β-氨基丁酸對煙草黑脛病的抗性誘導
2023-02-21 06:08:22任錫躍朱發亮王志江葉賢文王建軍
煙草科技 2023年1期
關鍵詞:煙草
任錫躍,劉 濤 ,朱發亮,王志江,葉賢文,梅 堅 ,王建軍
1. 云南農業大學農學與生物技術學院,昆明市盤龍區灃源路452 號 650201
2. 云南農業大學西南中藥材種質創新與利用國家地方聯合工程研究中心,昆明市茨壩鎮青松路21 號 650201
3. 云南農業大學云南省藥用植物生物學重點實驗室,昆明市茨壩鎮青松路21 號 650201
4. 云南省煙草公司昆明市公司,昆明市盤龍區北京路523 號 650051
寄生疫霉(Phytophthora parasitica)屬卵菌綱(Oomycetes)、霜 霉 目(Peronosporales)、腐霉科(Eythictceae)、疫霉屬(Phytophthora),是一種寄主范圍較廣泛的病原菌,可侵染煙草、茄子和番茄等植物[1]。其中,P. parasitica 侵染煙草可引起煙草黑脛病,染病煙株莖基部產生黑斑,葉片自下而上變黃,最后全株枯萎,對我國的煙葉生產造成巨大的損失。目前常使用甲霜靈、烯酰嗎啉等農藥防治煙草黑脛病,但大量使用農藥會出現藥品殘留、病原菌產生抗藥性和環境污染等問題,不利于煙草行業的可持續發展[2-3]。
植物誘抗劑又名激發子,是一類可誘導寄主植物產生防衛反應的特殊化合物的總稱。相比農藥,植物誘抗劑具有安全、高效和抗性穩定等優點,可為煙草黑脛病防治提供新思路。β-氨基丁酸(β-aminobutyric acid,BABA)是從經暴曬番茄根系中分離得到的一種次生代謝非蛋白氨基酸[4],作為一種高效的植物誘抗劑,能夠增強植物對多種病害的抵抗能力。Wang 等[5]研究發現,BABA 處理可誘導PR 表達量顯著增加,有助于葡萄抵御灰霉病菌(Botrytis cinerea)的侵染;Yao 等[6]研究表明,BABA有利于異黃酮生物合成和胼胝質沉積,從而增強大豆對蚜蟲的抗性;何小龍等[7]報道,5.0 mmol/L BABA 可提高煙草對煙草花葉病毒(Tobacco mosaic virus,TMV)的抑制率,降低田間煙草花葉病的發病率和病情指數。然而,目前關于BABA 誘導煙草對P. parasitica 引起的煙草黑脛病抗性的研究還鮮見報道。為此,以云煙87為試驗材料,對供試煙草分別進行BABA 和蒸餾水噴施處理,并用P.parasitica 侵染處理后的煙草,調查BABA 對煙草黑脛病的防治效果,測定并分析BABA 處理前后煙葉的防御酶活性、活性氧和MDA含量以及抗性相關基因的表達量變化,以期為BABA 在煙草病害防治中的應用提供依據。
1 材料與方法
1.1 試驗材料與試劑
供試煙草品種為云煙87。煙苗培育采用漂浮育苗法,煙草長至成苗期時移栽至裝有滅菌營養土的塑料缽中,待煙草長到5~6 葉期時用于試驗處理。供試菌種為P. parasitica,由西南生物多樣性實驗室分離并保存。BABA購于上海麥克林生化科技有限公司。
1.2 試驗設計
1.2.1 BABA對煙草黑脛病的防治效果測定
試驗共設置2個處理(處理A:噴施3次蒸餾水,每次間隔3 d,最后一次噴施3 d 后用菌絲塊創傷莖基部接種法[8]接種 P. parasitica;處理 B:噴施 3 次5 mmol/L BABA,每次間隔3 d,最后一次噴施3 d后接種P. parasitica),每個處理共計煙株10 株。處理后,以株為單位對煙草黑脛病的嚴重度進行分級(0~9級)調查。其中全株無病的煙株記為0級;莖部病斑不超過莖圍的1/2,或1/3以下葉片凋萎的煙株記為1級;莖部病斑環繞莖圍 1/3~1/2,或1/3~1/2 葉片輕度凋萎,又或下部少數葉片出現病斑的煙株記為3級;莖部病斑超過莖圍的1/2,但未全部環繞莖圍,或1/2~2/3 葉片凋萎的煙株記為5 級;莖部病斑全部環繞莖圍,或2/3以上葉片凋萎的煙株記為7級;病株基本枯死的煙株記為 9 級。依據 GB/T 23222—2008[9]統計發病率、病情指數和防治效果,計算公式:

1.2.2 煙葉生理生化指標和抗性相關基因表達量測定
試驗共設置4 個處理(處理1:噴施蒸餾水,不接種P. parasitica;處理2:噴施5 mmol/L BABA,不接種 P. parasitica;處理 3:噴施蒸餾水,3 d 后接種P. parasitica;處理4:噴施5 mmol/L BABA,3 d 后接種P. parasitica),每個處理共計煙株20 株,處理4 d后,取各處理的中部煙葉保存,后續用于煙葉生理生化指標和相關基因表達量的測定。
1.3 試驗方法
1.3.1 防御酶活性測定
過氧化物酶(Peroxidase,POD)、多酚氧化酶(Polyphenol oxidase,PPO)和過氧化氫酶(Catalase,CAT)為防御酶,這類酶不僅能清除植物受病原物侵染后形成的活性氧,而且能催化生成許多與抗性相關的化合物,在植物抗病反應中有重要作用[10]。使用蘇州格銳思生物科技有限公司的酶活性檢測試劑盒檢測上述3種酶活性,試驗方法為分光光度法[11],試驗儀器為紫外-可見分光光度計(UV-2450,日本島津公司),依據試劑盒說明書中的計算方法計算POD、PPO和CAT活性。
1.3.2 超氧陰離子(O2-)和丙二醛(MDA)的含量測定
超氧陰離子自由基的含量可間接反映組織細胞受損狀況和抗性強弱;MDA的含量多少與膜脂過氧化程度、膜結構的受傷程度和植物的自我修復能力密切相關[12]。使用蘇州格銳思生物科技有限公司O2-和MDA 檢測試劑盒檢測 O2-和MDA 的含量(質量分數),試驗方法為分光光度法[11],試驗儀器為紫外-可見分光光度計(UV-2450,日本島津公司),依據試劑盒說明書中的計算方法計算O2-和MDA含量。
1.3.3 抗性相關基因表達量測定
用TRIzol 法提取煙草總RNA,并進行純化。總RNA 用于反轉錄合成第一條鏈cDNA。將cDNA 作為模板,進行qPCR反應。反應體系為:cDNA 1 μL、SYBR Green超混合液10 μL、上游引物和下游引物各1 μL、ddH2O 7 μL。PCR 反應條件:95 ℃預變性3 min;95 ℃變性10 s,60 ℃退火30 s,72 ℃延伸30 s,40個循環。用于qPCR的引物信息見表1。以Actin為內參基因,參照公式2-ΔΔCT計算基因的相對表達量[12]。
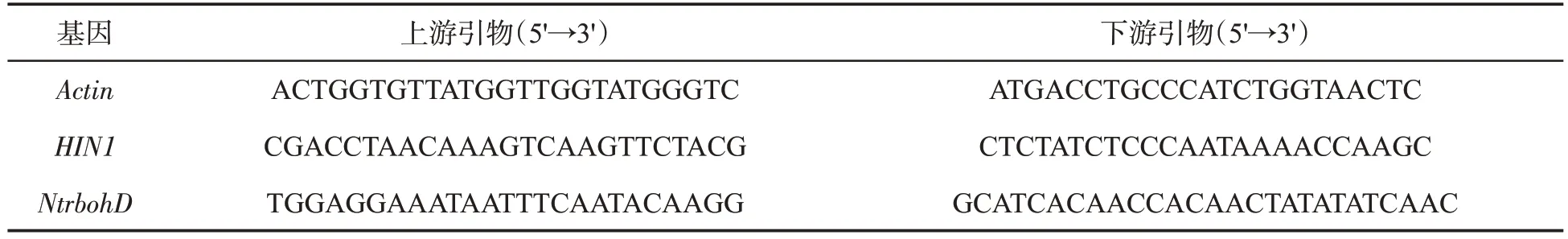
表1 實時定量PCR分析的基因和引物Tab.1 Genes and primers for real time quantitative PCR analysis
1.4 數據處理
使用Microsoft Excel 2010軟件進行數據統計分析,使用最小顯著差數法進行數據間差異的顯著性檢驗,使用GraphPad Prism軟件制圖。
2 結果與分析
2.1 BABA對煙草黑脛病的抗性誘導效果
研究發現兩個處理的煙草黑脛病發病率都為100.0%。處理A 的煙草病情指數為35.6,處理B 的煙草病情指數為15.6。5 mmol/L BABA對煙草黑脛病的防治效果為56.3%(表2)。
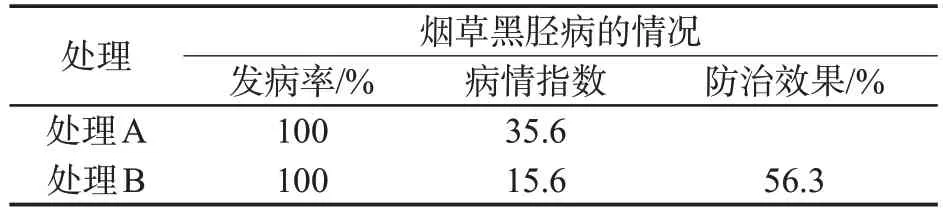
表2 不同處理對煙草黑脛病的防治效果Tab.2 Control effects of different treatments against tobacco black shank disease
2.2 BABA誘導煙葉產生枯斑與HIN1表達量
BABA處理煙草3 d后煙葉漸黃,形成壞死型枯斑,且下部煙葉枯斑的量和面積較上部煙葉大而多(圖1A),該病癥與超敏反應(Hypersensitive response,HR)的癥狀類似。HIN1(Harpin-induced gene 1)為Spm(精氨)的蛋白誘導基因,而Spm 可作為信號參與超敏反應途徑[13],測定該基因的循環閾值(Cycle threshold,CT)并計算其表達量,發現處理2的HIN1 基因表達量顯著高于處理1,處理4 的HIN1基因表達量顯著高于處理3(圖1B)。
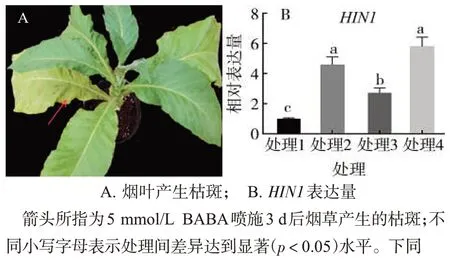
圖1 BABA噴施后煙葉的超敏反應Fig.1 Hypersensitive response of tobacco leaves after spraying BABA
2.3 BABA對煙草葉片防御酶的影響
由圖 2 可見,處理 2 的 POD、PPO 和 CAT 活性分別比處理1顯著提高了129.6%、81.3%和32.4%;處理4的POD和PPO分別比處理3顯著提高了264.4%和169.0%。表明在煙草感染黑脛病的過程中,BABA可誘導煙草葉片中PPO、POD和CAT活性提高。
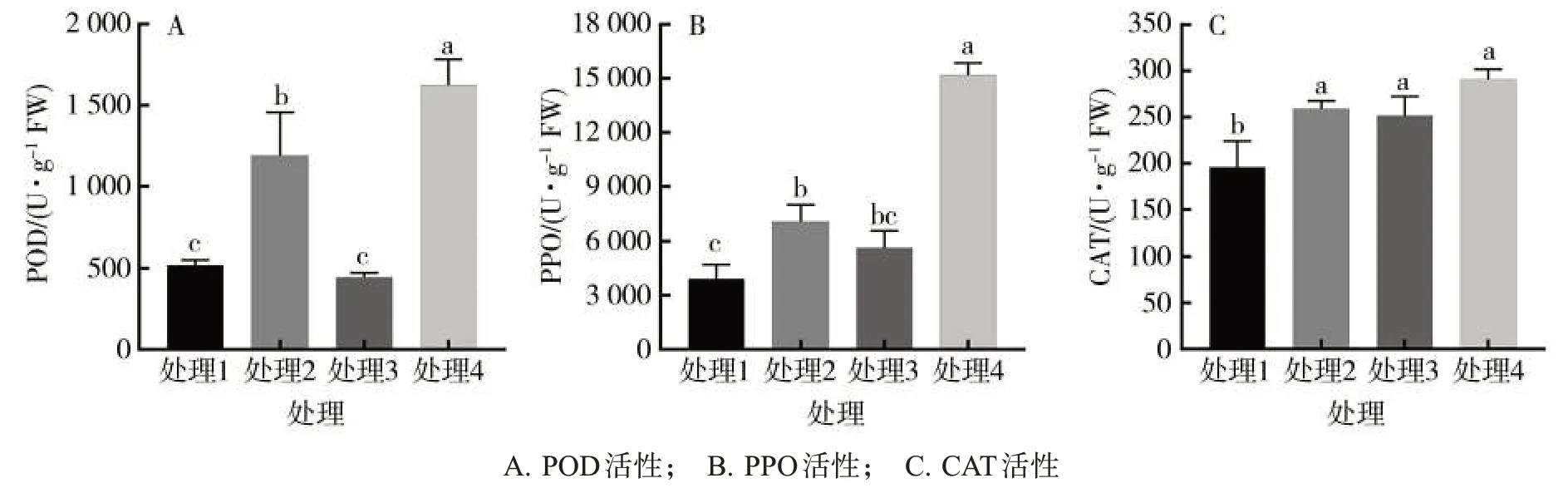
圖2 不同處理對煙草防御酶的影響Fig.2 Effects of different treatments on tobacco defense enzymes
2.4 BABA 對煙草 O2-、MDA 含量以及抗性相關基因表達量的影響
處理2 的O2-含量比處理1 顯著增加142.2%;處理4的O2-含量比處理3顯著增加31.8%,但處理4與處理2相比,O2-含量顯著降低(圖3A)。此外,處理2的MDA 含量比處理1 顯著增加311.0%,處理3 和處理4 的MDA 含量均比處理2 顯著降低,且處理3 和處理4的MDA含量無顯著差異(圖3B)。NtrbohD基因可編碼NADPH氧化酶[又名呼吸爆發氧化酶同源蛋白(Respiratory burst oxidase homolog ,RBOH)],而NADPH氧化酶又可促進活性氧的產生[14],故測定該基因的表達水平,發現處理1 和處理2 的NtrbohD基因相對表達量無顯著差異;處理4 的NtrbohD基因相對表達量比處理3顯著降低56.9%(圖3C)。
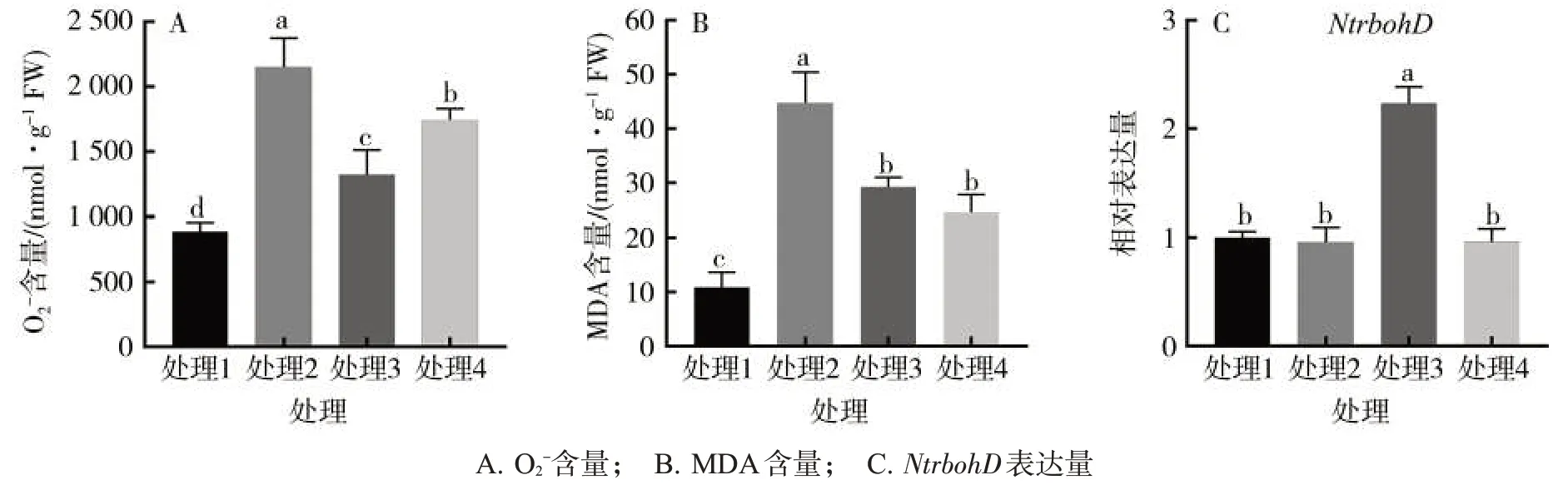
圖3 不同處理對煙草活性氧及相關基因的影響Fig.3 Effects of different treatments on reactive oxygen and related genes in tobacco
3 討論
5 mmol/L BABA 噴施煙草3 d 后煙葉產生較多枯斑,根據Siegrist等[15]的研究,BABA誘導葉面枯斑的產生是一種超敏反應,該反應可阻止P. parasitica的擴散,減弱黑脛病對植株的危害。本研究中發現 BABA 誘導 PPO、POD 和 CAT 活性提高,這與前人[16-20]的研究結果一致。其中,PPO 和 CAT 活性的提高,可維持煙草對氧自由基的清除能力,保持植物體內氧自由基的平衡。此外,PPO 和POD 活性的提高可使煙草中酚類物質和芳香胺氧化為植保素和木質素,抵御P. parasitica侵染[21-22]。
本研究中發現BABA 可誘導煙葉活性氧O2-含量顯著升高,活性氧能直接殺死病原菌并激活抗病基因的表達,其迸發是植物產生的最快的防御反應,在植物免疫過程中起著至關重要的作用,可增強煙草對黑脛病的抗性[23]。本研究中還發現處理1和處理2 NtrbohD 的表達量無顯著差異;處理4 NtrbohD的表達量比處理 3 顯著降低,這與 Peng 等[24]發現化學誘抗劑氟噻唑吡乙酮(Oxathiapiprolin)在增加擬南芥活性氧的同時還上調了RBOHD 表達水平的研究結果不一致,推測原因是BABA 誘導CAT 和POD酶活性提高從而抑制了接種P. parasitica 后活性氧的迸發,使該基因的表達水平達到動態平衡,在激活防御體系的同時緩解了活性氧對植物的氧化損傷。
4 結論
使用5 mmol/L BABA 處理煙草,發現BABA 可誘導煙草產生超敏反應,并提高HIN1 基因的表達量。此外,煙草未接種P. parasitica時,BABA處理可誘導煙葉O2-含量增加142.2%,說明BABA可促使活性氧迸發。煙草接種P. parasitica 后,BABA 處理可誘導煙葉POD 和PPO 活性分別提高264.4%和169.0%,但該處理的NtrbohD 基因表達量降低了56.9%。BABA 處理可維持煙草對由P. parasitica 傷害產生的氧自由基的清除能力,抑制活性氧迸發,緩解活性氧對煙草的傷害,對煙草黑脛病的防治效果達到56.3%。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
