FtsZ抑制劑作為新型抗菌分子的研究進展
2023-02-27 11:47:42郭澤莉吳文玉
廣州醫藥 2023年2期
郭 琦 郭澤莉 吳文玉 孫 寧
1 中山大學中山眼科中心,眼科學國家重點實驗室,廣東省眼科視覺科學重點實驗室(廣州 510060)
2 廣州市第一人民醫院/華南理工大學附屬第二醫院(廣州 510180)
細菌的耐藥性問題被認為是當今所面臨的最大科學挑戰之一。例如耐甲氧西林金黃色葡萄球菌(methicillin-resistant Staphylococcus aureus,MRSA)、多重耐藥金黃色葡萄球菌、耐萬古霉素腸球菌和多粘菌素耐藥大腸桿菌等新型耐藥菌的出現,嚴重降低了臨床上常用的抗菌藥物的療效,曾經被普遍用來治療常見的腸道與尿道感染、肺炎、新生兒感染以及淋病的抗菌藥物,因為細菌對抗生素產生的超強抵抗力,已經逐步失去抑制耐藥菌生長繁殖的能力,進而導致耐藥菌感染患者的死亡率逐年增加,威脅人類的健康。另一個令人擔憂的問題是新型抗生素的研發相對滯后。美國食品藥品監督管理局批準上市的新抗生素藥物近年來一直在減少, 從1962年起到現在, 只有3個新型抗生素藥物(利奈唑胺、達托霉素和Retapamulin) 獲批[1]。預計到2050年,全球因抗生素耐藥性感染而死亡的人數估計將從2014年的70 萬上升至1 000萬,由此累計的醫療和生產力成本將達到100萬億美元[2]。鑒于耐藥細菌對人類健康的威脅和臨床用藥選擇范圍的相對縮小,研發新作用機制和新化學結構的新一代抗菌藥物,有效地殺死耐藥細菌是解決這一全球性危機的重要舉措。
近幾年來,細菌的細胞分裂發生機制成為科研單位和藥物企業研發新型抗生素的新方向。在目前已明確有效的靶點中,細胞分裂溫度敏感突變體Z(filamenting temperature-sensitive mutant Z, FtsZ)是最具開發潛力和研究最為深入詳細的作用靶點之一[3]。有關FtsZ蛋白的結構與功能研究揭示,它存在于絕大多數細菌中,具有高度的保守性,是介導細菌細胞分裂的關鍵蛋白。當其生物活性受到抑制時,細菌的細胞分裂將受到影響,進而導致細菌的生長繁殖被抑制。2008 年,英國Prolysis制藥公司在Science上發表了能有效抑制MRSA生長繁殖的FtsZ蛋白抑制劑PC190723[4]。2012年,Merck制藥公司在ScienceTranslationalMedicine雜志上詳細闡述了該化合物的作用機制,并揭示了其與甲氧苯青霉素聯合對抗MRSA的協同作用。目前,基于這一化合物的衍生物仍在進行前期的臨床研究,有望開發成為新的抗菌藥物。隨著FtsZ蛋白在細菌分裂中的功能和作用機制的深入研究,越來越多生物醫藥研究者和藥物開發公司,開始針對FtsZ蛋白著力開發新一代的抗菌藥物。
1 FtsZ蛋白的結構和功能
FtsZ蛋白是細菌分裂增殖過程所必需的功能性蛋白,它存在于絕大部分原核生物中[5]。FtsZ蛋白的分子量約為40.3 kDa,許多FtsZ蛋白的高分辨的X單晶衍射晶體結構揭示了其具有兩個結構域組成的保守結構(圖1)[5]。N 端為核苷結合區(GTP結合位點),而C端包含一個柔性很強的環(T7 loop),這兩個結構域由結構中央的H7 螺旋分隔開,由H5螺旋連接,形成小分子結合口袋(主要氨基酸有G193、G196、N263)。研究表明,FtsZ 以頭接尾的模式聚合,一個單體的T7環會插入另一個單體的GTP結合位點,此時催化殘基得以靠近γ-磷酸,從而催化GTP的水解[6]。
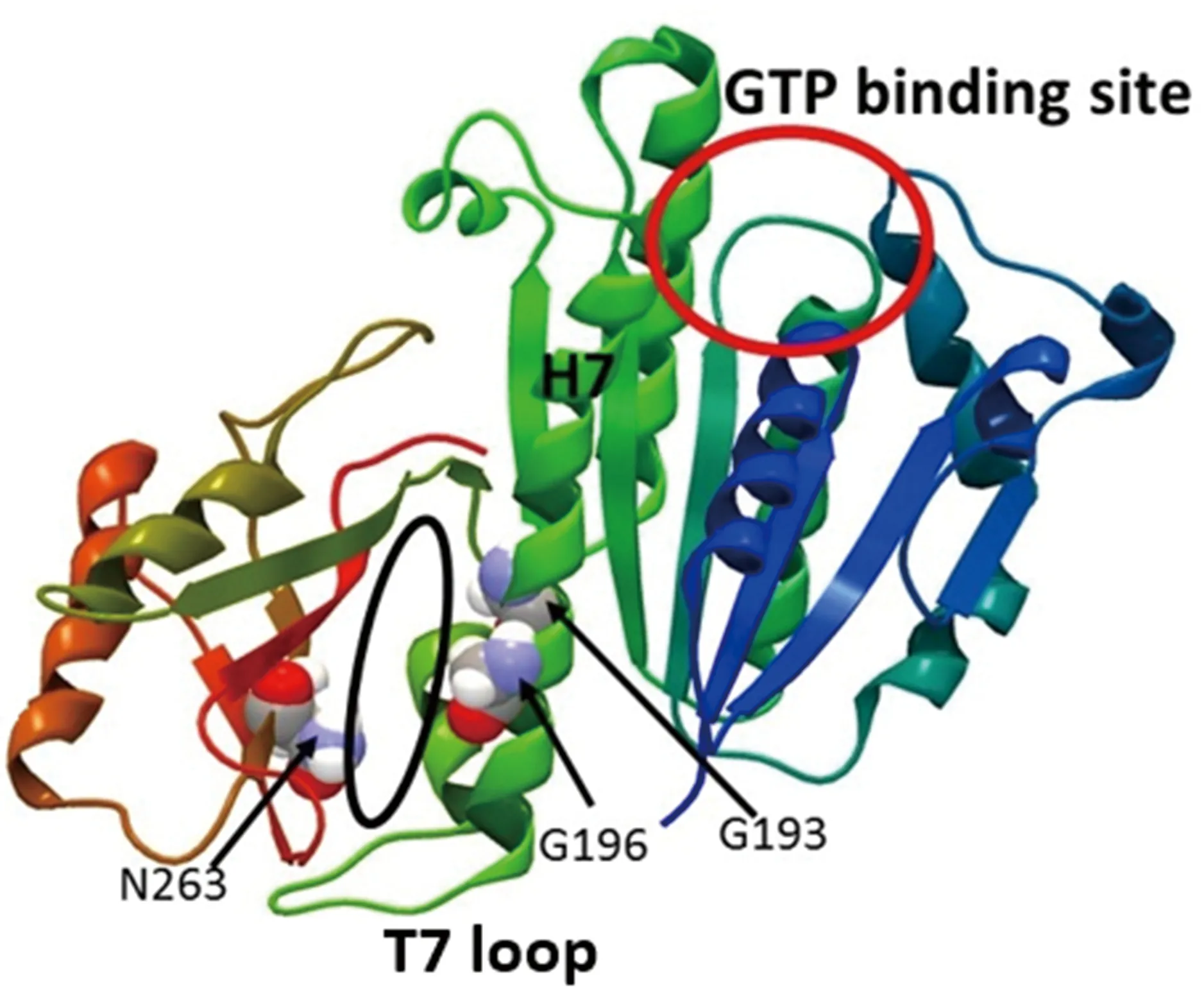
圖1 FtsZ蛋白結構示意圖
FtsZ蛋白的結構和功能角色與真核細胞的微管蛋白類似,具有一定的同源性,但是,微管蛋白的C端結構域更大。通過比較FtsZ蛋白與α-和β-微管蛋白的二維晶體結構,可以看出FtsZ蛋白和β-微管蛋白的GTP結合位點和C端在結構上有很強的相似性,所以FtsZ蛋白也被認為是微管蛋白的類似物。但兩者在蛋白質氨基酸序列上只有7%的相似性,在核苷酸結合方式上也有很大不同。此外,微管蛋白是由α和β異二聚體組成,具有明顯的極性,而FtsZ蛋白聚合物只由一個個FtsZ單體構成[3]。這些結構上的差異,為開發靶向FtsZ蛋白卻又對微管蛋白無影響的藥物提供了有效的條件。
細菌細胞的分裂是一個由多蛋白參與調控的復雜生物學過程,FtsZ蛋白在這一過程中起著至關重要的作用。在細菌啟動細胞分裂時,FtsZ蛋白是第一個遷移到分裂位點的蛋白,通過GTP供能條件下首尾相接形成單條直鏈原絲蛋白,然后直鏈原絲蛋白平行相接形成FtsZ蛋白束,最后在細胞中部圍繞細胞形成環狀結構,相關領域的學者稱這一過程為FtsZ蛋白的動態聚合,稱這個環狀結構為Z環。Z環形成后,會進一步招募其他細胞分裂蛋白(FtsA、ZipA、ZapA等),通過蛋白-蛋白直接或間接作用形成細胞間隔膜。在FtsZ蛋白水解GTP提供的能量驅動下,Z環不斷向內收縮,母體細胞進而一分為二成為兩個子細胞(圖2)[3,7]。這些發現為進一步研究FtsZ蛋白動態聚合的詳細機制和基于蛋白結構的藥物設計提供了可能。FtsZ 蛋白的動態聚合對Z 環的形成起著至關重要的調控作用。當FtsZ 蛋白的動態聚合或GTP 水解活性受到抑制時,Z 環將無法形成或無法保留其完整功能,即使細菌的DNA 復制和蛋白質合成等功能均運轉正常,但細菌在此狀態下只能形成狹長絲狀(桿狀細菌)或肥大球狀(球狀細菌)的形態,無法進行增殖分裂,從而導致細菌的死亡[3]。
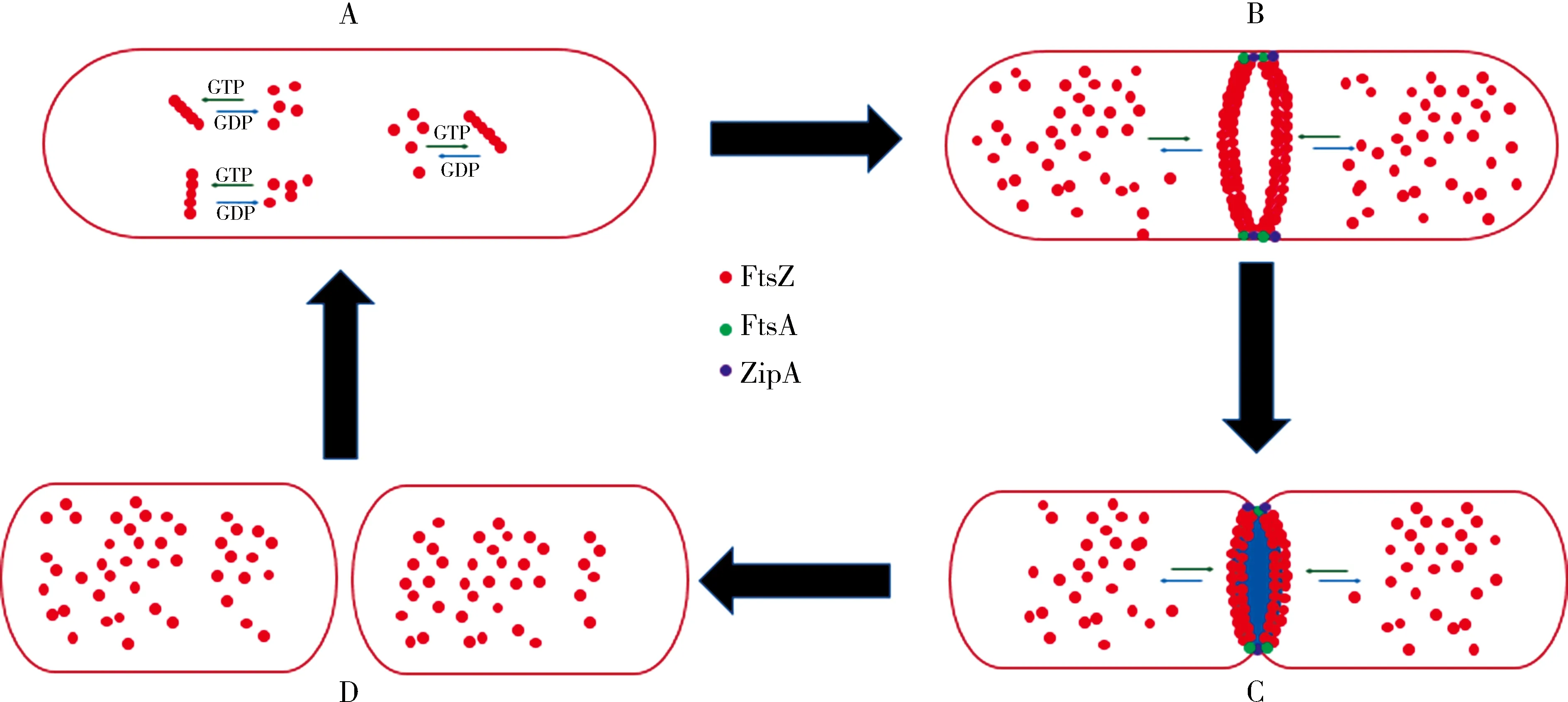
圖2 細菌分裂增殖示意圖
2 FtsZ蛋白抑制劑
隨著對FtsZ蛋白的研究不斷深入,醫藥研究者對其生物學功能的認識不斷加深,開發靶向FtsZ蛋白的抗菌先導化合物也越來越受到研究者們的重視。目前,已有一些國內外研究團隊在開展FtsZ蛋白抑制劑的研究,從現有的研究報道可以看出,當前FtsZ蛋白抑制劑主要包括天然產物及其衍生物和合成類抑制劑。
2.1 天然產物及其衍生物
2.1.1 黃連素及其衍生物 黃連素(圖3)是一種常見的天然生物堿, 2008年,Dasgupta團隊的研究發現黃連素可能是通過抑制FtsZ蛋白的GTP酶活性(IC50=16 μmol)和動態聚合(IC50=10 μmol),起到抑制細菌生長的活性。分子模擬研究表明,黃連素的結合部位與大腸桿菌FtsZ蛋白中的GTP結合口袋相重疊[8]。Sun等人[9]通過基于FtsZ蛋白結構的設計方法,合成了一些9-苯氧基烷基黃連素的衍生物,分子對接的結果顯示,這些衍生物的結合位點位于金黃色葡萄球菌FtsZ蛋白的C-末端結構域間隙。這類化合物具有較強的抗革蘭氏陽性菌的活性,對MRSA和VRE的最低抑菌濃度(minimum inhibitory concentration,MIC)值分別為2~8 μg/mL和4~16 μg/mL。此外,這些化合物對革蘭氏陰性桿菌也表現出中等的抗菌活性,如大腸桿菌,其MIC值為32~128 μg/mL。在這些衍生物中,化合物2(圖3)具有最強的抗菌活性,對MRSA和VRE的MIC值分別為2和4 μg/mL。此外,它還能抑制FtsZ蛋白的聚合和GTP酶的水解活性(IC50=38 μg/mL)。透射電子顯微鏡圖像顯示,該化合物能顯著減小FtsZ聚合物的尺寸和厚度。
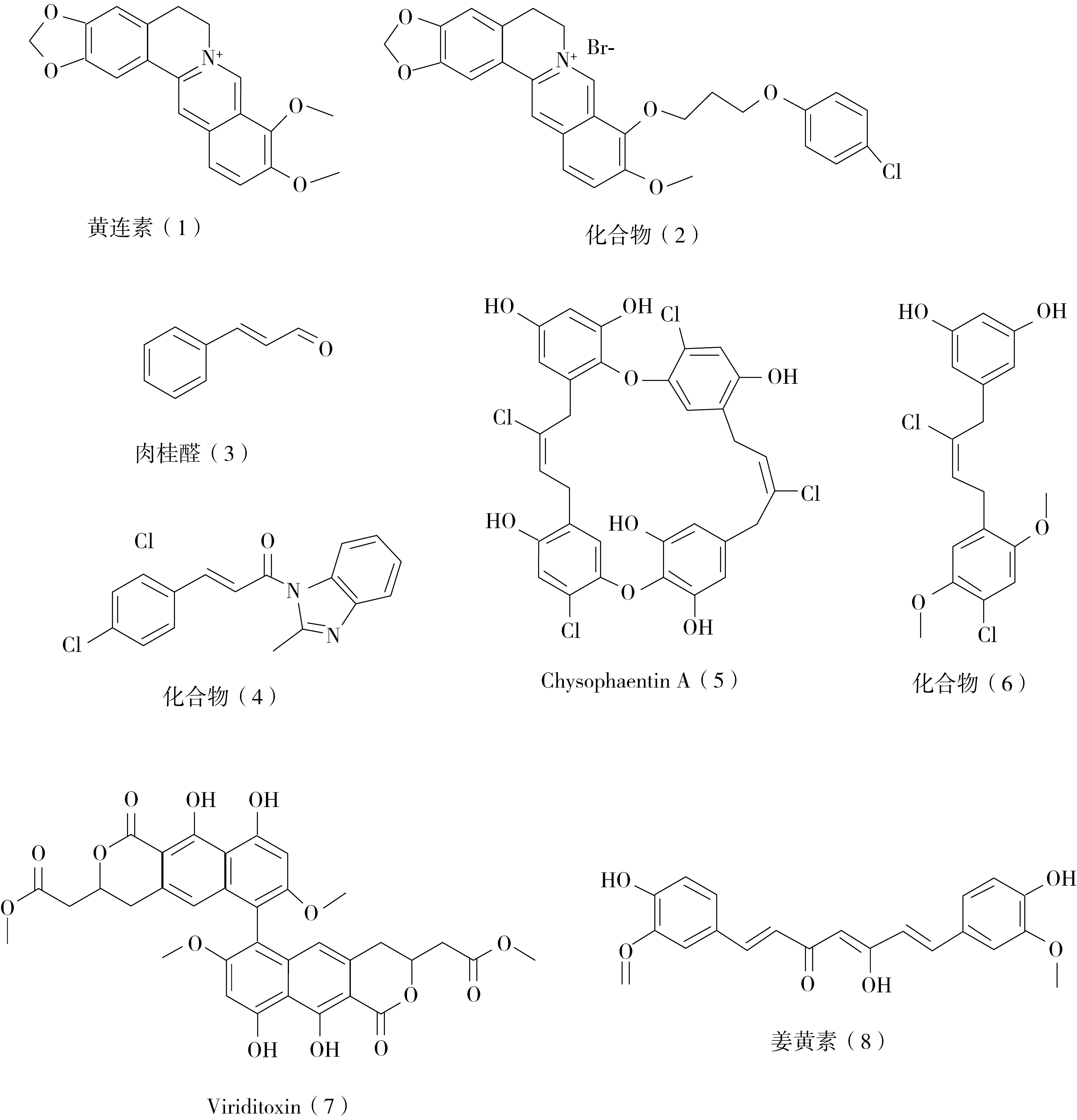
圖3 部分天然產物及其衍生物來源的FtsZ抑制劑化學結構
2.1.2 肉桂醛及其衍生物 肉桂醛(圖3)是一種從肉桂樹皮中提取的天然芳香化合物。它能夠抑制大腸桿菌FtsZ蛋白的GTP酶活性和動態聚合聚合,IC50值分別為5.81 μmol和6.86 μmol。分子模擬結果和STD-核磁共振的檢測結果表明,肉桂醛結合在FtsZ蛋白的C-末端區域T7環周圍的結合口袋中[10]。基于這個結果,Ma和他的同事合成了一系列肉桂醛衍生物,并測試了它們對革蘭氏陽性和陰性細菌的抗菌活性。其中部分化合物能夠抑制金黃色葡萄球菌ATCC25923的細胞分裂,作用濃度為0.25~4 μg/mL。其中含有2-甲基苯并咪唑和2,4-二氯苯基基團的化合物具有最強的活性 (化合物4)。生物活性測試的結果表明,這些化合物能夠劑量依賴地抑制金黃色葡萄球菌FtsZ的聚合和GTP酶活性[11]。
2.1.3 Chrysophaentins Chrysophaentins A-H是Bewley等人從海洋大黃藻中分離得到的新的抗菌天然產物。這類化合物對幾種革蘭氏陽性細菌株都有很強的抗菌作用。其中,Chrysophaentins A(圖3)對多種耐藥菌均顯示出很強的抗菌活性。例如,該化合物對MRSA和肺炎鏈球菌的MIC值分別為1.5 μg/mL和2.9 μg/mL。此外,它對大腸桿菌FtsZ蛋白的GTP酶有抑制作用,其IC50值為6.7 μg/mL,并能抑制FtsZ的動態聚合。進一步的GTPγS競爭性STD-核磁共振實驗證明,該化合物能競爭性地結合在FtsZ蛋白的GTP結合位點[12]。基于這些結果,Bewley等人[13]合成改造了這類化合物,在GTP酶活性測定中,新化合物6對金黃色葡萄球菌FtsZ和大腸桿菌FtsZ的GTP酶活性均有抑制作用,IC50值分別為(38±9) μmol和(37±7)μmol。熒光靶向性實驗結果表明,化合物6的結合位點也是FtsZ蛋白的核苷酸結合位點。
2.1.4 Viriditoxin Viriditoxin(圖3)來源于綠垂曲霉。它能有效抑制大腸桿菌FtsZ蛋白的GTP酶活性和動態聚合,IC50值分別為7.0 μg/mL和8.2 μg/mL。對細菌形態的測定結果表明,該化合物能引起枯草桿菌的延長。此外,該化合物對許多耐藥革蘭氏陽性病原菌,如金黃色葡萄球菌(MIC值為4~8 μg/mL)、糞腸球菌和屎腸球菌(MIC值為2~16 μg/mL)均顯示出較強的抗菌作用。此外,在細菌細胞中誘導FtsZ蛋白的高表達可以引起該化合物MIC值的升高,這表明在細菌菌株中,Viriditoxin具有靶向FtsZ的體內靶向性[14]。
除了以上所述的天然產物類抑制劑,還有姜黃素(圖3)、秋水仙堿、白藜蘆醇、紫杉烷及其衍生物等,這些天然產物都能夠作用于FtsZ 蛋白,對FtsZ 抑制劑的研究工作有一定的指導意義[3]。
2.2 合成類FtsZ蛋白抑制劑
2.2.1 苯甲酰胺類衍生物 3-甲氧基苯甲酰胺(3-mercaptobutanoic acid,3-MBA)(圖4)因為能顯著抑制枯草芽孢桿菌的細菌分裂而被發現能在細菌體內靶向FtsZ蛋白[15]。鑒于3-MBA的低分子質量、良好的水溶性等優點,Czaplewski等人在500多個3-MBA衍生物的分子庫中發現了化合物PC190723(10)。PC190723對枯草芽孢桿菌、MRSA、多藥耐藥金黃色葡萄球菌等革蘭氏陽性菌均有很強的抗菌活性。MIC值低至1 μg/mL。PC190723對金黃色葡萄球菌FtsZ蛋白的GTP酶活性具有劑量依賴性抑制作用,IC50值為55 ng/mL。細胞形態研究發現,經PC190723處理的桿狀枯草芽孢桿菌的長度顯著變長,而球形金黃色葡萄球菌的體積顯著增大。表明細菌的細胞分裂受到明顯的抑制。此外,研究還發現PC190723能干擾枯草芽孢桿菌細胞內Z環的形成。分子對接研究表明,PC190723結合在C末端和H7螺旋之間的結構域間隙上[4]。Andreu等人[16]發現,PC190723通過誘導和穩定FtsZ聚合物形成直束和條帶,抑制了枯草芽孢桿菌FtsZ蛋白的GTP酶活性。Lumb等人通過FtsZ蛋白與PC190723的共晶體分析發現,PC190723結合在FtsZ蛋白的C末端結構域,并能穩定FtsZ的聚合形態,這一發現也驗證Andreu等人的結論[17]。在后續的動物實驗中,PC190723的藥代動力學特征不甚理想,限制了它的進一步應用。Kate等人[18]在該化合物的基礎上進一步改造出具有更好的代謝穩定性、藥代動力學特性和抗菌功效的類似物TXA436、TXA707 和TXA709 (圖4)。目前TXA709 正處于臨床試驗階段,已經完成了首次人體Ⅰ期臨床試驗,并在2016 年9月被美國食品和藥物管理局指定為合格傳染病產品;后續將加入額外的I 期人類安全臨床試驗,與臨床抗生素聯用進行耐甲氧西林金黃色葡萄球菌感染的治療。
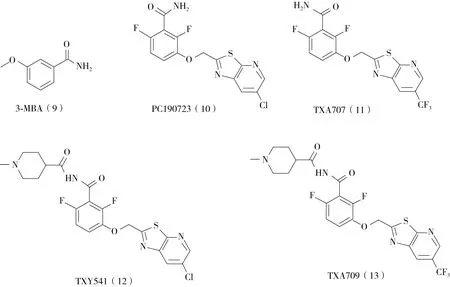
圖4 部分苯甲酰胺類FtsZ抑制劑化學結構
2.2.2 芳烴二醇-雙沒食子酸衍生物 Ruiz-Avila等人通過小分子與枯草芽孢桿菌FtsZ蛋白 GTP結合位點的分子對接方法,從化合物庫中篩選出一系列與GTP競爭的FtsZ蛋白小分子抑制劑,并通過與MANT-GTP的競爭性實驗,進一步驗證這些小分子的靶向性。實驗結果表明,UCM05(14)、UCM44(15)和UCM53(16)對細菌生長有明顯的抑制作用,MIC值分別為100 μmol、25 μmol和13 μmol。此外,這三種化合物都能有效地誘導枯草芽孢桿菌細胞伸長并干擾Z環的形成[19]。根據這些化合物的結構,他們設計合成了一些小分子抑制劑,并評價了它們對細菌和FtsZ的抑制活性。最有效的化合物17(圖5)顯示出與FtsZ蛋白極強的結合親和力(Kd=0.5 μmol)和對MRSA的抗菌活性(MIC值為7 μmol)。大多數芳烴二醇在100 μmol時對微管蛋白聚合幾乎沒有抑制作用,表明對該類化合物對FtsZ蛋白具有很高的選擇性[20]。
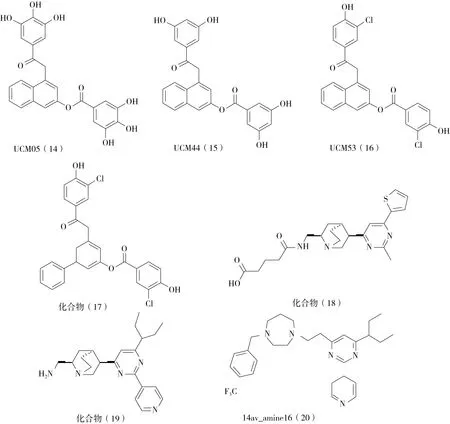
圖5 芳烴二醇-雙沒食子酸衍生物和嘧啶-奎寧類衍生物及其類似物的化學結構
2.2.3 嘧啶-奎寧類衍生物及其類似物 為了識別針對FtsZ的GTP結合位點的命中化合物,Wong等人[21]基于FtsZ蛋白的結構和功能特征,利用計算機藥物輔助篩選軟件,對20 000多種化合物進行了基于結構的虛擬篩選,并對篩選出的10個化合物進行了體外GTP酶抑制作用和對病原菌的抑菌活性測試。其中,化合物18(圖5)對FtsZ蛋白(IC50=317 μmol)表現出中等的GTP酶抑制活性,對金黃色葡萄球菌和大腸桿菌的MIC值分別為449 μmol和897 μmol,其余9個化合物均無活性。在該化合物的基礎上,他們擴大對結構類似物的篩選,成功發現了化合物19,其在生物學試驗中顯示出顯著的活性改善。該化合物的GTP酶抑制活性(IC50=37.5 μmol)和對金黃色葡萄球菌(MIC=24.6 μmol)和大腸埃希菌(MIC=49.6 μmol)的抗菌活性均比27提高10倍以上[21]。此外,該化合物能夠選擇性地抑制FtsZ蛋白的活性,而不影響微管蛋白。抗菌實驗還發現,該化合物能夠恢復MRSA(ATCCBAA- 41)對β-內酰胺類抗生素的敏感性[22]。以該化合物為模版,Wong等人成功地合成了99個胺基連接的2,4,6-三取代嘧啶衍生物,并研究了它們的生物活性。對金黃色葡萄球菌的藥敏試驗結果表明,這些化合物具有較強的抗葡萄球菌活性(MIC值為3~8 μg/mL)。化合物14av_amine16(20)的STD-核磁共振、光散射和GTP酶活性實驗表明,它通過抑制FtsZ的聚合和GTP酶活性起到抑制細菌分裂的作用。此外,枯草芽孢桿菌Z環和延長的細菌表型都揭示了該分子在細菌體內的FtsZ靶向特性。此外,14av_amine16不易引起金黃色葡萄球菌耐藥株的產生,且對大額螟蟲幼蟲幾乎沒有毒副作用[23]。
2.2.4 喹啉衍生物 之前的研究表明,在血根堿中加入疏水取代基能夠顯著提高抗菌活性。LaVoie等人[24]進一步簡化了血根堿的化學骨架,設計合成出一系列3-苯基異喹啉衍生物,用于抗菌活性評價。結果表明, 3′-位取代基的親脂性與抗菌效果有關。化合物21(圖6)對耐甲氧西林金黃色葡萄球菌和沙門氏菌的MIC值分別為1 μg/mL和4 μg/mL。熒光滴定法的結果表明,該類化合物能與FtsZ蛋白相互作用,它們的離解常數范圍為1~10 μM。此外,這些化合物對哺乳動物微管蛋白幾乎沒有影響,對哺乳動物細胞的毒性也很小。在上述喹啉類化合物研究的基礎上,Sun等人對喹啉衍生物進行了研究。研究了N-甲基苯并吲哚并[3,2-b]喹啉和N-甲基苯并呋喃并[3,2-b]喹啉衍生物以及噻唑橙-喹啉衍生物22(圖6)[25- 26]的抗菌活性。結果表明,這些化合物對MRSA和VRE等細菌具有較強的抗菌活性。生物化學研究結果表明,這些化合物通過抑制FtsZ蛋白的GTP酶活性,破壞了FtsZ蛋白的動態組裝和Z環的形成,起到抑制細菌分裂的作用[25- 26]。
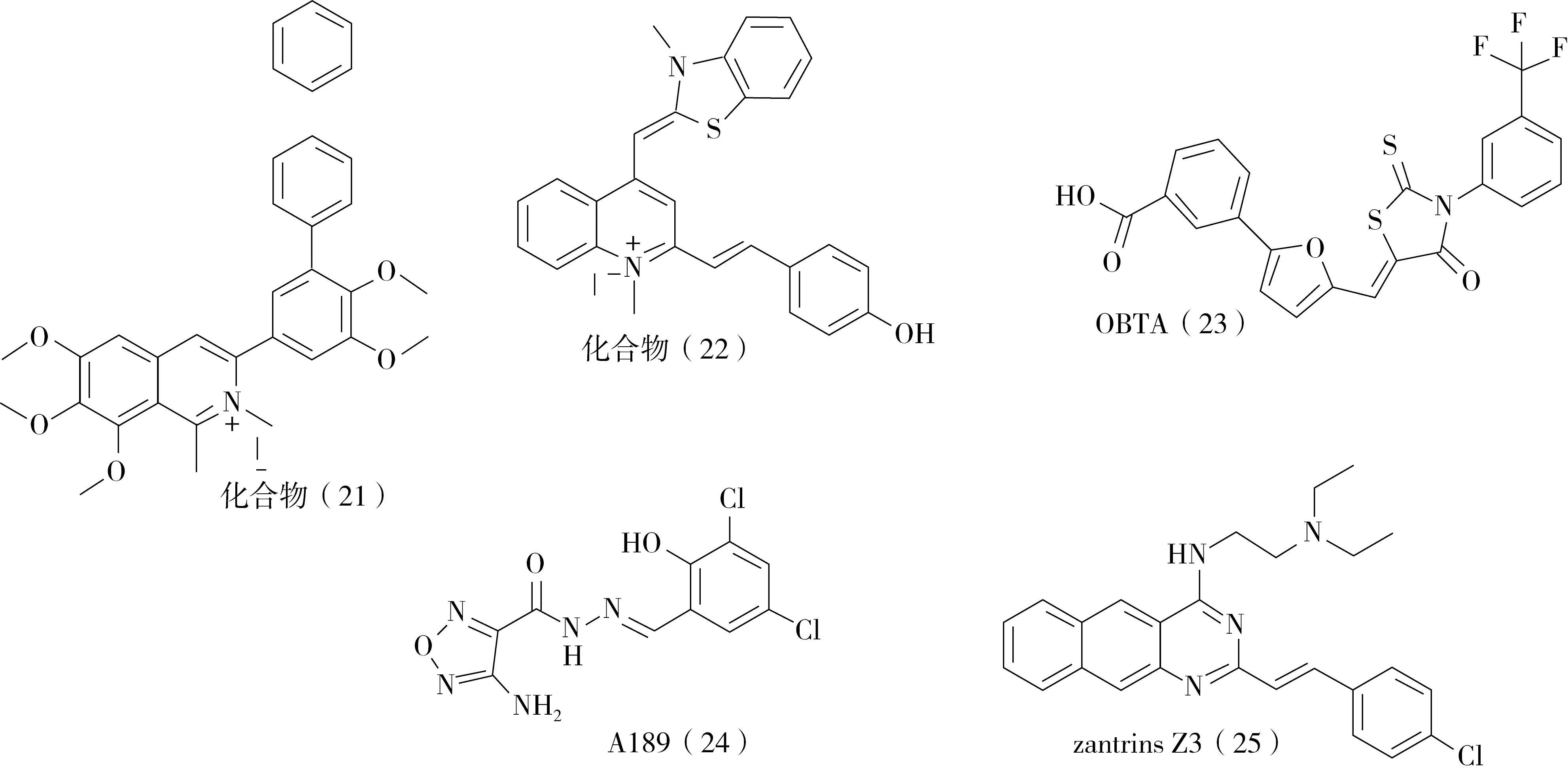
圖6 部分合成類FtsZ抑制劑的化學結構
2.2.5 其他結構類型的FtsZ蛋白抑制劑 除了上述的合成類FtsZ 蛋白抑制劑,研究人員還發現OTBA、A189、zantrins(圖6) 系列等合成類小分子能通過影響FtsZ蛋白起到抗菌效果[3]。OTBA是在81 個化合物中篩選出來的結構,它可以促進FtsZ 原絲組裝,同時抑制其GTP 酶活性。氨基呋喃類的A189是通過無核細胞藍色測定鑒定出來的,該實驗能夠有目的性地尋找抑制FtsZ 的GTP 酶活性的抑制劑,后續的研究表明A189 能抑制FtsZ 蛋白聚合,并且對金黃色葡萄球菌和大腸桿菌有較好的抑制作用。另外zantrin Z3通過抑制FtsZ 蛋白的GTP 酶活性和穩定FtsZ 原絲組裝,進而使細菌細胞絲狀化。
2.3 多肽類FtsZ蛋白抑制劑
在多細胞生物體中發現的一種由37個氨基酸殘基組成的抗菌肽CRAMP,具有一定的抗菌活性,且能夠誘導細菌細胞伸長,表明它可以抑制細菌胞質分裂。Ray等人[27]在此基礎上,發現截短的CRAMP多肽片段(GEKLKKIGQKIKNFFQKL,16- 33)具有不錯的抗菌活性和FtsZ抑制作用。該多肽在20 μmol和50 μmol濃度下均能完全抑制枯草芽孢桿菌和大腸桿菌的生長。FtsZ的光散射實驗表明,該多肽對FtsZ的聚合有劑量依賴性的抑制作用,并且不影響微管蛋白聚合。此外,該肽還降低FtsZ蛋白的GTP水解活性,抑制枯草芽孢桿菌的細胞分裂和Z-環的形成,導致細菌細胞伸長。Pieraccini等人[28]使用計算丙氨酸掃描技術(CAS),識別FtsZ亞基之間蛋白質-蛋白質相互作用界面上的熱點,設計合成了與FtsZ亞序列相對應的八肽,該八肽能干擾FtsZ蛋白自組裝。隨后,他們通過兩種不同的環化方案對該多肽進行修飾,以限制其螺旋幾何形狀。這些環肽在FtsZ動態聚合的初始階段具有顯著的抑制作用。
3 結論與展望
目前的研究結果表明,FtsZ 蛋白是一個極具發展潛力的抗菌藥物新靶點。主要表現在:(1)FtsZ蛋白是細菌生長繁殖的關鍵性蛋白。它在細菌細胞分裂和生長繁殖中必不可少,在調控細菌分裂增殖過程中起核心作用[3]。(2)FtsZ蛋白具有高度的保守性和相似性。絕大部分細菌中都發現含有FtsZ蛋白,包括各種耐藥菌和肺結核桿菌等臨床致病菌。在所有細菌分裂蛋白中,FtsZ蛋白的保守性和相似性最高,不易產生變異。因此,FtsZ蛋白抑制劑具有廣譜抗菌活性,并且不易產生細菌耐藥性[3]。(3)FtsZ蛋白只在細菌中特異性存在,雖然FtsZ 蛋白與人類的微管蛋白有類似的GTP 水解活性,但是兩者的蛋白質氨基酸序列只有不到20%的相似度,在生物學結構和功能活性等方面存在很大的區別[3]。從已報道抑制劑情況可知,我們可以根據FtsZ蛋白的結構和生物學特性,設計靶向FtsZ蛋白的抑制劑,通過干擾FtsZ蛋白的動態聚合和GTP水解活性,高效專一地抑制細菌細胞的分裂,進而達到抑菌或滅菌的效果。
目前,雖然在FtsZ蛋白抑制劑上取得了一定的研究進展,但尚未有靶向FtsZ蛋白的抗菌藥物進入二期臨床試驗。其原因可能是這些抑制劑雖然具有較好的體外抗菌活性,但仍存在毒副作用或體內分布代謝效果不理想等問題。另一方面,對FtsZ蛋白抑制劑的作用機制研究不夠深入,也阻礙了靶向FtsZ蛋白的抗菌分子的結構改造和進一步開發。目前,大部分已知的抑制劑都只針對FtsZ蛋白進行了簡單的機制探討,鮮有在細菌體內深入探討其作用機制,也沒有深入研究化合物與FtsZ蛋白的具體結合模式。因此,仍需在不斷擴大FtsZ蛋白抑制劑分子庫的同時,深入探討其作用機制,給臨床上發現體內效果良好的靶向FtsZ蛋白的先導化合物提供堅實的理論基礎。
