重金屬鎘對植物原初光能轉(zhuǎn)化的影響
2023-04-14 03:00:54唐佳樂于志民李潤唐
湖北畜牧獸醫(yī) 2023年1期
唐佳樂,于志民,李潤唐,王 靜
(湛江科技學(xué)院,廣東 湛江 524094)
土壤重金屬污染是世界各國面臨的重大生態(tài)問題,重金屬鎘是中國土壤的首要重金屬污染物。鎘作為一種生命非必需元素,具有極強的生理毒性,能夠通過離子取代、誘導(dǎo)氧化應(yīng)激等途徑破壞信號傳導(dǎo)通路及細胞生物膜結(jié)構(gòu),影響光合作用、呼吸作用等重要生理代謝過程,最終抑制植物的生長與發(fā)育,甚至造成植物死亡。光合作用效率降低是影響植物生長的根本原因,光合作用是光能被生物組織捕獲、儲存、轉(zhuǎn)化的過程,光合能量最終用于驅(qū)動細胞新陳代謝[1]。高等植物的光合作用主要可以分為2 個階段,光反應(yīng)階段和碳反應(yīng)階段,其中光反應(yīng)階段(即原初光能轉(zhuǎn)化過程)主要涉及植物對光合能量的捕獲、傳遞、轉(zhuǎn)化及光合電子傳遞等重要過程。隨著葉綠素?zé)晒鈩恿W(xué)及激光共聚焦熒光壽命成像顯微技術(shù)的發(fā)展,植物光合原初能量轉(zhuǎn)化過程得到了有效的監(jiān)測[2,3]。本研究針對植物光合能量的捕獲、傳遞及轉(zhuǎn)化過程對重金屬鎘的響應(yīng)展開論述,旨在揭示重金屬鎘對葉片光合能量捕獲、傳遞及轉(zhuǎn)化的影響機理,為減緩重金屬鎘的脅迫,提高植物光能吸收能力和光能傳遞效率,促進光能轉(zhuǎn)化提供理論依據(jù)。
1 重金屬鎘對植物葉綠素及類囊體膜結(jié)構(gòu)的影響
1.1 重金屬鎘抑制葉綠素合成、促進葉綠素降解
高等植物中光合色素主要包括葉綠素(Chl a、Chl b)及類胡蘿卜素。葉綠素除捕獲、傳遞光能之外,還在主體捕光色素蛋白復(fù)合體(LHC Ⅱb)三聚體形成過程中具有重要作用[4],少數(shù)特殊Chl a 分子還具有光敏化特性,參與光化學(xué)反應(yīng)(電荷分離)。重金屬鎘對光合色素影響的研究結(jié)果較為一致[5-8],即鎘脅迫下植物葉綠素含量顯著減少,約3周齡的菠菜幼苗經(jīng)90 mg∕kg Cd2+處理10 d 后,葉綠素含量下降68%[9]。即使是較低濃度的鎘脅迫也會導(dǎo)致植物葉綠素含量的降低,燈芯草經(jīng)1 μmol∕L Cd2+處理15 d 后葉綠素含量下降達27.3%[10]。
葉綠素生物合成的受阻是造成葉片內(nèi)葉綠素含量降低的主要原因,高等植物中葉綠素合成最少需要15 個過程并涉及15 種不同的生物合成酶[11],重金屬鎘通過與相關(guān)酶活性位點的巰基(-SH)結(jié)合[12]或下調(diào)葉綠素合成酶相關(guān)基因的表達[8],干擾5-氨基酮戊酸脫水酶(ALAD)、膽紅素原酶(PBGase)、尿卟啉原脫羧酶(UROD)、NADPH 原葉綠素酸酯氧化還原酶(POR)等酶的活性,使葉綠素合成受阻。此外,重金屬鎘引起的缺鐵也會抑制葉綠素的生物合成[13]。
重金屬鎘會通過促進葉綠素的降解以減少葉綠素的含量,通過熒光譜帶觀測到鎘脅迫下菠菜葉片內(nèi)存在大量葉綠素降解產(chǎn)物[9],在另一項研究中也觀察到大量與葉綠素降解途徑相關(guān)的基因(ACD2、NYE1、SGR)的表達量顯著上調(diào)[8]。有研究表明,重金屬鎘對葉綠素中心原子Mg2+具有較強取代作用,僅需30 min 就可將80%的Chl a 轉(zhuǎn)化為缺鎂葉綠素[14],從而降低葉片內(nèi)有效葉綠素的含量。
因此,重金屬鎘通過抑制葉綠素合成、促進葉綠素有降解及離子取代等途徑降低葉片內(nèi)葉綠素含量及有效葉綠素含量,進而影響葉片對光能的捕獲及轉(zhuǎn)化效率。
1.2 重金屬鎘影響葉綠素的組成
葉綠素組成也直接影響光系統(tǒng)Ⅱ捕光色素蛋白復(fù)合體(LHCⅡb)的結(jié)構(gòu)及功能。要形成穩(wěn)定的LHCⅡb 三聚體,Chl a ∕Chl b 只能在一定范圍內(nèi)變化,且必須保證每個三聚體中至少有一個Chl a 分子,Chl b 的合成對LHCⅡb 三聚體的含量也有一定的影響[15]。
不同的植物中Chl a 與Chl b 對重金屬鎘的敏感性有所差異,有研究表明桑樹、辣椒等植物在Pb、Cd脅迫下Chl a ∕Chl b 顯著下降[16,17]。將辣椒幼苗分別置于含0、100 μmol∕L Cd2+的MS 培養(yǎng)基中,Chl a∕Chl b 由3.40 下降為2.87。因此,在這些植物中Chl a對重金屬鎘脅迫可能更為敏感。但有研究提出,Chl b 較Chl a 對重金屬更加敏感,重金屬鎘脅迫下龍葵及茄子Chl a∕Chl b 較非脅迫狀態(tài)下更大[18]。Chl a 與Chl b 對重金屬鎘的敏感性差異可能與脅迫強度及植物種類有關(guān),需要進一步探討。
1.3 重金屬鎘對類囊體膜成分與超微結(jié)構(gòu)的影響
類囊體是由色素、脂類物質(zhì)及蛋白質(zhì)組成的膜系統(tǒng),是葉綠體中執(zhí)行光能吸收與轉(zhuǎn)化的場所。因此,類囊體膜結(jié)構(gòu)及功能的完整性是植物進行光合作用的基礎(chǔ)。研究表明,進入葉綠體的鎘離子有80%積累在類囊體膜中[19],造成類囊體膜結(jié)構(gòu)的解體。
類囊體膜結(jié)構(gòu)的受損與膜脂過氧化有關(guān),各類生物及非生物脅迫通常會導(dǎo)致膜脂過氧化的最終產(chǎn)物丙二醛(MDA)含量的增加。組成類囊體膜的脂類物質(zhì)主要包括磷脂及糖基甘油酯,二者均富含多不飽和脂肪酸[4]。重金屬鎘可以通過脂氧合酶(LOX)介導(dǎo)多不飽和脂肪酸的氧化,產(chǎn)生自由基,進而導(dǎo)致類囊體膜的破壞,如番茄幼苗經(jīng)100 μmol∕L Cd2+脅迫10 d 后,葉片內(nèi)脂氧合酶(LOX)活性顯著增加,同時MGDG(單半乳糖基甘油二脂)、DGDC(雙半乳糖基甘油二脂)及PG(磷脂酰甘油)含量顯著下降,類囊體膜嚴(yán)重受損且基粒解體[20]。
植物PSII 光合放氧中心及LHCⅡb 富含PG,而PG 分子中的反式十六碳-烯酸(trans-16:1)對保持LHCⅡb 寡聚體結(jié)構(gòu)起重要作用[4]。當(dāng)PG 分子中trans-16:1 含量降低時,LHCⅡb 的聚集度下降,LHCⅡb 單體數(shù)量增加,導(dǎo)致LHCⅡb 捕獲的光合能量無法有效地傳遞至反應(yīng)中心,捕光效率下降[21]。而trans-16:1 含量極易受到外界環(huán)境的影響。因此,重金屬鎘是否影響PG 分子中trans-16:1含量及trans-16:1 含量對LHCⅡb 結(jié)構(gòu)的影響機制值得進一步的研究。
2 重金屬鎘對植物PSⅡ光合能量捕獲及傳遞的影響
2.1 PSⅡ光合能量捕獲及傳遞過程
植物PSⅡ光合能量捕獲、傳遞及轉(zhuǎn)化過程如圖1 所示。激發(fā)能從PSⅡ捕光天線系統(tǒng)到反應(yīng)中心存在2 條途徑,①由主體外周捕光天線復(fù)合體(LHCⅡb)中的色素分子吸收光量子,通過輔助外周捕光天線CP26 將激發(fā)能傳遞到內(nèi)周捕光天線CP43 中的色素分子,最后再傳遞至反應(yīng)中心的P680,進行穩(wěn)定的光化學(xué)反應(yīng);②由主體外周捕光天線復(fù)合體(LHCⅡb)中的色素分子吸收光量子,通過輔助外周捕光天線CP29 及CP24 將激發(fā)能傳遞到內(nèi)周捕光天線CP47 中的色素分子,最后再傳遞至反應(yīng)中心的P680,進行穩(wěn)定的光化學(xué)反應(yīng)。
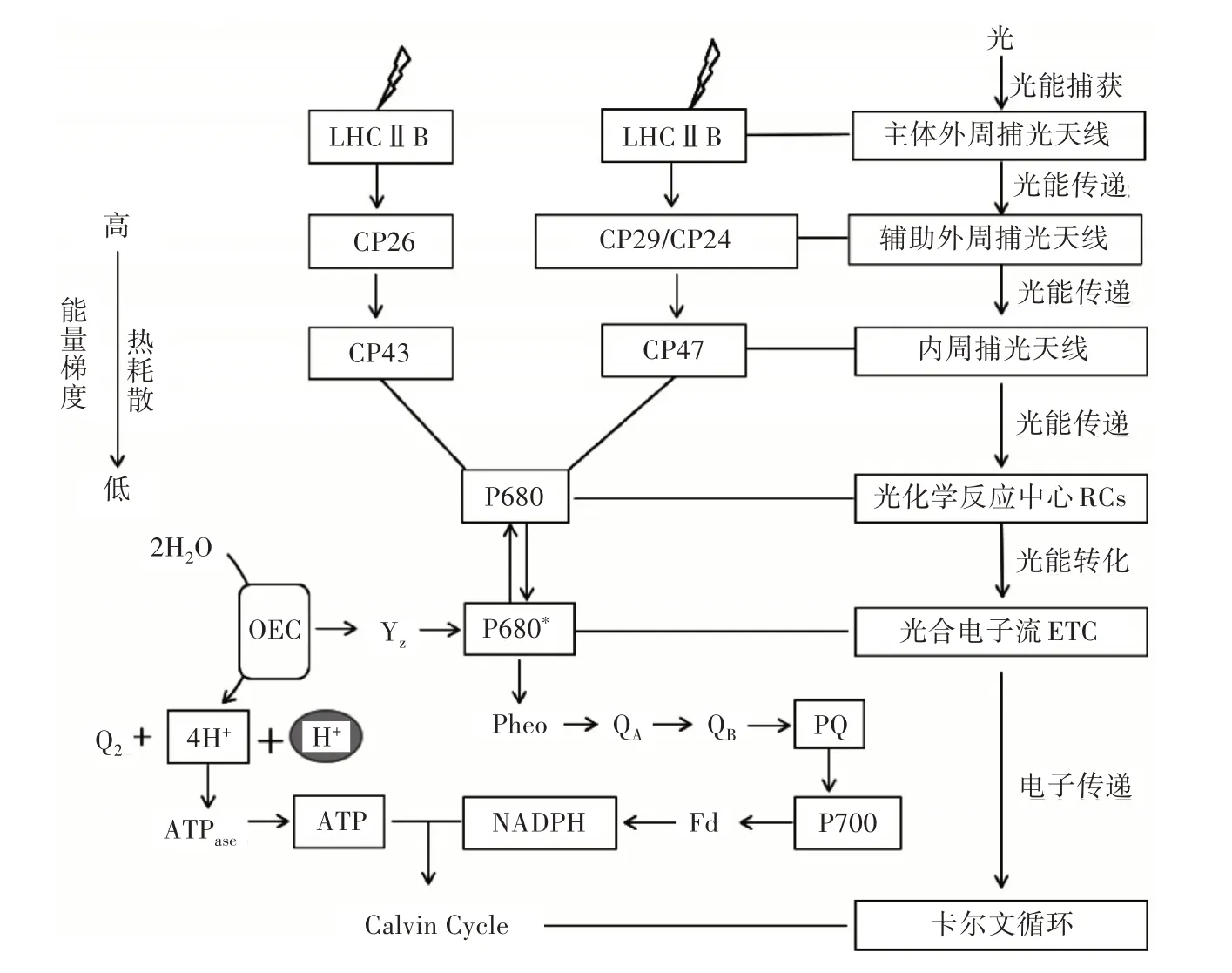
圖1 植物對光能的吸收、捕獲及轉(zhuǎn)化過程
2.2 重金屬鎘對植物PSⅡ光合能量捕獲的影響
2.2.1 植物PSⅡ光合能量捕獲過程對重金屬鎘的響應(yīng) 光合作用始于捕光系統(tǒng)對光能的捕獲,光合能量的捕獲效率直接影響光化學(xué)反應(yīng)效率。重金屬鎘對植物光能捕獲效率的影響尚未得出一致的結(jié)論。
大量研究表明重金屬鎘會促進葉片單位活性反應(yīng)中心(RC)對光能的捕獲,但單位面積葉片捕獲的光能在鎘脅迫下降低。如25 mg∕L Cd2+脅迫下梭魚草葉片單位活性反應(yīng)中心吸收的光能較對照增加32.97%[22],水稻[23]、煙草[24]、番茄[25]等植物中也呈現(xiàn)類似結(jié)果;單位面積葉片捕獲光能在鎘脅迫下降低的原因主要表現(xiàn)為單位活性中心數(shù)量(RC∕CS0)的減少,如雜交狼尾草(Hybrid pennisetum)在鎘脅迫(c(Cd2+)=10~100 mg∕kg)下有18%~23%的光反應(yīng)活性中心轉(zhuǎn)化為非活性中心[26],最終導(dǎo)致單位面積上吸收的光能減少[27]。
有研究表明,雞爪槭處于高濃度鎘脅迫(c(Cd2+)>200 mg∕kg)時,其ABS∕RC 值較對照有所降低[28],而另一項研究中發(fā)現(xiàn)梭魚草葉片在鎘脅迫下ABS∕CSm較對照增加28.53%,但單位面積熱耗散的能量(DI0∕CS0)也響應(yīng)增加了80.16%[22],這可能是不同的研究材料所處環(huán)境差異所致。
2.2.2 重金屬鎘對植物PS Ⅱ光合能量捕獲的影響機制 主體外周捕光天線系統(tǒng)(LHCⅡb)結(jié)合葉片中約45%的葉綠素,起主要的光能捕獲功能[4]。因此,重金屬鎘對植物光能捕獲效率的影響在本質(zhì)上是重金屬鎘對LHCⅡb 結(jié)構(gòu)及功能的影響。
在植物體內(nèi)LHCⅡb 主要以三聚體的形式存在,也有單體和寡聚體形式的LHCⅡb存在,LHCⅡb寡聚體和單體中不但色素吸收的光能減少,且從類胡羅卜素到Chl b 及從Chl b 到Chl a 的能量傳遞效率均低于三聚體[4]。因此,LHCⅡb 三聚體結(jié)構(gòu)的形成是高效光能捕獲的前提。
重金屬鎘會降低LHCⅡb 的聚集度[29]。重金屬鎘可能通過降低磷脂酰甘油PG 及反式十六碳-烯酸(trans-16:1)的含量影響LHCⅡb 的聚集度,進而降低LHCⅡb 對光能的捕獲效率。
光合色素對LHCⅡb 的聚集度也具有一定的影響,葉綠素b 在維持LHCIIb 結(jié)構(gòu)穩(wěn)定中具重要作用[30,31]。擬南芥中Chl a ∕Chl b 由2.85 下降為2.65時,其外周捕光色素蛋白復(fù)合體含量增加20%[15],而在缺失Chl b 的萊茵衣藻突變體中,PSⅡ天線的葉綠素從320 分子降到了95 分子[32]。此外,要形成穩(wěn)定的LHCⅡb 三聚體,至少要保證每個三聚體中有一個Chl a[33]。因此,重金屬鎘可以通過影響葉綠素的含量及組成來影響LHCⅡb 的結(jié)構(gòu),進而影響LHCⅡb 對光能的捕獲效率。
此外,重金屬鎘可以通過影響LHCⅡb 狀態(tài)的轉(zhuǎn)變來影響LHCⅡb 對光能的捕獲效率。當(dāng)植物處于脅迫條件下時,捕光系統(tǒng)捕獲的光能無法被及時消耗,植物葉片可能通過LHCⅡ的磷酸化引起的狀態(tài)轉(zhuǎn)換及LHCⅡ的可逆脫離降低PSⅡ捕光系統(tǒng)有效吸收截面,從而抑制捕光復(fù)合體對光能的吸收[34]。
在鎘脅迫下植物葉片單位活性中心對光能捕獲的增加的同時常觀察到LHC Ⅱb 表觀尺寸的增大[35]。在PS Ⅱ-LHC Ⅱ超級復(fù)合體周圍存在一定數(shù)量的含Chl a 較少的輔助捕光色素蛋白單體(CP29、CP26、CP24)[36]及部分沒有結(jié)合或松散結(jié)合的LHCⅡb 三聚體,當(dāng)植物處于逆境脅迫時,葉綠素含量的降低及光反應(yīng)活性中心的部分失活導(dǎo)致葉片對光能的捕獲能力下降,PS Ⅱ-LHC Ⅱ超級復(fù)合體將結(jié)合更多的單體捕光色素蛋白及游離的LHCⅡ三聚體,以增強葉片的捕光能力[37]。
重金屬鎘會通過改變LHCⅡb 結(jié)構(gòu)及功能來降低植物葉片光能捕獲效率,而單個LHCⅡb 對鎘脅迫的響應(yīng)存在差異,可能與鎘脅迫程度及植物種類有關(guān)。
2.3 重金屬鎘對植物PS Ⅱ光合能量傳遞的影響
主體外周捕光復(fù)合體LHC Ⅱb 捕獲光量子后通過微量LHCⅡ(CP29、CP26、CP24)將激發(fā)能傳遞給內(nèi)周捕光復(fù)合體(CP43、CP47),而內(nèi)周捕光復(fù)合體將進一步激發(fā)能傳遞到反應(yīng)中心D1∕D1 異二聚體上引起電荷分離(P680→P680*),因此,光合能量的傳遞效率直接影響著光化學(xué)反應(yīng)的正常進行。
重金屬鎘會抑制光合能量向PSⅡ反應(yīng)中心的傳遞。在正常生理狀態(tài)下,植物的最大光化學(xué)效率Fv∕Fm較為穩(wěn)定,一般為0.80~0.85[37],而在干旱、高溫、高鹽、強光等脅迫下常表現(xiàn)一定程度的下降,鎘脅迫下水稻[38,39]、辣椒[40]、龍葵[18]、芥菜[41]等植物中均發(fā)現(xiàn)Fv∕Fm隨鎘脅迫程度加深而降低,其中不乏一些鎘超富集植物(龍葵及芥菜)。由此,PSⅡ反應(yīng)中心對激發(fā)能的捕獲效率較易受到鎘脅迫的抑制。此外,有研究表明鎘脅迫下光合能量分配到光化學(xué)反應(yīng)中心的比例(TRo)降低,而熱耗散比例(DIo)顯著提高[39,42],即鎘脅迫抑制了光合能量的傳遞過程,導(dǎo)致外周捕光復(fù)合體捕獲的光合能量無法正常傳遞到光化學(xué)反應(yīng)中心[39],使更多的激發(fā)能在天線系統(tǒng)中熱耗散的形式消耗,以減少過剩激發(fā)能對光系統(tǒng)的傷害[39,42]。
重金屬鎘抑制光合能量傳遞的機制尚不清楚,理論上天線色素吸收的光子有95%~99%的能量可以傳遞至反應(yīng)中心,而天線色素間的能量傳遞屬于物理現(xiàn)象(熒光共振能量轉(zhuǎn)移(FRET),并不容易受到外界環(huán)境的干擾,因此,較為可能的原因是進行光合能量傳遞的載體受到鎘脅迫的抑制,通過藍色非變性凝膠電泳(BN-PAGE)技術(shù)及蛋白質(zhì)組技術(shù)發(fā)現(xiàn)鎘脅迫下魚腥藻[35](Anabaena)、煙草[43]等PSⅡ復(fù)合體CP43 及CP47 蛋白含量顯著降低,進而抑制了激發(fā)能向光化學(xué)反應(yīng)中心的傳遞。
3 重金屬鎘對植物PSⅡ光合能量轉(zhuǎn)化的影響
3.1 植物PSⅡ光合能量捕獲效率對重金屬鎘的響應(yīng)
當(dāng)激發(fā)能傳遞到PS Ⅱ光化學(xué)反應(yīng)中心特殊葉綠素分子P680(P),產(chǎn)生激發(fā)態(tài)P680*,而P680*具有極強的還原性,將在3ps 內(nèi)將1 個電子傳遞給缺鎂葉綠素(Pheo)而產(chǎn)生激發(fā)態(tài)P+·Pheo-,通過這一過程,光合能量由激發(fā)能轉(zhuǎn)化為氧化還原能[1]。理論上光能的傳遞效率超過95%,即被吸收的光量子都能夠參與原初光物理反應(yīng),但每個光子僅有25%的能量可以被轉(zhuǎn)化為化學(xué)能,其余能量轉(zhuǎn)化為熱能,而在實際情況中C3 植物PSⅡ能量轉(zhuǎn)化效率大約為4.3%[44],此外,PS Ⅱ能量轉(zhuǎn)化效率還會受到各類環(huán)境因素的限制。
重金屬鎘會抑制PSⅡ反應(yīng)中心將吸收的激發(fā)能轉(zhuǎn)化為氧化還原能。在一定濃度的鎘脅迫下常發(fā)現(xiàn)PSⅡ?qū)嶋H光化學(xué)效率的下降,如超過0.2 mmol∕L的Cd2+脅迫會使部分品種水稻實際光化學(xué)效率(ΦPSⅡ)及光化學(xué)猝滅系數(shù)(qP)顯著下降[38,39],超富集植物龍葵及芥菜等植物在較低濃度(約25~40 μmol∕L)鎘脅迫下ΦPSⅡ及qP 即受到抑制[41,45],但有研究表明部分品種水稻在鎘脅迫(0.2 mmol∕L)下依然能保持較高的實際光化學(xué)效率[38],甚至在低濃度(0.1 mmol∕L)下略有上升[39]。植物葉片PSⅡ反應(yīng)中心將激發(fā)能轉(zhuǎn)化為氧化還原能的過程對重金屬鎘較為敏感,但不同種類及品種間存在一定差異。
3.2 重金屬鎘對植物PS Ⅱ光合能量轉(zhuǎn)化的影響機制
3.2.1 重金屬鎘通過降低光合能量捕獲、傳遞效率及PSⅡ 反應(yīng)中心數(shù)量影響光合能量轉(zhuǎn)化效率 有學(xué)者提出重金屬鎘引起PSⅡ激發(fā)能轉(zhuǎn)化效率下降的原因可能是重金屬鎘通過抑制激發(fā)能向PSⅡ反應(yīng)中心的傳遞,同時使部分PSⅡ反應(yīng)中心數(shù)量減少,導(dǎo)致PSⅡ反應(yīng)中心對激發(fā)能轉(zhuǎn)化效率的下降[46]。但在另一項研究中發(fā)現(xiàn)鎘脅迫下擬柱孢藻單位活性反應(yīng)中心捕獲的用于電子傳遞的能量(ET0∕RC)較非脅迫條件下下降8.33%,而單位活性反應(yīng)中心RC 捕獲的能量(TR0∕RC)增加6.04%[27],由此可以推測PSⅡ活性反應(yīng)中心的數(shù)量及激發(fā)能捕獲及傳遞效率的降低對于光合能量轉(zhuǎn)化效率的影響主要表現(xiàn)為量的變化,這并不能解釋鎘脅迫下PSⅡ反應(yīng)中心對激發(fā)能轉(zhuǎn)化能力下降的質(zhì)的變化。
3.2.2 重金屬鎘通過抑制光合電子傳遞降低PS Ⅱ光合能量轉(zhuǎn)化效率 PSⅡ活性反應(yīng)中心對激發(fā)能轉(zhuǎn)化的實質(zhì)是傳遞至PSⅡ光化學(xué)反應(yīng)中心的激發(fā)能使P680 發(fā)生電荷分離產(chǎn)生陰陽離子自由基對(P680+·Pheo-)。Pheo-及P680+能否正常得失電子、P680及Pheo 結(jié)構(gòu)的完整性是電荷分離能夠正常且持續(xù)進行的關(guān)鍵,其中光合電子的得失構(gòu)成了光合電子傳遞鏈,且尚沒有證據(jù)表明重金屬鎘會直接引起P680及Pheo 結(jié)構(gòu)的變化[47],因此光合電子傳遞過程受阻可能時是光合能量轉(zhuǎn)化效率下降的主要原因。PSII 的光合電子傳遞易受到重金屬鎘的抑制[48,49],在一定濃度的鎘脅迫下,水稻[38,39]、大豆[50]、菜豆[51]及龍葵[45]等植物PSⅡ光合電子傳遞速率均表現(xiàn)出不同程度的下降。
重金屬鎘分別通過影響PSⅡ受體側(cè)及供體側(cè)抑制光合電子的傳遞過程[36,47,52,53],重金屬鎘不同的作用位點對PSⅡ光化學(xué)反應(yīng)中心存在不同的破壞機制。當(dāng)重金屬鎘使P680受體側(cè)電子傳遞受阻時,P680+·Pheo-發(fā)生電荷重組,產(chǎn)生三線態(tài)P680(3P680),隨后3P680將電子傳遞給O2反應(yīng)生成單線態(tài)氧1O2,而1O2通過直接攻擊P680使PS Ⅱ光化學(xué)反應(yīng)中心失去電荷分離能力,最終引起D1蛋白的裂解,導(dǎo)致光合能量轉(zhuǎn)化效率下降[54]。而當(dāng)重金屬鎘使P680供體側(cè)電子傳遞受阻時,P680+無法接受電子,導(dǎo)致P680+積累,引起β-Car 和Chl a 等色素分子的氧化,并進一步導(dǎo)致D1 蛋白構(gòu)象的變化及其自身水解,最終引起光合能量轉(zhuǎn)化效率下降[54]。
3.2.3 重金屬鎘通過影響PS Ⅱ光化學(xué)反應(yīng)中心結(jié)構(gòu)降低PS Ⅱ光合能量轉(zhuǎn)化效率 PS Ⅱ光化學(xué)反應(yīng)中心由D1-D2-Cyt b559所組成,其結(jié)構(gòu)的完整性也是影響光合能量轉(zhuǎn)化的重要影響因素。其中D1、D2蛋白共同組成異源二聚體通過與P680、Pheo、質(zhì)體醌(QA及QB)結(jié)合,構(gòu)成了原初光反應(yīng)和電子傳遞的關(guān)鍵場所。重金屬鎘能夠直接或間接引起D1、D2蛋白含量的降低,如水稻、小麥等作物在鎘脅迫下D1蛋白顯著下降[55,56],部分水稻品種D2蛋白含量也受到鎘脅迫的影響[55],但D1蛋白對重金屬鎘更為敏感。活性氧(ROS)在重金屬鎘引起D1、D2蛋白降解的過程中可能起重要作用。
Cyt b559 是一種完整的膜蛋白,由α、β 2 個亞基組成,在植物體內(nèi)存在2 種電勢態(tài)(高電勢態(tài)和低電勢態(tài)),Cyt b559 在PS Ⅱ光化學(xué)反應(yīng)中心的生物合成及組裝方面發(fā)揮重要作用[57]。有研究表明Cyt b559 從低電勢態(tài)到高電勢態(tài)的轉(zhuǎn)變是放氧復(fù)合體組裝到PS Ⅱ光化學(xué)反應(yīng)中心的關(guān)鍵步驟之一[58],而重金屬鎘會通過Cd2+與Ca2+位點競爭結(jié)合等多種途徑導(dǎo)致Cyt b559 由高電位向低電位形式轉(zhuǎn)變[59,60],進而影響PS Ⅱ光化學(xué)反應(yīng)中心結(jié)構(gòu)的完整性。此外,也有研究發(fā)現(xiàn)Cyt b559 由高電勢態(tài)轉(zhuǎn)向低電勢態(tài)時PS Ⅱ光化學(xué)反應(yīng)中心放氧速率下降[61],雖然Cyt b559 不直接參與光合放氧過程,但可以通過高低電勢態(tài)轉(zhuǎn)化過程中蛋白構(gòu)象的改變影響放氧復(fù)合物的活性及錳簇組裝活化等過程[62]。
4 小結(jié)
重金屬鎘可以通過降低葉綠素含量、改變?nèi)~綠素的組成、破壞類囊體膜結(jié)構(gòu)等途徑影響植物原初光能轉(zhuǎn)化的生理結(jié)構(gòu)基礎(chǔ)。此外,重金屬鎘會通過改變主體外周捕光復(fù)合體LHC Ⅱb 及內(nèi)周捕光復(fù)合體(CP43、CP47)結(jié)構(gòu)和功能來降低植物葉片光能捕獲、傳遞效率,并通過抑制光合電子傳遞鏈及改變Cyt b559 電勢態(tài)破壞PS Ⅱ光化學(xué)反應(yīng)中心而降低植物對光能的轉(zhuǎn)化效率。
類囊體膜的膜脂和蛋白質(zhì)復(fù)合體的組成及分子結(jié)構(gòu)在維持光合系統(tǒng)的超分子結(jié)構(gòu)、原初光能轉(zhuǎn)化中起重要作用,關(guān)于重金屬鎘對植物類囊體膜脂及蛋白質(zhì)含量的影響已有較多的研究成果,但對構(gòu)成膜脂和蛋白質(zhì)的脂肪酸及氨基酸的變化研究較少,而這些小分子物質(zhì)往往發(fā)揮某些重要的生理功能,如PG 分子中的trans-16:1。因此,重金屬鎘對脂肪酸及氨基酸等小分子代謝物質(zhì)的影響及這些小分子物質(zhì)如何影響PSⅡ-LHCⅡb 的組裝及穩(wěn)定性值得進一步的研究,其中重金屬鎘對LHCⅡb 三聚體結(jié)構(gòu)的影響與光合能量的捕獲與傳遞效率有直接聯(lián)系,應(yīng)該成為研究的重點課題。
重金屬鎘除了影響蛋白質(zhì)含量以外,更多的影響可能來自于鎘對蛋白質(zhì)構(gòu)象的改變,如重金屬鎘脅迫下D1蛋白的水解可能與蛋白質(zhì)構(gòu)象的變化密切相關(guān),因此有必要從蛋白質(zhì)水平上解析重金屬鎘對PSⅡ-LHCⅡb 中關(guān)鍵蛋白各級結(jié)構(gòu)的影響。
植物對光合能量的捕獲、傳遞及轉(zhuǎn)化在很大程度上與PSⅡ-LHCⅡb 的分子空間結(jié)構(gòu)、分子間距離及起分子間的相互作用密切相關(guān)。在光能捕獲及傳遞過程中多涉及激發(fā)能在各色素分子間的傳遞(如Chl b 到Chl a、Chl a 到Chl a 及β-Car 到Chl a 等),即使在PSⅡ光化學(xué)反應(yīng)中心,也存在各輔助色素分子與反應(yīng)中心P680的能量傳遞,因此,進一步探究重金屬鎘對PSⅡ-LHCⅡb 分子空間結(jié)構(gòu)的影響,可以從分子水平上探究鎘脅迫對光合能量傳遞的影響。
在未來的研究中,將從代謝組、蛋白質(zhì)結(jié)構(gòu)及分子空間構(gòu)型等水平,探究重金屬鎘對植物光合能量捕獲、傳遞及轉(zhuǎn)化過程的影響,闡明鎘脅迫下植物光合作用的光物理特征及光化學(xué)反應(yīng)的變化機制,能夠為提高鎘脅迫下植物的光能傳遞、轉(zhuǎn)化效率、減少激發(fā)能損耗,提高光合效率提供理論依據(jù)。
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
甘肅教育(2020年14期)2020-09-11 07:57:42
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國衛(wèi)生(2014年11期)2014-11-12 13:11:32
