牛源犬新孢子蟲NcGRA9基因原核表達及免疫原性分析
2023-03-08 05:01:48金衛東唐澤宇李作臣王海軍賈立軍
中國獸醫學報 2023年2期
金衛東,唐澤宇,李作臣,王海軍,陳 健,賈立軍
(1.延邊大學 東北寒區肉牛科技創新教育部工程研究中心,吉林 延吉 133002;2.延邊特色產業發展中心,吉林 延吉 133002;3.延邊州畜牧總站,吉林 延吉 133002;4.延邊州動物疫病預防控制中心,吉林 延吉 133002)
犬新孢子蟲(Neosporacaninum)是引起孕畜流產、死胎及運動神經系統障礙的胞內寄生性原蟲,主要寄生于動物的腦、肝臟、脾臟等臟器組織以及生殖系統[1-2]。新孢子蟲病(Neosporosis)呈世界性分布,我國北京、山東、吉林、新疆、青海等多個省、市(區)都有該病的報道[3]。犬新孢子蟲對牛的危害最為嚴重,羊、馬、驢、鹿、狐、鴿、水貂等多種動物均有該病感染的報道[4-5]。目前尚無用于預防新孢子蟲病的商品化疫苗[6]。
致密顆粒蛋白(GRA)是新孢子蟲致密顆粒細胞器分泌的蛋白,致密顆粒蛋白被釋放到寄生液泡(pv)中,在維持寄主-寄生關系和獲取營養物質方面發揮重要作用[7]。目前鑒定得到的新孢子蟲GRA9蛋白與弓形蟲GRA9蛋白同源性達到60%,且NcGRA9主要是親水蛋白質,但也包含較短的疏水區。而NcGRA9蛋白作為新孢子蟲疫苗的候選抗原之一,國內尚未有該蛋白研究的相關報道,本試驗PCR擴增新孢子蟲NcGRA9基因,構建NcGRA9重組表達載體,誘導表達重組蛋白并進行純化,通過Western blot鑒定和小鼠免疫來驗證NcGRA9蛋白的抗原性,以期為新孢子蟲疫苗的研究奠定基礎。
1 材料與方法
1.1 蟲株和實驗動物牛源犬新孢子蟲、Vero細胞、pGEX-4T-1表達載體均由延邊大學預防獸醫實驗室提供;BALB/c小鼠購自延邊大學實驗動物中心。
1.2 主要試劑BamHⅠ 酶、Not Ⅰ酶、GST純化試劑盒、BCA試劑盒均購自生工生物工程(上海)股份有限公司;HRP標記的山羊抗小鼠IgG、蛋白質Marker均購自北京索萊寶科技有限公司;其他試劑均為國產分析純。
1.3 引物的設計與合成根據ToxoDB網站發布的NcGRA9(NCLIV_066630)基因序列,應用Oligo 7.0軟件設計特異性引物,引物序列P1:5′-TCTTATCGGGATCCATGCAGGGCGTGAC-3′,P2:5′-ATAAGAATGCGGCCGCTATTTCTCCGTT-ATGGTTCG-3′。
1.4 NcGRA9基因PCR擴增以提取的新孢子蟲基因組DNA作為模板,進行PCR擴增。反應條件:94℃ 5 min;94℃ 45 s,56℃ 45 s,72℃ 45 s,35個循環;72℃ 7 min。PCR產物進行1%瓊脂糖凝膠電泳。
1.5 NcGRA9基因重組克隆質粒的構建及測序將PCR擴增正確的條帶回收,連接pMD18-T克隆載體,轉化到DH5α中,提取重組質粒進行PCR和雙酶切鑒定,并進行測序分析。
1.6 NcGRA9基因原核表達質粒的構建雙酶切pGEX-4T-1載體,瓊脂糖凝膠回收載體片段。將NcGRA9基因與pGEX-4T載體4℃過夜連接。
1.7 表達蛋白SDS-PAGE電泳和Western blot分析將pGEX-4T-NcGRA9重組質粒轉化至BL21感受態細胞中IPTG誘導表達,GST標簽蛋白純化試劑盒純化表達蛋白。SDS-PAGE電泳后,進行Western blot分析。
1.8 重組蛋白的小鼠免疫將純化后的重組蛋白與弗氏佐劑(完全佐劑和不完全佐劑)混合。將30只BALB/c小鼠隨機分為3組,每組10只,分為免疫組(重組蛋白+佐劑)、質粒組(pGEX-4T-NcGRA9)及對照組(PBS)。免疫組的重組蛋白含量200 μg/只,質粒組為等量pGEX-4T-NcGRA9,對照組為等體積PBS。每次接種間隔14 d,共接種3次,每次接種前及第3次接種后14 d采集血液,分離血清并保存待檢。
1.9 免疫小鼠血清中抗體水平和細胞因子水平檢測應用間接ELISA方法[8]檢測免疫小鼠血清中IgG抗體水平,按照ELISA試劑盒說明書檢測免疫小鼠血清中抗體亞類IgG1、IgG2a水平和細胞因子IFN-γ、IL-4水平。
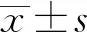
2 結果
2.1 牛源犬新孢子蟲NcGRA9基因的克隆與測序分析以牛源犬新孢子蟲的DNA為模板,經PCR擴增出大小為522 bp的條帶,擴增條帶與預期大小相符。構建pMD18-T-NcGRA9重組克隆質粒,經PCR和雙酶切鑒定,PCR擴增出522 bp片段;經NotⅠ與BamHⅠ雙酶切后,獲得522 bp的目的條帶與2 692 bp的載體片段。經測序表明,獲得的NcGRA9基因序列與ToxoDB網站發布NcGRA9(NCLIV_066630)基因序列同源性為100%。
2.2 重組表達質粒pGEX-4T-NcGRA9鑒定pGEX-4T-NcGRA9進行PCR和雙酶切鑒定,PCR擴增出522 bp目的片段;經NotⅠ、BamHⅠ雙酶切后,獲得522 bp的目的條帶與4 969 bp的載體片段(圖1)。

M.DL2000 DNA Marker;1.pGEX-4T-NcGRA9雙酶切產物
2.3 蛋白的表達與純化取誘導3 h后的重組蛋白和純化后的重組蛋白,進行SDS-PAGE電泳,在約45 kDa處出現目的條帶,并以可溶性蛋白表達(圖2)。Western blot分析表明,該蛋白能被小鼠抗新孢子蟲陽性血清所識別,具有較好的反應原性(圖3)。

M.蛋白質Marker;1.誘導后總蛋白;2.純化后的蛋白;3.沉淀樣本;4.上清樣本

M.蛋白質Marker;1.NcGRA9重組蛋白;2.陰性對照
2.4 免疫小鼠血清中IgG、IgG1及IgG2a抗體水平檢測結果ELISA檢測免疫小鼠血清中IgG、IgG1、IgG2a抗體水平(圖4),經統計學分析結果顯示,免疫重組蛋白后,免疫組小鼠IgG抗體及其亞類IgG1、IgG2a水平均極顯著高于質粒組與PBS對照組(P<0.01)。

**P<0.01表示差異極顯著;*P<0.05表示差異顯著;P>0.05表示差異不顯著。下同
2.5 免疫小鼠血清中IFN-γ、IL-4細胞因子水平檢測結果ELISA試劑盒檢測免疫小鼠血清中細胞因子IFN-γ和IL-4水平(圖5),經統計學分析,免疫組小鼠IFN-γ與IL-4水平均極顯著高于質粒組與PBS對照組(P<0.01)。

圖5 不同免疫組小鼠血清IFN-γ(A)、IL-4(B)的水平檢測
3 討論
新孢子蟲病嚴重影響我國的畜牧業發展,而目前尚無有效藥物和疫苗防控新孢子蟲病。國內外對NcGRA2、NcGRA6、NcGRA7、NcGRA16等均有不同程度的研究報道,而對NcGRA9的研究較少[9]。國內金春梅等[10]對NcGRA2進行了表達與鑒定;刑明等[11]對NcGRA6基因進行表達,并對其免疫效果進行初步分析;賈立軍等[12]對NcGRA7進行了表達;杜博亞等[13]對NcGRA16進行了初步分析;但尚未見對NcGRA9的相關研究報道。
致密顆粒蛋白質結構和功能多樣,在宿主細胞入侵后被釋放到寄生液泡中,使蟲體獲得營養以滿足生長增殖的需要[14]。近年來研究發現,致密顆粒蛋白在參與蟲體入侵宿主細胞后以高水平分泌,在其后處于較低水平。NcGRA9作為可溶性蛋白存在于液泡中,在宿主細胞入侵中起重要作用,并且C末端截短的NcGRA9蛋白保留在這些細胞器中因為折疊不當,隨后可能會退化。
以往的研究證實,GRA2、GRA6和GRA7的抗原分子是新孢子蟲有效的候選疫苗蛋白基因[15]。而關于NcGRA9疫苗方面的研究甚少,僅有YU等[16]將新孢子蟲致密顆粒蛋白NcGRA1、NcGRA4、NcGRA9、NcGRA14、NcGRA17及NcGRA23構建的pcNcGRA DNA疫苗接種BALB/c小鼠,這些真核表達質粒可引起強烈的Th1型免疫反應,誘導機體產生IgG2a抗體和IFN-γ細胞因子水平,免疫保護試驗表明DNA疫苗均能夠延長小鼠的存活時間和減少腦內蟲荷量。由此可見,NcGRA9基因作為疫苗抗原的可行性。
為了解NcGRA9基因的表達形式及作為疫苗候選抗原的免疫原性,本試驗對NcGRA9基因進行了原核表達,確定了NcGRA9基因表達重組蛋白為可溶性蛋白,Western blot分析具有良好的反應原性;接種BALB/c小鼠能夠刺激機體產生細胞免疫應答與體液免疫應答,具有良好的免疫原性。本試驗為NcGRA9基因蛋白作為新孢子蟲病診斷的候選抗原和疫苗候選抗原奠定了基礎。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
