037)
桂花Osmanthusfragrans是木犀科Oleaceae 木犀屬Osmanthus常綠木本植物,具有花朵形小繁多、花香迷人的特點,是一種應用廣泛的優良園林樹種[1-2],并在食品和香水等商品中具有較高的應用價值[3]。有研究表明:桂花花香揮發物依據化學結構可劃分為6 個類別,即萜烯類、酯類、烷烴類、酮類、醇類和醛類[4-6]。也有研究通過氣相色譜-嗅覺測量法(GC-Olfactometry)分析,共鑒定出11 種對桂花香氣形成具有貢獻的萜類活性物質[7];同時,在‘日香桂’O.fragrans‘Rixianggui’中也發現芳樟醇的含量與花香強度的變化高度相關[8];此外,β-紫羅蘭酮、芳樟醇及其衍生物等關鍵花香物質在不同桂花品種中相對含量的改變還是導致桂花品種間花香感官品質差異的重要原因[9]。
花香物質合成的轉錄調控是一個復雜的過程,其中轉錄因子對植物次生代謝物合成途徑中結構基因的轉錄調控被認為是植物揮發性芳香物質生物合成的重要環節[10-11]。在桂花中,轉錄因子OfWRKY3 和OfERF61 可以直接調控催化產生揮發性芳香物紫羅蘭酮的類胡蘿卜素裂解雙加氧酶4 基因(OfCCD4)的表達,并認為它們可能參與了調節酮類花香物質紫羅蘭酮的合成[12-13]。此外,有研究通過氣相色譜-質譜聯用法(GC-MS)分析桂花花香揮發物,并結合桂花全基因組WRKY 和MYB 轉錄因子的鑒定,獲得了一批與桂花萜烯類花香物質合成相關的候選基因[3,14]。
根據MYB 結構域的數量,MYB 轉錄因子分為4 個亞家族:1R-MYB、R2R3-MYB、3R(R1R2R3)-MYB 和4R-MYB 蛋白[15]。其中,R2R3-MYB 已被證實與多種生物過程有關,其在調控揮發性香氣物質的合成與釋放以及增強非生物脅迫能力等方面均具有重要的作用[15-18]。1R-MYB 蛋白統稱為MYBrelated 蛋白,是最大的MYB 基因亞家族,在某些物種中MYB-related 成員的數量甚至是R2R3-MYB 的2 倍[18-19]。目前,在R2R3-MYB 亞家族中已鑒定出多個與花香揮發物合成有關的成員,如在花香模式植物矮牽牛Petuniahybrida中已鑒定出4 個通過調控苯基/苯丙烷代謝途徑中結構基因的表達,來參與花香物質合成的R2R3 型MYB 轉錄因子[20-21];此外,在玫瑰Rosarugosa[22]、留蘭香Menthaspicata[23-24]、姜花Hedychiumcoronarium[25]、百合Liliumbrowniivar.viridulum[26]等觀賞植物中,R2R3-MYB 轉錄因子也會參與調控植物花香物質的合成,但關于MYB-related 亞家族成員在調節植物花香合成的研究仍鮮有報道。
基于前期桂花全基因組和轉錄組的測序工作[27],本研究篩選出了1 個與花香釋放規律較為一致的MYB-related 轉錄因子,并根據其在染色體上的位置命名為OfMYB1R47。本研究擬先通過實時熒光定量PCR (RT-qPCR)技術,分析OfMYB1R47 基因在‘日香桂’不同發育時期中的表達特性;并結合亞細胞定位和酵母自激活實驗進行基因特性分析;然后將該基因在本氏煙草Nicotianabenthamiana中瞬時表達;最后通過GC-MS 分析檢測瞬時轉化植株與瞬時轉化空載植株中主要揮發性有機物質質量分數的變化,進而探討該基因在桂花花香合成過程中的功能,旨在揭示桂花花香合成的轉錄調控機制。
1 材料與方法
1.1 材料
2021年10月,采集南京林業大學校園內‘日香桂’5 個不同發育階段的花朵,分別為鈴梗期(S1,聚傘花序開始分散延伸),香眼期(S2,花被紋理明顯,花頂端出現孔隙),初花期(S3,大部分花處于半閉合狀態,花梗挺立),盛花期(S4,花大部分或近全部呈開放狀態)和末花期(S5,花全部開放,花瓣有所褶皺)[28]。所有樣品采集后立即放入液氮中速凍處理,并置于-80 ℃超低溫冰箱保存。每個花期具有3 個生物學重復。此外,還采集了瞬時轉化OfMYB1R47 基因和空載體的本氏煙草葉片,用于后續目的基因半定量表達分析。本氏煙草種子由南京林業大學風景園林學院保存。
1.2 桂花OfMYB1R47 轉錄因子的克隆及進化分析
使用RNAprep Pure Plant 試劑盒[ 天根生化科技(北京) 有限公司] 從桂花5 個花期提取桂花的總RNA。隨后從總RNA 中去除gDNA 后,用SuperMix 反轉錄試劑盒[天根生化科技(北京)有限公司]合成cDNA,條件如下:65 ℃ 5 min,冰浴2 min,42 ℃ 30 min,最后85 ℃ 5 s[29]。將cDNA 用去離子水稀釋10 倍,用于后續的基因克隆。使用 Primer Premier 5.0 軟件設計克隆OfMYB1R47 轉錄因子編碼區的引物(表1),基因克隆的反應體系為:cDNA 1 μL、上下游引物各1 μL、PrimeSTAR mix 10 μL,ddH2O 補至20 μL。PCR 擴增程序為:98 ℃ 10 s;58 ℃ 30 s,72 ℃ 90 s,35 個循環;72 ℃ 10 min[30]。通過凝膠電泳檢測后,使用北京全式金生物技術公司的DNA 切膠回收試劑盒對擴增產物進行收集,再將目標片段連接至用SmaI和KpnI酶切過的Super1300 載體上,并通過熱激法轉化大腸埃希菌Escherichiacoli,隨后挑選陽性菌落送至測序公司測序,最后通過凍融法將測序正確的質粒轉化根癌農桿菌Agrobacterium tumefaciens。此外,使用美國生物技術信息中心(NCBI)在線網址(https://blast.ncbi.nlm.nih.gov/Blast.cgi)得到了與OfMYB1R47 序列同源性較高的物種中的基因序列,并用MEGA 7.0 軟件,采用鄰接法(Neighbor-joining)分析1 000 次來構建系統進化樹。

表1 本研究所使用引物Table 1 Primers used in this study
1.3 桂花OfMYB1R47 轉錄因子的RT-qPCR 分析以及瞬時轉化植株的半定量表達分析
按照上述方法提取了桂花5 個花期的總RNA,并將其反轉錄為cDNA,用于基因表達分析。使用Primer Premier 5.0 設計OfMYB1R47 轉錄因子的RT-qPCR 引物。并且選擇桂花OfACTIN作為內參基因[31]。每個RT-qPCR 的表達量均來源于3 個生物學重復和3 個技術重復。原始數據使用SPSS 26.0 中的Turkey 檢驗進行差異顯著性分析,利用2-ΔΔCt方法進行表達量分析。同時,提取瞬時轉化OfMYB1R47 基因和空載體植株葉片中的總RNA,并反轉錄成cDNA 進行目的基因OfMYB1R47 的半定量表達分析[14]。煙草NbL25 基因被選為內參基因。此外,還選取了與桂花OfCCD4 和OfCCD1 同源的基因NbCCD4 和NbCCD1 來探究OfMYB1R47 在煙草揮發性香氣物質合成過程中潛在的調控關系。
1.4 桂花OfMYB1R47 轉錄因子的亞細胞定位及酵母自激活
使用不含終止密碼子,但包含了OfMYB1R47 轉錄因子蛋白質編碼區(CDS)的Super 1300 載體以及SmaI和KpnI限制位點構建了35S∷OfMYB1R47∷GFP。然后將含有35S∷OfMYB1R47∷GFP的根癌農桿菌(GV3101)和陰性對照載體(EV)注射到生長了35 d 的本氏煙草葉片中進行亞細胞定位分析。注射所用的煙草植株生長在溫度為(26±2) ℃、光強為144 μmol·m-2·s-1、光照為15 h 光/9 h 暗的生長室中。注射完載體的煙草澆透水后放置在生長室中生長2 d 后,滴入稀釋100 倍后的4,6-二脒基-2-苯基吲哚(DAPI)染液,再放置在LSM710 激光共聚焦顯微鏡下觀測綠色熒光信號。
此外,利用EcoRI和SmaI限制性酶切位點,將缺失終止密碼子的OfMYB1R47 基因的CDS 區域連接到pGBKT7 載體上。然后使用酵母菌株AH109 (上海唯地)獲得pGBKT7-OfMMB1R47 和pGBKT7 載體。最后經PCR 陽性檢測后,挑取陽性克隆的單菌落在選擇性培養基SD/-Trp、SD/-Trp-Ade 和SD/-Trp-Ade+X-α-gal 中黑暗培養3 d,在恒溫培養箱(30 ℃)中觀察目的基因是否具有自激活活性。
1.5 桂花OfMYB1R47 轉錄因子在本氏煙草葉片中的瞬時表達及芳香成分分析
聯合頂空固相微萃取(SPME)和GC-MS 方法檢測瞬時轉化植株與瞬時轉化空載體植株中主要揮發性有機物質量分數的變化。目的基因瞬時轉化方法同1.4。測定葉片中芳香性揮發物的方法如下:每30 mL SPME 瓶底部加入1.5 g 新鮮葉片樣品,中間加入1 μL 稀釋1 萬倍的癸酸乙酯[29]。在65 ℃條件下將蓋瓶中間暴露30 min 后,將萃取頭放到加熱的注射器端口3 min,并在250 ℃下進行解吸附作用。隨后,通過Trace DSQ GC-MS 裝置鑒定解吸后葉片中的芳香揮發物[32],并用正構烷烴混標DLM-1342-5 計算揮發性成分的保留指數(IR),在美國國家標準技術研究所(NIST) 數據庫中(https://webbook.nist.gov/chemistry/cas-ser/),與含有DB-5MS (30.00 m×0.25 mm×0.25 μm)的值進行比較,從而確認花香成分[32-33]。此外,采用以下公式計算芳香揮發物質量分數:各組分質量分數=(各組分峰面積×內標質量/內標峰面積/樣品質量)。在SIMCA 14.1 軟件中,采用主成分分析法(PCA)對數據組內的重復性和組間的差異性進行分析,借助正交偏最小二乘判別分析法(OPLS-DA)和變量重要性投影值(VIP)對轉基因組和對照組間關鍵的有機揮發物進行鑒定。
2 結果與分析
2.1 桂花OfMYB1R47 轉錄因子的克隆及編碼蛋白分析
以桂花‘日香桂’盛花期花朵的cDNA 為模板,使用特異性引物擴增獲得了長度為1 485 bp,可編碼494 個氨基酸的OfMYB1R47 基因序列。進一步分析發現:OfMYB1R47 基因具有MYB-related 基因家族的保守結構域(圖1)。將從NCBI 在線網址中獲得的13 個物種中與桂花同源性較高的目的基因進行系統發育樹的構建,結果表明:OfMYB1R47 與木犀科木樨欖Oleaeuropaeasubsp.europaea中的基因(CAA3020406.1)具有最高的同源性(圖2)。

圖1 桂花OfMYB1R47 轉錄因子編碼的氨基酸序列Figure 1 Amino acid sequence of OfMYB1R47

圖2 OfMYB1R47 蛋白的系統發育樹分析Figure 2 Phylogenetic tree analysis of OfMYB1R47
2.2 RT-qPCR 分析
為了得到可靠的基因表達結果,選擇內參基因ACTIN進行了OfMYB1R47 基因在桂花花朵發育5 個典型花期的表達分析(圖3)。發現OfMYB1R47 基因在5 個花期中的表達展現出與桂花花朵轉錄組數據較為相似的表達趨勢,即OfMYB1R47 的表達在開花期間呈現先上升后下降的模式,在S3 時期達到最高值,S3~S5 時期逐漸下降。

圖3 OfMYB1R47 基因在桂花花朵5 個發育時期的表達模式分析Figure 3 Expression pattern of OfMYB1R 47 in the five flower development stages of O. fragrans
2.3 亞細胞定位及酵母自激活
經過亞細胞定位分析,發現在細胞核和細胞質中均檢測到了35S::OfMYB1R47::GFP融合蛋白的綠色熒光蛋白(GFP) 熒光信號,經DAPI 染色后發現細胞核綠色熒光與DAPI 染色結果重合,證實了OfMYB1R47 轉錄因子主要定位于細胞核(圖4)。除此之外,將該基因的編碼區克隆到酵母表達載體pGBKT7 中,然后將陰性對照pGBKT7 和含有OfMYB1R47 轉錄因子的pGBKT7 載體在連續稀釋的SD/-Trp、SD/-Trp-Ade 和SD/-Trp-Ade+x-α-gal 培養基上培養并進行對比。結果表明:pGBKT7-OfMYB1R47在SD/-Trp 和SD/-Trp-Ade 的平板上能夠正常生長,并在含有x-α-gal 的SD/-Trp-Ade 培養基上變成藍色,這表明OfMYB1R47 轉錄因子具有自激活功能(圖5)。
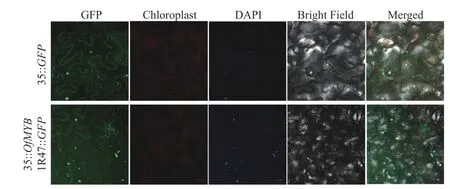
圖4 OfMYB1R47 蛋白在本氏煙草葉片中的亞細胞定位結果Figure 4 Subcellular localization analysis of OfMYB1R47 in the N. benthamiana leaves

圖5 OfMYB1R47 蛋白轉錄自激活分析Figure 5 Transcriptional activation analysis of OfMYB1R47
2.4 瞬時表達植物揮發性香氣成分的驗證與測定
OfMYB1R47 基因的半定量表達分析表明:在瞬時轉化OfMYB1R47 的植株中,都可以檢測到OfMYB1R47 的表達,而在轉化空載體植物中均未檢測到OfMYB1R47 的表達(圖6A),這說明本氏煙草的瞬時轉化體系較為成功,可以用于后續基因功能的分析。從瞬時表達OfMYB1R47 本氏煙草葉片中共鑒定出17 種芳香性揮發物,包括6 種酯類、6 種醛類、2 種酮類等(表2)。通過對瞬時表達OfMYB1R47 基因和空載體植株揮發物的主成分PCA 分析,發現瞬時轉OfMYB1R47 基因的植株可以與瞬時轉化空載體的植株很好地區分開來,表現出較顯著的代謝差異(圖6B)。同時,最小偏二乘法(OPLS-DA)分析的結果也表明瞬時表達OfMYB1R47 和空載體的組分可以完全分離(圖6C)。綜上所述,瞬時超量表達OfMYB1R47 可導致本氏煙草葉片芳香性揮發物質量分數的構成發生改變。

圖6 OfMYB1R47 基因的功能分析Figure 6 Functional analysis of OfMYB1R47

表2 瞬時表達植株中芳香性揮發物的出峰時間和質量分數Table 2 Peak time and content of aromatic volatiles in transient expression plants
基于OPLS-DA 分析,通過篩選VIP>1 和P<0.05 的差異物質,發現與瞬時表達空載體相比,OfMYB1R47 瞬時表達植株中辛醛和β-紫羅蘭酮的相對含量都發生了顯著改變(表2和圖6D)。從GC-MS的峰面積來看,OfMYB1R47 瞬時表達植株中,辛醛的面積明顯比對照瞬時表達植株小,表現為辛醛的相對含量顯著下降;而β-紫羅蘭酮的面積明顯比對照瞬時表達植株大,表現為β-紫羅蘭酮的相對含量明顯上升(圖6E)。此外,RT-qPCR 結果顯示:與瞬時表達空載體植株相比,在瞬時表達OfMYB1R47-2 植株中NbCCD4.1、NbCCD4.2、NbCCD4.3 以及NbCCD1 等與β-紫羅蘭酮合成相關的酶基因的表達量都顯著上升(圖7)。

圖7 本氏煙草CCD4 和CCD1 基因在瞬時表達植株中的表達量Figure 7 Expression of CCD 4 and CCD 1 gene in transiently transformed plants
3 討論與結論
MYB-related 和R2R3-MYB 轉錄因子為植物中MYB 基因家族的2 個主要成員,它們在調節植物生長發育過程和響應非生物脅迫等方面均具有重要的作用。目前,已有較多的與植物揮發性有機化合物合成相關的R2R3-MYB 轉錄因子成員被鑒定出來[24,34-35],然而,MYB-related 成員在花香揮發物合成過程中的功能目前仍鮮有報道。植物MYB-related 轉錄因子通常具有一段高度保守的由51 或52 個氨基酸殘基構成的結構域,該結構域可以用于識別不同類型的順式作用元件并發揮調控作用,這也是MYB-related 基因成員的重要特征[16,36-37]。在OfMYB1R47 轉錄因子的N 端也發現了這段保守結構域,表明該基因為典型的MYB-related 轉錄因子成員。通過系統進化樹分析,在13 個代表性物種中均發現了OfMYB1R47 的直系同源基因,然而這些同源基因的功能仍然未知。
花香調控基因的表達水平往往隨著花朵的發育而表現出顯著的變化,且往往與不同花期花香釋放的強度變化具有較高的相關性[25]。為了獲得較為可靠的基因表達分析結果,本研究使用桂花ACTIN這個內參基因進行RT-qPCR 分析,結果發現:OfMYB1R47 轉錄因子在以內參基因為參考的分析結果中表現為先上升后下降的表達趨勢,這與桂花花香物質釋放的規律一致[38]。在姜花、矮牽牛、留蘭香、蠟梅Chimonanthuspraecox等觀賞植物中研究表明:MYB 轉錄因子家族中的成員可與花香合成酶基因的啟動子結合,通過調控相應靶基因的表達水平來調節植物揮發性有機化合物的合成[20,24-25,39]。在桂花中,也發現核定位轉錄因子OfERF61 可通過調節OfCCD4的表達來影響桂花重要花香物質β-紫羅蘭酮的合成[13]。在本研究中,通過亞細胞定位預測發現:OfMYB1R47 轉錄因子蛋白主要位于細胞核內,為典型的核定位轉錄因子。同時,酵母自激活實驗發現:OfMYB1R47 轉錄因子還具有較強的自激活活性。可見,OfMYB1R47 轉錄因子在桂花花香合成過程中可能具有重要的調控作用。
目前,在本氏煙草中進行基因功能瞬時驗證的體系已經在本源物種轉基因較為困難的植物中廣泛應用[13,25-26]。本研究將OfMYB1R47 轉錄因子在本氏煙草中進行了瞬時超量表達,發現瞬時轉化空載體的植株可與瞬時轉化OfMYB1R47 載體的植株較好地劃分為2 個類群,這說明在本氏煙草中,異源表達OfMYB1R47 基因后會引起本氏煙草芳香性揮發物構成的改變。OPLS-DA 的VIP 值分析表明:辛醛和β-紫羅蘭酮是導致瞬時轉化OfMYB1R47 載體植株與瞬時轉化空載體植株的芳香性揮發物聚類發生明顯區分的重要香味物質,且辛醛和β-紫羅蘭酮的含量與轉化空載體的植株相比都發生了顯著變化,這表明OfMYB1R47 轉錄因子在芳香性揮發物的合成過程中具有重要的調節作用。進一步研究表明:瞬時表達OfMYB1R47 植株中NbCCD4.1、NbCCD4.2、NbCCD4.3 以及NbCCD1 基因的表達明顯上升,表明在本氏煙草中瞬時過表達OfMYB1R47 激活了NbCCD4.1、NbCCD4.2、NbCCD4.3 和NbCCD1 等與β-紫羅蘭酮合成相關的酶基因的表達,從而導致OfMYB1R47 瞬時表達植物中β-紫羅蘭酮相對含量的變化。
本研究鑒定了一個具有轉錄激活活性,在桂花花朵中的表達模式與花香釋放規律一致,且參與調控芳香揮發物合成的核定位轉錄因子OfMYB1R47,有助于加深對MYB-related 轉錄因子亞家族功能的了解,可為今后桂花的花香分子育種提供可靠的基因資源。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
山東工業技術(2016年15期)2016-12-01 05:31:22
當代化工研究(2016年9期)2016-03-20 16:22:08
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
聲屏世界(2014年6期)2014-02-28 15:18:09
終身教育研究(2014年5期)2014-02-28 01:23:06
中國煙草學報(2012年5期)2012-04-12 06:21:18
