人源GRK2的真核表達、純化及活性檢測
2023-03-02 08:43:42蔣勵萍陳露穎蒯佳婕王鳳玲關艷玲韓陳陳
安徽醫科大學學報 2023年2期
蔣勵萍,陳露穎,蒯佳婕,王鳳玲,李 浩,關艷玲,馬 旸,韓陳陳,魏 偉
G蛋白偶聯受體激酶 2(G protein-coupled receptor kinase 2, GRK2)屬于絲/蘇氨酸激酶,參與調節G蛋白偶聯受體(G protein coupled receptors, GPCRs)的磷酸化、脫敏和內化[1]。GRK2活性和表達的改變在類風濕關節炎(rheumatoid arthritis, RA)等炎癥免疫相關疾病中起著重要作用[2]。課題組前期研究[3-5]表明,GRK2與前列腺素E2受體4 (prostaglandin E2 receptor 4, EP4)結合,介導受體過度脫敏,導致RA中成纖維樣滑膜細胞(fibroblast-like synoviocytes, FLS)的異常增殖和巨噬細胞極化。因此,GRK2可能通過調控與其結合的蛋白活性或表達,從而發揮多種生理和病理作用。
為了研究GRK2調控EP4的分子機制,本研究利用前期獲得的GRK2野生型質粒,連接到高效真核表達載體pcDNA3.1-EGFP上,構建重組質粒。將重組質粒轉染到HEK 293T 細胞中進行蛋白表達,通過 Ni-NTA 純化及超濾獲得 His-GRK2 重組蛋白。采用SDS-PAGE、Western blot、免疫共沉淀及His pull down等方法,鑒定GRK2重組蛋白的表達和生物活性,旨在為進一步研究GRK2蛋白對EP4的調控作用奠定基礎。
1 材料與方法
1.1 實驗材料
1.1.1細胞、菌株、載體 人胚腎HEK 293T細胞購自中國科學院上海細胞庫;pcDNA3.1-EGFP質粒、pIRES-EGFP-GRK2、pIRES-EGFP-EP4 質粒由本課題組前期構建和保存;大腸埃希菌DH5ɑ 購自上海唯地生物技術有限公司。
1.1.2主要試劑 胎牛血清(fetal bovine serum,FBS)、DMEM 培養基及PBS購自以色列BI公司;Prime STAR酶、限制性內切酶Hind Ⅲ、限制性內切酶 BamH Ⅰ、T4 DNA連接酶、DNA Marker DL10000購自日本TaKaRa公司;切膠回收試劑盒和質粒小提取試劑盒購自TIANGEN天根生化科技(北京)有限公司;兔源GRK2、鼠源EP4購自美國Santa Cruz公司;山羊抗兔和山羊抗鼠二抗購自北京中杉金橋生物技術有限公司;6FF瓊脂糖凝膠微珠購自美國GE公司;JetPRIME transfection reagent轉染試劑購自法國PolyPlus公司;考馬斯亮藍染色液購自上海碧云天生物技術有限公司;PVDF膜和蛋白超濾管(截留30~50 ku蛋白)購自美國Millipore公司。
1.1.3主要儀器 T100TMPCR 儀購自美國 Bio-Rad公司;核酸電泳儀和 DYY-6C 型電泳系統購自北京六一公司;Scientz-IID 超聲波細胞粉碎機購自南京賽飛生物科技有限公司;Tanon 1600全自動數碼凝膠成像系統購自上海天能科技有限公司;ImageQuant LAS 4000熒光及化學發光成像系統購自美國GE公司。
1.2 方法
1.2.1引物設計、合成及PCR擴增 從 https://www.ncbi.nlm.nih.gov/pmc/ 網站獲取人源 GRK2 基因序列(NM 001 619.5),設計并添加Hind Ⅲ 和BamH Ⅰ 酶切位點和C端6×His標簽(CATCATCACCATCACCAT)的引物擴增全長約為2 067 bp的GRK2基因,上游引物:5′-CCCAAGCTTATCCACCTGACCATGAATG-3′ (下劃線部分是Hind Ⅲ 酶切位點),下游引物:5′-CGCGGATCCATGGTGATGGTGATGATGGAGGCCG-3′ (下劃線部分是BamH Ⅰ 酶切位點,加粗是6×His標簽),所有引物由通用生物(安徽)股份有限公司合成。PCR反應體系 (50 μl):cDNA 1 μl、上游引物 1 μl、下游引物 1 μl、Prime Star DNA 聚合酶 0.5 μl、dNTP 4 μl、5×Prime Star buffer 10 μl、H2O 32 .5 μl。反應程序如下:30個循環(98 ℃ 10 s,55 ℃ 5 s,72 ℃ 2 min) 進行擴增。將PCR產物進行1.0% 瓊脂糖凝膠電泳,出現約2 067 bp 的條帶時利用瓊脂糖凝膠DNA回收試劑盒回收產物。
1.2.2pcDNA3.1-EGFP-His-GRK2重組真核表達質粒的構建及雙酶切鑒定 將His-GRK2 的PCR產物回收片段與pcDNA3.1-EGFP 質粒,37 ℃,Hind Ⅲ 和BamH Ⅰ 雙酶切2 h,產物分別經1.0%瓊脂糖凝膠電泳分離,切膠回收試劑盒回收產物。用T4 DNA連接酶在4 ℃條件下連接 His-GRK2 基因片段回收產物與pcDNA3.1-EGFP 回收產物 12 h。連接產物轉化DH5α感受態細胞,轉化液涂板,37 ℃培養12~16 h,挑取菌落搖菌培養,利用質粒提取試劑盒提取質粒。
1.2.3pcDNA3.1-EGFP-His-GRK2重組真核表達質粒的測序驗證 挑選重組質粒,經 Hind Ⅲ 和BamH Ⅰ 雙酶切鑒定為陽性克隆后送至南京擎科生物科技有限公司進行測序,以載體的通用引物進行測序鑒定。測序結果利用https://blast.ncbi.nlm.nih.gov網址進行測序比對。
1.2.4細胞培養與pcDNA3.1-EGFP-His-GRK2重組真核表達質粒的轉染 細胞準備:人HEK 293T細胞以每孔1×106個細胞接種于6孔板中,以10% DMEM 培養液培養,置于37 ℃、5 % CO2箱中孵育24 h,至細胞達到80%~90%融合進行轉染。
轉染方法(6孔板):① 制備轉染復合物:取無菌1.5 ml EP管,加入200 μl的jetPRIME buffer,再加入2 μg DNA,快速渦旋10 s,混勻后,加入4 μl的jetPRIME reagent轉染試劑,快速渦旋10 s,混勻后室溫靜置10 min。② 轉染:將6孔板中培養基棄去,加入2 ml 10%DMEM,將轉染復合物加入孔中,將其置于37 ℃、5% CO2孵箱內培養48 h,觀察轉染效果。
1.2.5倒置熒光顯微鏡觀察pcDNA3.1-EGFP-His-GRK2轉染效率 HEK 293T細胞轉染pcDNA3.1-EGFP-His-GRK2質粒后(帶有EGFP標簽),培養48 h,在倒置熒光顯微鏡下觀察GRK2的轉染效率并拍照。
1.2.6Western blot鑒定GRK2蛋白在細胞中的表達 HEK 293T細胞轉染pcDNA3.1-EGFP-His-GRK2質粒48 h后,加細胞裂解液冰上裂解30 min,提取蛋白,BCA蛋白定量后,加入5×蛋白上樣緩沖液,沸水煮沸10 min,Western blot檢測蛋白表達情況。
1.2.7His-GRK2重組蛋白的純化和鑒定 將GRK2真核重組質粒轉染至HEK 293T細胞,培養48 h后,收集細胞,加入蛋白裂解液,通過細胞破碎儀破碎細胞,4 ℃條件下,13 400 r/min離心15 min,收集蛋白上清液,將蛋白上清液與6FF瓊脂糖凝膠微珠,4 ℃搖床孵育2 h,離心收集微珠,4 ℃條件下,蛋白洗液(含20 mmol/L 咪唑)洗滌3次,30 min/次,離心收集微珠,4 ℃條件下,蛋白洗脫液(含300 mmol/L 咪唑)洗滌3次,30 min/次,分別收集3次洗脫蛋白(第1次洗脫蛋白、第2次洗脫蛋白、第3次洗脫蛋白),將3次洗脫蛋白加入5 ml超濾管(截留蛋白30~50 ku)離心,收集蛋白,加入蛋白置換液(低鹽蛋白緩沖液)于5 ml超濾管,進行離心,置換蛋白緩沖液,得到脫鹽后的蛋白,通過考馬斯亮藍染色和Western blot檢測3次洗脫蛋白和脫鹽后蛋白的表達。
1.2.8His pull down 檢測GRK2蛋白的活性 本研究前期已成功構建 pIRES-EGFP-EP4 質粒,將與GRK2真核重組質粒分別轉染HEK 293T細胞,收集轉染后的細胞,加入蛋白裂解液,通過細胞破碎儀破碎細胞后,4 ℃條件下,13 400 r/min離心15 min,將帶有His標簽的GRK2與6FF瓊脂糖凝膠微珠,4 ℃搖床孵育2 h,離心收集微珠,4 ℃條件下,蛋白洗液(含20 mmol/L 咪唑)洗滌3次,30 min/次,離心收集微珠。將過表達pIRES-EGFP-EP4的蛋白與處理的微珠4 ℃搖床孵育12~16 h。離心收集微珠,加入20~30 μl的蛋白上樣緩沖液,沸水煮10 min,Western blot檢測GRK2與EP4的共表達。
1.3 統計學處理采用GraphPad Prism 8 軟件進行統計學處理,實驗數據以均數±標準誤(SEM)表示。采用t檢驗比較兩組間差異,P<0.05為差異有統計學意義。
2 結果
2.1 pcDNA3.1-EGFP-His-GRK2重組質粒的構建利用設計的引物以pIRES-EGFP-GRK2基因為模板,利用合成引物進行PCR擴增 His-GRK2目的基因全長,進行1%瓊脂糖電泳。凝膠成像系統檢測發現,線性His-GRK2目的片段位置在2 067 bp左右(圖1A)。將PCR產物進行1%瓊脂糖電泳,切膠回收產物。將PCR回收產物與pcDNA3.1-EGFP空載體通過Hind Ⅲ和BamH Ⅰ進行雙酶切,酶切產物通過T4 DNA連接酶進行連接,構建pcDNA3.1-EGFP-His-GRK2質粒,雙酶切鑒定結果表明,在 2 067 bp左右出現線性His-GRK2目的片段,在6 154 bp左右出現線性pcDNA3.1-EGFP目的片段,提示pcDNA3.1-EGFP-His-GRK2質粒成功構建(圖1B)。同時隨機挑選1個酶切鑒定陽性的質粒進行測序,并對測序結果進行拼接,利用https://blast.ncbi.nlm.nih.gov網址進行測序比對,發現 GRK2 standard Seq 與測序結果一致,pcDNA3.1-EGFP-His-GRK2重組質粒未發生突變,測序序列對比未出現異常,說明 pcDNA3.1-EGFP-His-GRK2 重組質粒成功構建(圖1C)。

圖1 人源GRK2基因PCR擴增、酶切鑒定和測序
2.2 pcDNA3.1-EGFP-His-GRK2蛋白成功表達HEK 293 T細胞以2×106個/孔細胞密度接種于6孔板中,轉染pcDNA3.1-EGFP-ctr和pcDNA3.1-EGFP-His-GRK2重組質粒,培養 48 h后,在倒置熒光顯微鏡下觀察轉染效果,可見綠色熒光蛋白(EGFP標簽)的表達,pcDNA3.1-EGFP-His-GRK2質粒轉染效果達到60%以上(圖2A)。收集轉染后的細胞,細胞裂解液冰上裂解30 min,Western blot用于檢測GRK2重組蛋白的表達,結果表明pcDNA3.1-EGFP-His-GRK2在HEK 293 T細胞中過表達(t=6.433,P=0.003)(圖2B)。
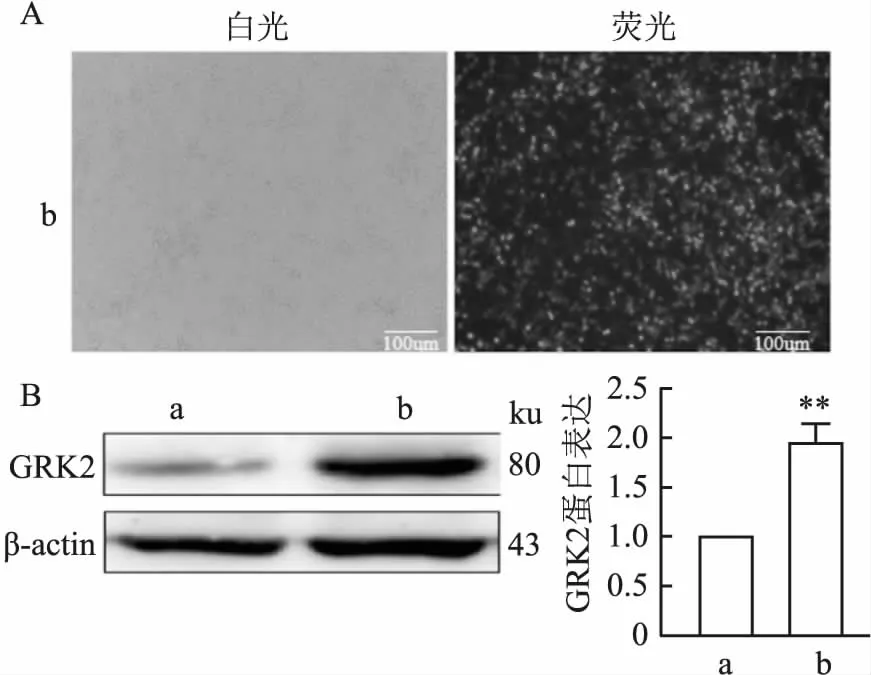
圖2 Western blot檢測GRK2蛋白表達
2.3 表達和純化獲得His-GRK2重組蛋白HEK 293T細胞轉染 pcDNA3.1-EGFP-His-GRK2 質粒后,裂解蛋白,加入 6FF 瓊脂糖凝膠微珠孵育后。加入1 ml蛋白洗脫液(高鹽),洗脫蛋白進行超濾,得到 500 μl 蛋白溶液。考馬斯亮藍染色檢測GRK2蛋白純化效果見圖 3A,Western blot 檢測 GRK2 蛋白表達結果見圖3B。結果表明,考馬斯亮藍染色和Western blot檢測均可以在80 ku左右檢測GRK2陽性蛋白表達。

圖3 考馬斯亮藍染色和Western blot分析GRK2蛋白的表達和純化
2.4 GRK2結合EP4HEK 293T細胞轉染pcDNA3.1-EGFP-His-GRK2質粒,同時轉染pIRES-EGFP-ctr和pIRES-EGFP-EP4質粒,裂解蛋白后,與結合His蛋白的鎳珠共同孵育,處理后的蛋白樣品沸水煮10 min,Western blot檢測EP4蛋白的表達。結果表明,過表達EP4可以促進GRK2與EP4的結合(t=13.5,P=0.000 2)(圖 4)。

圖4 His pull down 檢測GRK2與EP4的結合能力
3 討論
GRK2由689個氨基酸(amino acid, aa)組成,具有3個重要的結構域:氨基端(1-185 aa)、激酶活性區(186-513 aa)和羧基端(514-689 aa)[6]。GRK2的氨基端具有一個G 蛋白信號同源域調節(regulator of G protein signaling homology, RH)區,通過結合Gβγ等蛋白發揮轉膜功能[7]。GRK2激酶活性的改變是GRK2發揮多種生理和病理功能的重要條件,有研究[8]表明220位的賴氨酸(Lys, K)是決定GRK2激酶活性的關鍵氨基酸位點,Lys220突變為精氨酸(Arg, R)可使GRK2激酶活性喪失。GRK2的羧基端具有RH區和 Pleckstrin同源(Pleckstrin homology, PH)區,有利于其亞細胞定位和激動劑依賴的轉膜,可以與游離Gβγ亞單位、細胞外信號調節激酶(extracellular signal-regulated kinase, ERK)等蛋白結合[9-10]。目前已有許多研究表明GRK2表達和活性的改變與疾病的發生相關,如GRK2表達和活性增加在高血壓、心衰以及RA的發生和發展中起著重要作用,而GRK2的表達和活性的降低與肝癌、多發性硬化癥等疾病的發生密切相關[11]。這些研究結果提示,GRK2的活性維持在一個平衡狀態是研究GRK2活性調節劑的主要方向。現已有多種GRK2活性調節劑被嘗試用于治療心力衰竭、RA等疾病,其中帕羅西汀、Gallein、GSK180736A等已經進入動物試驗階段[12-13]。
本實驗在GRK2的羧基末端引入了6×His (CATCATCACCATCACCAT)的短肽是蛋白純化常用的標簽。目前常用于蛋白純化的標簽蛋白包括His、Myc、Flag、GST等標簽,但Myc重組蛋白的低pH洗脫條件往往會降低蛋白質的活力,所以較少用于純化蛋白,Flag重組蛋白的純化成本較高,GST標簽純化成本低但增溶能力一般。His 標簽相較于Myc、Flag、GST等標簽具有以下優勢:① 標簽蛋白非常小,不會影響目的蛋白質本身的結構;② 不會影響目的蛋白自身的活性;③ 便于采用固化金屬離子親和層析;④ 操作方便,純化蛋白成本低。本實驗選擇pcDNA3.1-EGFP真核表達載體構建His-GRK2真核表達重組質粒,這與原核表達體系相比,雖然不能一次性獲得大量的蛋白,但具有以下優勢:外源性真核表達特異性強,具有翻譯后加工修飾功能,使其結構、糖基化方式更接近天然蛋白,具有高活性。
獲得有活性的GRK2蛋白是進一步研究其功能和作用機制的前提。本研究通過設計引物,利用PCR擴增技術得到帶有His標簽的GRK2基因,并采用分子克隆手段將GRK2基因連接到pcDNA3.1-EGFP真核表達載體上,成功構建pcDNA3.1-EGFP-His-GRK2真核表達質粒,通過酶切鑒定和測序結果證明pcDNA3.1-EGFP-His-GRK2構建成功。再利用瞬時轉染技術將其轉染入HEK 293T中,通過Western blot及考馬斯亮藍染色驗證GRK2在真核細胞293T中正確表達。通過His pull down 的方法鑒定出GRK2與EP4的相互作用,具有生物學活性,為后續研究GRK2的作用機制奠定了基礎。
