重組豬干擾素α的原核表達及抗病毒活性
2023-03-08 05:01:54李秀麗張武超雷白時袁萬哲
中國獸醫學報 2023年2期
李秀麗,韓 穎,趙 款,張武超,梁 飛,楊 歡,雷白時*,袁萬哲*
(1.河北農業大學,河北 保定 071001;2.河北省獸醫生物技術創新中心,河北 保定 071001;3.石家莊市動物基因工程藥物技術創新中心,河北 石家莊 050800;4.河北伯瑞動物藥業有限公司,河北 正定 050800;5.山東省莒南縣動物疫病預防控制中心,山東 莒南 276600)
20世紀30年代,人們發現機體感染一種病毒后,會出現阻止另一種毫無關系的病毒增殖的干擾現象(viral interference)[1]。1957年,有報道利用雞胚絨毛尿囊膜研究流感病毒的干擾現象時,發現被感染的細胞能產生一種因子,稱為干擾素(IFN)[2]。IFN在病毒感染和細胞免疫中起著至關重要的作用,是體內的第一道防線。IFN家族分為3種不同的類型:即Ⅰ、Ⅱ、Ⅲ型。Ⅰ型IFN包括IFN-α、IFN-β、IFN-ε、IFN-ω、IFN-κ、IFN-δ、IFN-τ、IFN-ζ[3],IFN-α是其中之一。Ⅱ型僅有γ型1種,含有內含子。Ⅲ型IFN家族(IFN-λ) 由IL-29(IFN-λ1)、IL-28A(IFN-λ2)和IL-28B(IFN-λ3)組成[4]。
在人醫方面,IFN的研究較為全面,在動物方面,豬IFN(porcine interferon)是被最早研究的動物IFN之一,20世紀80年代就有關于豬IFN的報道。在各種類型的IFN當中,IFN-α的抗病毒作用最強,是異質性的,可以分為不同的亞型,目前已知有20余種亞型[5]。IFN通過結合細胞膜上的特異性受體,引發信號放大過程,將信號最后傳遞到細胞核內,對信號下游的基因表達進行調控,引發各種生理反應。在病毒侵襲的各個階段都有IFN刺激基因(ISGs)的參與,如Mx1、OAS、IFIT1、ISG15等,它們具有抗病毒及免疫調節等多種功能,能夠通過不同的途徑抵御病毒入侵和復制[6-7]。
我國是養豬大國,病毒性豬病種類繁多,危害巨大。這些豬病的流行特點是病毒變異速度快,發病特征非典型性,并且容易暴發。由于豬IFN-α是一類具有廣譜抗病毒、抗腫瘤和免疫調節等多種作用的糖蛋白,所以被廣泛應用于病毒性疾病的治療和預防,這已逐漸引起研究人員的關注[8]。腦心肌炎病毒(EMCV)是一種重要的人畜共患病病原體,感染后可引起動物和人類發生腦炎和心肌炎,以及神經系統疾病、糖尿病和繁殖障礙等多種疾病[9]。豬偽狂犬病病毒(PRV)感染主要是通過控制轉錄水平實現的,減壓閥的基因表達可以分為3個階段:立即早期基因(IE)、早期基因(E)和晚期基因(L)[10]。已有研究證實PoIFN-α可以有效治療口蹄疫[11]、豬流行性腹瀉[12]、豬繁殖與呼吸綜合征等[13]病毒性疾病,但rPoIFN-α是否可以有效抑制EMCV和PRV,相關研究報道較少。
近年來,基因重組技術迅速發展,己廣泛應用于各種生物制品的生產。基因工程重組人IFN-α是應用重組技術生產的最早的蛋白藥品之一[14],也是當今能自主生產的少數基因工程藥物之一,利用基因重組技術獲得的人IFN,不僅實現了產業化,同時保留了蛋白的天然活性。鑒于此,本試驗將豬IFN-α基因克隆入原核表達載體上,轉入大腸桿菌中,由此獲得工程菌株,實現高表達。經一系列的體內體外研究發現,rPoIFN-α在體內外均具有良好的抗病毒活性,這為進一步重組蛋白生產的工業化提供了理論依據。
1 材料與方法
1.1 主要材料DMEM培養基、胎牛血清(FBS)購自美國Gibco公司;小鼠96T ELISA試劑盒購自北京冬歌生物技術有限公司;EMCV、PRV、豬睪丸細胞(ST)和豬腎細胞(PK-15)均由本實驗室保存;水泡性口炎病毒(VSV)由中國農業科學院上海獸醫研究所惠贈;BALB/c小鼠購自北京華阜康生物科技股份有限公司。
1.2 rPoIFN-α的表達、純化和復性參照GenBank登錄的PoIFN-α序列(登錄號:KF414740.1)將信號肽序列去除,優化后合成重組質粒pET-32a-PoIFN-α。將重組質粒pET-32a-PoIFN-α 轉入感受態細胞 BL21(DE3),使用引物PoIFN-F和PoIFN-R(表1)對單菌落進行鑒定。陽性菌進行擴大培養,按體積比1∶100接種LB中(含Amp),37℃、200 r/min培養,待D600為0.4~0.6時,加入終濃度1.0 mmol/L 的IPTG,誘導4 h,離心收集菌體,超聲波破碎(360 W、30%、10 min)2次后,分離上清液和沉淀,進行SDS-PAGE檢測。參照Ni-Agatose Resin的純化步驟進行純化,采用透析復性法對蛋白進行復性。用0.22 μm 濾膜過濾除菌,分裝,-20℃ 保存。
1.3 重組蛋白Western blot檢測以Anti-His MousemAb為一抗,按照1∶5 000進行稀釋,充分混勻后覆蓋在PVDF膜上,室溫孵育1 h。以Goat Anti-MouseIgG作為二抗,按照1∶10 000進行稀釋,室溫孵育1 h,使用ECL顯色。
1.4 rPoIFN-α的細胞毒性檢測用MTT法測定rPoIFN-α作用PK-15和ST細胞后的活性。PK-15細胞在96孔細胞培養板生長至單層后,取制備好的重組蛋白用含2% FBS的DMEM營養液進行4倍倍比稀釋,共稀釋8個梯度,每個梯度8個重復孔。每孔100 μL,同時設置蛋白陽性對照孔及細胞陰性對照孔,孵育16~24 h后不棄培養基,加入10~20 μL MTT(質量濃度為5 g/L),包裹錫紙避光,37℃ 4 h。棄掉培養基,加入DMSO,100 μL/孔,避光搖勻,15 min內測定D450值。
1.5 rPoIFN-α在體外的抗病毒活性采用細胞病變抑制法測定rPoIFN-α蛋白在PK-15細胞上抗VSV增殖活性。PK-15在96孔細胞培養板生長至單層后,4倍倍比稀釋IFN,共稀釋9個梯度,每個梯度8個重復孔。每個梯度取100 μL加至96孔板中,于37℃、5% CO2細胞培養箱中孵育12~16 h,棄去培養液。將保存的VSV、PRV和EMCV溶液用含 2% FBS的DMEM營養液稀釋至100 TCID50,每孔加 100 μL。同時設計陽性對照(只用病毒處理),陰性對照(共稀釋 64 倍的蛋白處理)和正常細胞對照(不做處理)。當觀察到的陽性對照組細胞出現75%以上病變時判定結果,按照Reed-Muench 法計算重組蛋白的抗病毒活性。
1.6 IFN刺激基因的表達水平使用能夠完全抑制VSV感染且劑量較低的rPoIFN-α蛋白對PK-15進行孵育,于不同時間(6,12,24,36,48 h)收集細胞,同時設置DMEM組,TRIzol法抽提細胞總RNA,將總RNA樣品反轉錄得到cDNA,引物序列見表1。RT-qPCR反應體系20 μL:2×AgueGreen qPCR Master Mix 10 μL,上、下游引物各1 μL,cDNA模板1 μL,ddH2O 7 μL。反應條件:95℃ 5 min;95℃ 20 s,58℃ 20 s,72℃ 20 s,45個循環;進行熔解曲線分析,95℃ 10 s,65℃ 60 s,97℃ 1 s。
1.7 rPoIFN-α在小鼠體內的抗病毒活性6周齡BALB/c 小鼠25只隨機分為5組,分別為對照組、EMCV組、PRV組、 EMCV+rPoIFN-α組和PRV+rPoIFN-α組,EMCV病毒液3×105TCID50/0.1 mL,0.1 mL/只腹腔注射;PRV病毒液5×102TCID50/0.1 mL,0.1 mL/只背部皮下注射,攻毒1 d后腿部肌肉注射重組蛋白,50 μg/只。給藥后3,5,7,10,14 d每組尾尖采血,測定血清細胞因子TNF-α、IL-6、IL-10和IL-1β的變化;第14天全部處死,收取腦、心臟、肝臟和脾臟組織測定病毒載量,同時腦部組織測定IE180、EPO轉錄水平;在整個試驗期間記錄體質量變化以及根據臨床癥狀進行打分。
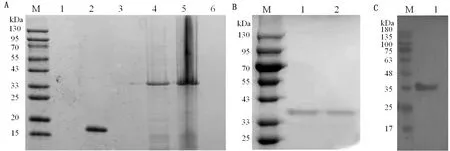
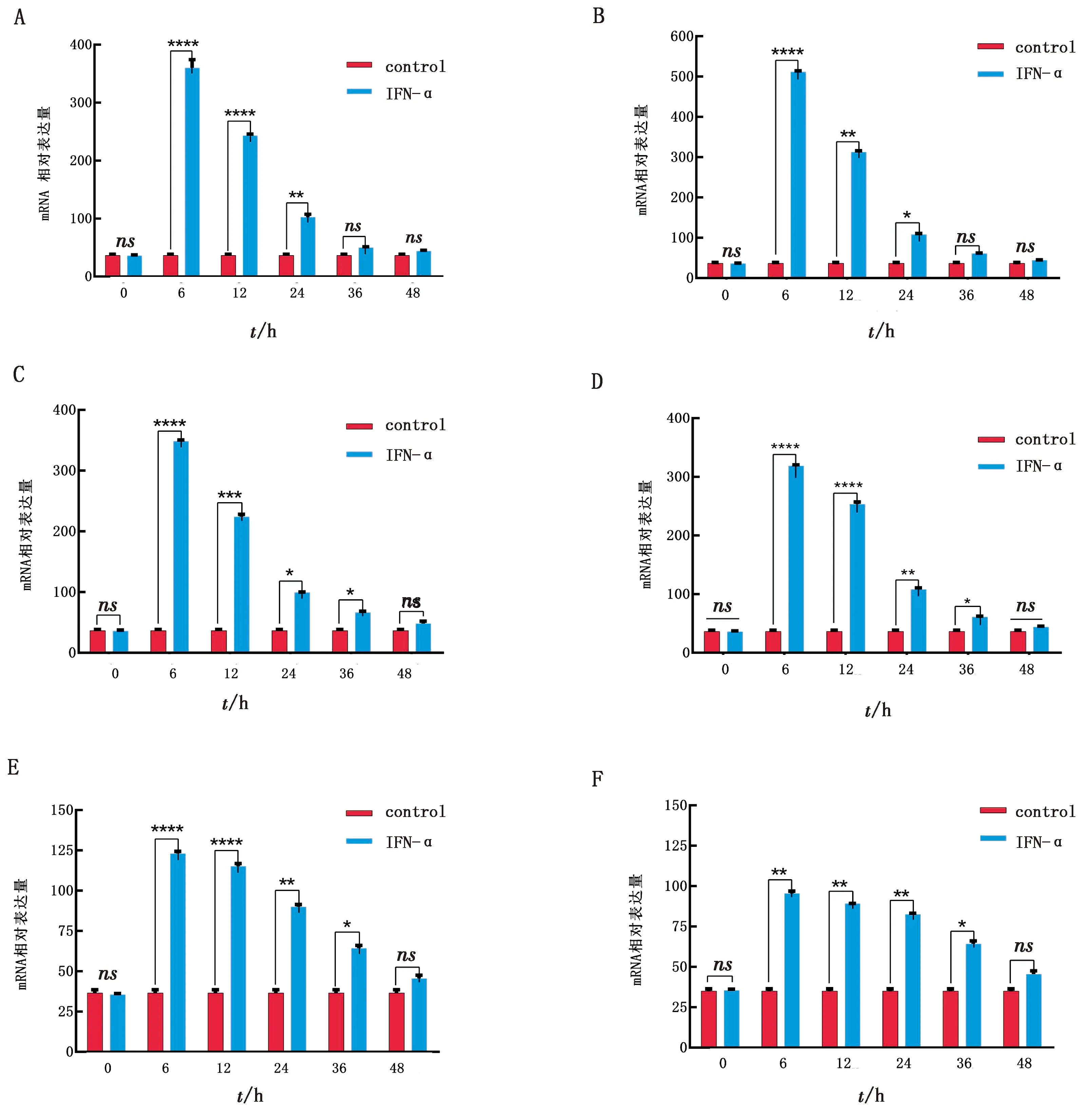
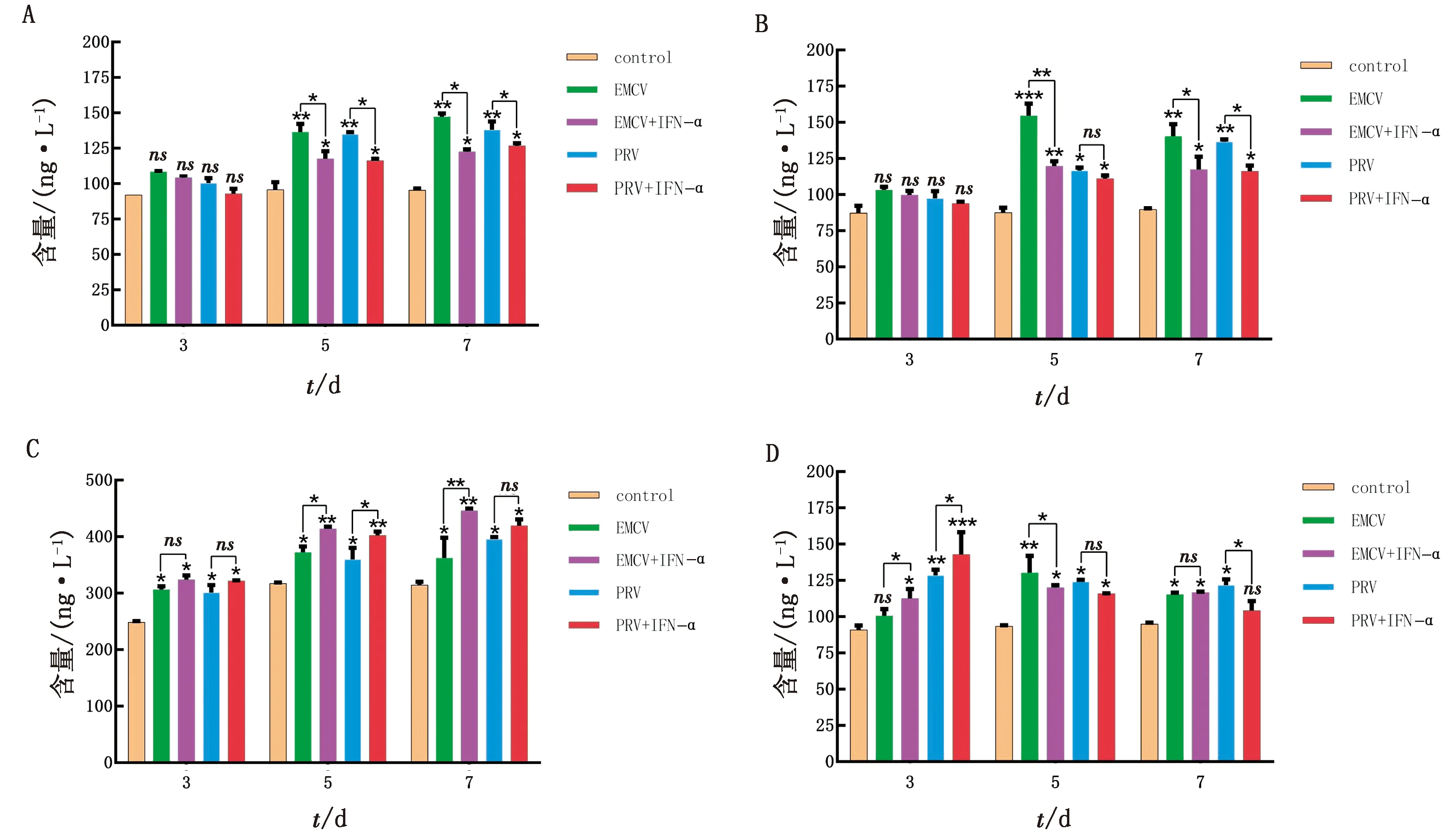
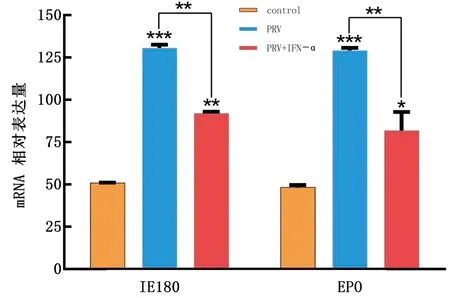
1.8 統計分析每個試驗重復3次。所有統計分析均采用SPSS 16.0軟件進行,通過RT-qPCR 檢測后,數據處理采用 2-△△Ct法計算。以*0.01 表1 引物信息 2.1 rPoIFN-α的表達、純化和復性將合成的rPoIFN-α小提質粒轉入BL21中,經T7通用引物和序列特異性引物鑒定。將成功轉化后的陽性菌誘導表達,在1.0 mmol/L的IPTG誘導下進行可溶性分析,分別取誘導前全菌、誘導后的上清、沉淀和空載質粒進行SDS-PAGE分析,結果表明rPoIFN-α為包涵體表達;按照Ni-Agatose Resin的包涵體蛋白純化說明書進行蛋白的純化,透析復性后使用Nano Drop2000測定蛋白質量濃度平均為1.0 g/L,并進行SDS-PAGE鑒定,由圖1可知,純化復性后蛋白大小正確(35 kDa),條帶單一。經Western blot驗證,成功獲得特異性目的蛋白。 A.重組蛋白的可溶性鑒定(M.蛋白Marker;1~6.pET-32a空載體未誘導、pET-32a空載體誘導、全菌未誘導、全菌誘導、沉淀、上清);B.重組蛋白純化與復性(M.蛋白Marker,1~2.分別為純化和復性后SDS-PAGE,大小為35 kDa);C.重組蛋白Western blot驗證(M.蛋白Marker;1.重組蛋白) 2.2 rPoIFN-α的細胞毒性如圖2所示,MTT法測定rPoIFN-α對細胞活力的影響,在PK-15和ST細胞上,隨著蛋白稀釋倍數的增加,其對細胞活性的影響越接近對照組,當稀釋度為4-4時,80%以上的細胞未受損傷,這為后續試驗提供可能。 A.蛋白作用后PK-15細胞活力;B.蛋白作用后ST細胞活力。**示P<0.01。下同 2.3 rPoIFN-α在體外抗病毒活性通過PK-15/VSV系統檢測rPoIFN-α蛋白的抗病毒活性,參照Reed-Muench法,計算距離比例為(高于50%細胞無病變百分比-50%)/(高于50%細胞無病變百分比-低于50%細胞無病變百分比),可得重組蛋白的活性效價為1.25×105U/mL。同理測得rPoIFN-α在PK-15和ST細胞上抗EMCV和PRV的活性(表2)。 表2 rPoIFN-α在不同細胞上抗病毒活性 U/mL 2.4 ISGs轉錄水平的測定收取rPoIFN-α作用后的PK-15細胞,測定Mx1、OAS、IFIT1、ISG15、MHC-Ⅰ和MHC-Ⅱ的轉錄水平。結果顯示,重組IFN孵育細胞后,與DMEM對照組相比顯著上調了各細胞因子,且在6 h時,表達水平達到峰值(圖3)。 A~F.ISG15、Mx1、OAS、IFIT1、MHC-Ⅰ和MHC-Ⅱ在0,6,12,24,36,48 h時的表達水平 2.5 rPoIFN-α在小鼠體內抗病毒作用 2.5.1生存曲線 根據試驗期間小鼠的死亡情況繪制了生存曲線,如圖4所示,EMCV、PRV組在4 d 出現死亡,EMCV、PRV組分別在9,10 d全部死亡,病死率為100%,EMCV治療組病死率為20%,PRV治療組病死率為0%。 圖4 病毒感染組以及IFN治療組小鼠的死亡情況 2.5.2各組織器官病毒載量 第14天處死全部小鼠,取腦、心臟、脾臟、肝臟組織,測定各組織的病毒載量,如圖5所示,EMCV治療組腦中病毒載量低于EMCV陽性對照組,呈極顯著差異;脾臟中EMCV載量也顯著低于對照組。 A.EMCV感染組和rIFN-α治療組心臟、肝臟、脾臟和腦的病毒載量;B.PRV感染組和IFN-α治療組心臟、肝臟、脾臟和腦的病毒載量 2.5.3血清細胞因子水平的變化 EMCV和PRV感染后,IL-6、IL-1β、TNF-α的水平升高,重組IFN治療后5~7 d,三者水平較未感染組略有升高,但仍低于感染組,而治療組IL-10的水平在3,5,7 d均高于病毒感染組(圖6)。 A~D.依次為3,5,7 d時,小鼠血清中TNF-α、IL-1β、IL-10和IL-6的含量 2.5.4臨床癥狀 病毒感染后,觀察記錄小鼠表現,當出現體質量減輕、被毛粗亂、互相啃咬、采食量降低和神經癥狀時分別記1分,沒有癥狀記為0分,根據所得分值繪制曲線。結果顯示,在第3天時病毒感染組開始出現癥狀,而重組IFN治療后在第5天才開始出現輕微的癥狀(圖7)。 2.5.5IE180、EPO的轉錄水平 取小鼠腦組織,測定其相對表達量。結果顯示,病毒感染后IE180和EPO的表達水平均顯著高于未感染組和治療組,這說明用重組IFN治療后,通過減少2種蛋白的轉錄,抑制了PRV的增殖,起到抗病毒效果(圖8)。 圖7 小鼠臨床癥狀得分情況 圖8 PRV感染組和IFN-α治療組小鼠腦組織中IE180、EPO的表達水平 當今豬病形勢復雜,病毒性疾病占主導地位,特別是近幾年暴發的豬流行性腹瀉、豬偽狂犬病給養殖業造成了巨大的經濟損失[15]。IFN生物制劑由于其自身優勢成為了新的研究熱點。1986年,LEFEVRE等[16]首次利用大腸桿菌表達PoIFN-α,因其具有速效多能、無殘留、無副作用等特點,且不會對人類健康造成危害,已成為替代化學合成抗病毒藥物的理想新藥。因此我國學者也相繼開展了PoIFN-α的研究,實現了PoIFN-α的體外表達,在生產實踐中操作簡單、產量大,有良好的應用前景[17]。 本試驗成功獲得穩定表達的rPoIFN-α,并使用多種體外系統進行了活性測定。其中,VSV/PK-15系統是豬IFN抗病毒活性檢測系統中使用最廣泛的活性測定方法[18]。本研究表達的rPoIFN-α抗病毒比活性達到1.67×106IU,MTT法測定rPoIFN-α細胞的增殖活性,結果發現,當重組蛋白稀釋到4-4時,80%以上的細胞未損傷,這說明rPoIFN-α對豬源PK-15和ST細胞的損傷均在可控范圍內,與吳夢礬[19]的研究相比有了較大提升。Ⅰ型 IFN 通過 JAK-STAT 信號通路發出信號,以刺激 ISGs 等抗病毒效應物的產生。通過檢測IFN信號通路下游ISGs 的轉錄水平,能夠更直觀地證實重組融合蛋白的抗病毒潛力。試驗獲得的rPoIFN-α在作用PK-15細胞后6 h顯著上調Mx1、OAS、IFIT1、ISG15基因的表達水平,結果與報道相一致[20],在檢測的細胞因子中,MHC-I和 MHC-Ⅱ的表達不顯著,這與其抗病毒機制有關。 在小鼠體內,EMCV和PRV感染組各組織器官的病毒載量均高于IFN-α治療組,病毒滴度在腦內最高,其原因可能與PRV和EMCV屬嗜神經病毒有關。結果表明,rPoIFN-α可以有效抑制病毒粒子在腦、心臟、肝臟和脾臟中的增殖,與病毒感染組相比具有顯著性差異。細胞因子參與免疫和炎癥反應,在保護身體免受外來病原體侵害方面發揮關鍵作用,TNF-α、IL-Iβ和IL-6屬于促炎因子,參與促進急性炎癥反應以防御感染。本研究中,病毒感染后促炎細胞因子IL-1β、IL-6和TNF-α水平升高;這提示PRV和EMCV激活了先天免疫[21]。而rPoIFN-α治療組,這些促炎細胞因子水平在5 d時低于感染對照組,提示rPoIFN-α可在感染早期減弱炎癥反應,抑制病毒復制,在7 d時逐漸恢復正常,避免嚴重炎癥的發生;與此同時,rPoIFN-α治療組的小鼠體內IL-10水平在5,7 d顯著上升,高于病毒感染組,IL-10作為抗炎因子之一,結果表明rPoIFN-α起到了積極的抗病毒作用。根據EMCV和PRV感染小鼠后出現的癥狀,設置打分標準,對試驗期間的小鼠進行觀察記錄,同時記錄小鼠的死亡情況,結果顯示,EMCV和PRV感染組在4 d出現死亡,于9,10 d全部死亡,而PRV治療組全程未出現死亡,EMCV治療組在6 d死亡2只,其余均存活;根據打分結果,治療組小鼠出現臨床癥狀較感染組晚,且癥狀較輕。 病毒復制的關鍵步驟包括病毒基因轉錄,因此,通過阻斷基因轉錄抑制病毒增殖是一種有效的方法。IE180是PRV唯一的直接早期基因,是一種可啟動早期基因轉錄的轉錄激活因子。EPO是PRV復制周期的早期蛋白之一,其水平的高低意味著PRV增殖的快慢。在本研究中,重組豬IFN治療組IE180和EPO的表達水平均低于PRV感染組,即PRV在小鼠體內增殖速度減慢,這也說明了重組豬IFN可在小鼠體內起到抗PRV的作用。 本試驗結果表明,原核表達rPoIFN-α具有理想的體內外抑制PRV和EMCV的活性,為今后rPoIFN-α抑制PRV和EMCV的臨床試驗奠定了基礎。
2 結果





3 討論
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
學苑創造·A版(2015年11期)2016-01-14 09:03:27
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
