植物ABC轉(zhuǎn)運蛋白功能研究進(jìn)展
2023-02-15 07:26:42唐桃霞孔維萍任凱麗
西北農(nóng)業(yè)學(xué)報 2023年1期
唐桃霞,孔維萍,任凱麗,程 鴻
(甘肅省農(nóng)業(yè)科學(xué)院 蔬菜研究所,蘭州 730070)
ABC轉(zhuǎn)運蛋白,又名ATP結(jié)合盒式蛋白(ATP-binding cassette transporter,ABC),得名于含有一個腺苷三磷酸(ATP)結(jié)合盒,且依賴于 ATP 水解產(chǎn)生的能量而實現(xiàn)底物在細(xì)胞內(nèi)外的跨膜轉(zhuǎn)運,是目前已知數(shù)量和功能最多的一類蛋白[1]。據(jù)報道,ABC轉(zhuǎn)運蛋白家族在所有生物中廣泛存在,已在人類、酵母和大腸桿菌中發(fā)現(xiàn)了數(shù)量不等的ABC轉(zhuǎn)運蛋白及其基因,而在陸生植物中鑒定到的 ABC 轉(zhuǎn)運蛋白數(shù)量遠(yuǎn)高于其他生物[2-3],其中,模式作物擬南芥和水稻基因組中分別存在130個和128個 ABC 轉(zhuǎn)運蛋白[4-5],此外,玉米(Zeamays)[6]、小麥(Triticumaestivum)[7]、甘藍(lán)型油菜(Brassicanapus)[8]、百脈根(Lotuscorniculatus)[9]、菠蘿(Ananascomosus)[10]以及番茄(Solanumlycopersicum)[11]中均有報道,而研究最多的擬南芥僅有一小部分ABC 轉(zhuǎn)運蛋白被鑒定出來[12],還有大部分成員的轉(zhuǎn)運底物及其功能尚不清楚,而有關(guān)其他作物ABC轉(zhuǎn)運蛋白及其功能還未見報道。因此,本文通過對已鑒定的植物ABC轉(zhuǎn)運蛋白成員及功能進(jìn)行歸類總結(jié),旨在為研究其他作物ABC轉(zhuǎn)運蛋白及功能提供思路和依據(jù)。
1 ABC轉(zhuǎn)運蛋白的結(jié)構(gòu)及分類
典型的ABC轉(zhuǎn)運蛋白由親水核苷酸結(jié)合結(jié)構(gòu)域(Nucleotide-binding domain, NBD)和疏水跨膜結(jié)構(gòu)域(Transmembrane domain, TMD)組成[13],其中親水性的 NBDs 包含3個保守的特征基序:Walker A,Walker B 和ABC 標(biāo)簽序列(又稱 Walker C)[4],因此,其結(jié)構(gòu)和序列在 ABC 轉(zhuǎn)運蛋白家族中高度保守,而疏水的TMD是由數(shù)個跨膜 α- 螺旋構(gòu)成,形成跨膜運輸通道[14],轉(zhuǎn)運時NBDs負(fù)責(zé)結(jié)合并水解 ATP 使其構(gòu)象發(fā)生改變,從而激活轉(zhuǎn)運過程,而 TMDs則形成通道使得底物通過脂質(zhì)雙分子層,二者共同參與實現(xiàn)物質(zhì)轉(zhuǎn)運[15]。ABC 轉(zhuǎn)運蛋白家族的轉(zhuǎn)運分為向外和向內(nèi)兩種轉(zhuǎn)運方式,大多數(shù)的ABC向內(nèi)轉(zhuǎn)運蛋白還包括一個可溶性的位于細(xì)胞外或是錨定在膜上的胞外區(qū)結(jié)合蛋白(Substrate-binding protein,SBP),SBP 可以結(jié)合不同種類的底物并把底物傳遞到相應(yīng)的 TMDs 中。圖1展示了ABC 轉(zhuǎn)運蛋白轉(zhuǎn)運過程中依靠構(gòu)象改變的轉(zhuǎn)運機(jī)制[16]。
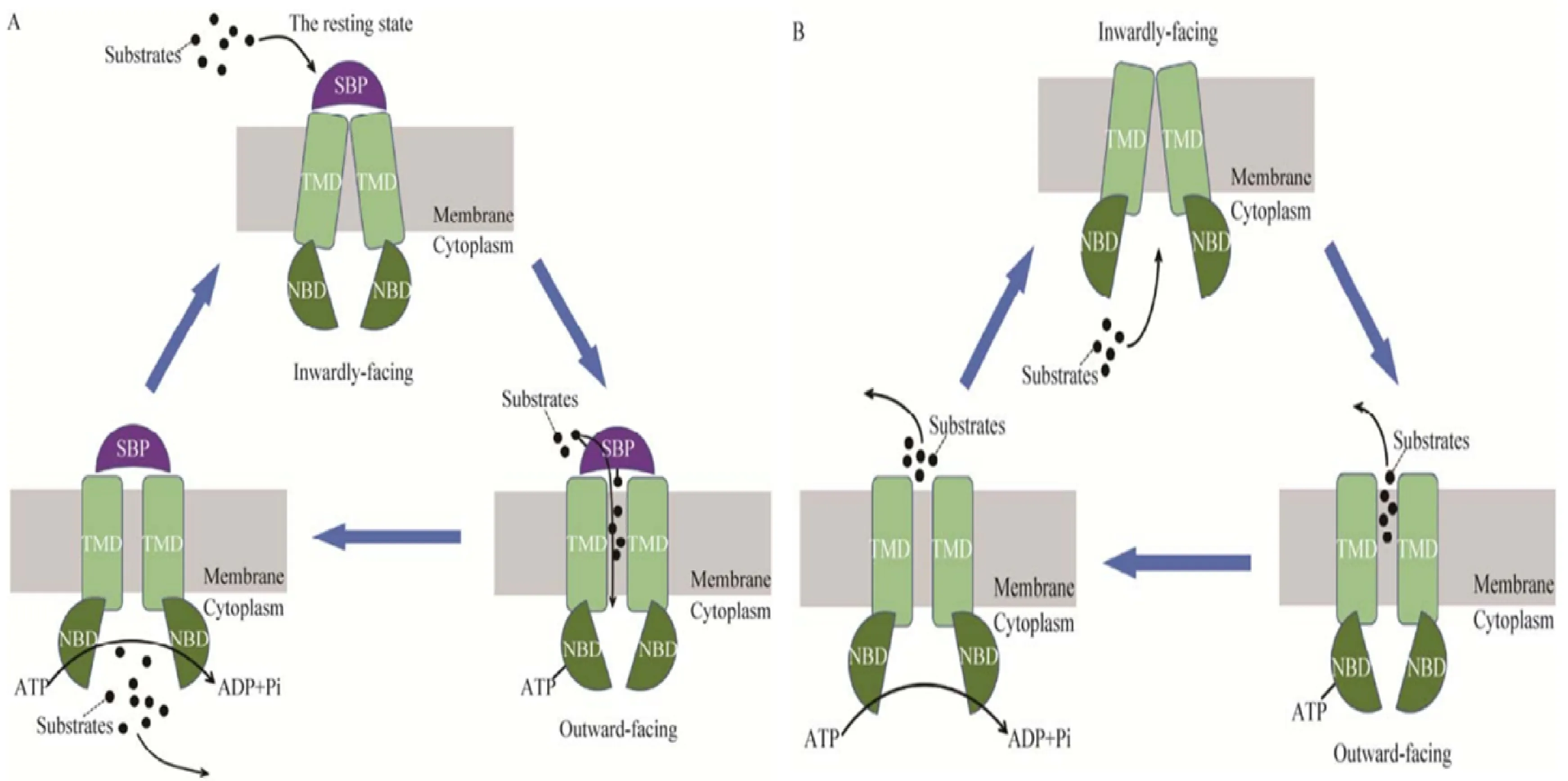
A.向內(nèi)轉(zhuǎn)運機(jī)制;B.向外轉(zhuǎn)運機(jī)制A.Import mechanism; B.Export mechanism
由于 ABC 蛋白包含的NBD 和 TMD 數(shù)量以及排列方式存在差異,其又被分為全分子 ABC 蛋白、半分子ABC 蛋白和可溶性ABC蛋白,全分子 ABC 蛋白由 2 個 NBD 和 2 個 TMD構(gòu)成,4 個結(jié)構(gòu)域結(jié)合在一起才可執(zhí)行轉(zhuǎn)運功能,而半分子 ABC 蛋白僅由 1 個 NBD 和 1個 TMD 構(gòu)成,從而排列形成 TMD-NBD 或NBD-TMD,任一的 TMD-NBD 或 NBD-TMD 是沒有功能的,它們必須形成同源或者異源二聚體才能發(fā)揮功能[17]。通常,研究者們按照國際命名系統(tǒng)將植物ABC分為8個亞族(ABCA~ABCG和ABCI),植物中未發(fā)現(xiàn)ABCH 亞族成員,但存在一類特殊的細(xì)菌類型 ABC 轉(zhuǎn)運蛋白,被命名為 ABCI 亞族[1]。目前,研究較多的為 ABCG、ABCB和ABCC[18]。ABC 轉(zhuǎn)運蛋白參與植物體中多種重要的轉(zhuǎn)運過程,其轉(zhuǎn)運功能需要NBD和TMD的共同參與。由于NBD的高度保守性和每個亞族結(jié)構(gòu)的相似性,導(dǎo)致其轉(zhuǎn)運功能具有專一性,也具有相似性。
2 ABC轉(zhuǎn)運蛋白功能
2.1 ABCA亞族
迄今為止,在人類基因組中已鑒定出12個ABCA亞族成員,主要與身體各部位的脂質(zhì)轉(zhuǎn)運有關(guān)[19],而擬南芥中的 ABCA1 與人類 ABCA1具有高度的序列同源性,推測植物ABCA1 也可能具有脂質(zhì)運輸功能[20]。目前,在擬南芥中也發(fā)現(xiàn)12個ABCA成員,且只有 At ABCA9的功能被鑒定出來,并發(fā)現(xiàn)其過量表達(dá)可促進(jìn)種子三酰甘油 (Triacylglycerol,TAG)的生物合成,表明AtABCA9 可能是增加油籽脂質(zhì)產(chǎn)量的有力工具[12,21],隨后李健春[22]在大豆中發(fā)現(xiàn)GmABCA7轉(zhuǎn)運蛋白,并將其定位于過氧化物酶體,表明其在擬南芥中的過量表達(dá)影響脂肪酸的 β氧化并促進(jìn)種子萌發(fā);此外,擬南芥根的轉(zhuǎn)錄組分析表明 AtATH14 和 AtATH15 表達(dá)對鹽脅迫有反應(yīng)[23],也有報道AtABCA1 與花粉萌發(fā)、種子萌發(fā)和種子成熟有關(guān)[9,24],但具體功能未做解析。
ABCA包括全分子的AOH和半分子的ATH兩種類型。在蔬菜作物番茄中發(fā)現(xiàn),SlABCA1 是唯一全分子ABCA 和最大的 ABC 蛋白,由1 910 個氨基酸殘基組成,另一主要特征是 ABCA 亞族在雙子葉植物中存在一個全分子的ABCA (AOH),包括番茄[11]、擬南芥[4]、百脈根[9]和葡萄[24],截至目前還未在水稻[25]、玉米[6]等單子葉植物中發(fā)現(xiàn),這表明這種全分子 ABCA可能特定于雙子葉植物;據(jù)番茄[11]的基因表達(dá)譜顯示 SlABCA1 和 SlABCA2 優(yōu)先在根中表達(dá),它們可能參與根的分泌活動,SlABCA4-7 在花中特異性表達(dá),表明在花器官中具有特定功能。此外,吳鵬等[26]在黃瓜中克隆到控制ABCA19轉(zhuǎn)運蛋白的基因,通過功能分析發(fā)現(xiàn)黃瓜果實中的AB-CA19對霜霉威脅迫的響應(yīng)比較迅速。以上研究只是對ABCA 成員做了初步表達(dá),其轉(zhuǎn)運底物可能比較多樣,具體功能還需進(jìn)一步解析驗證。
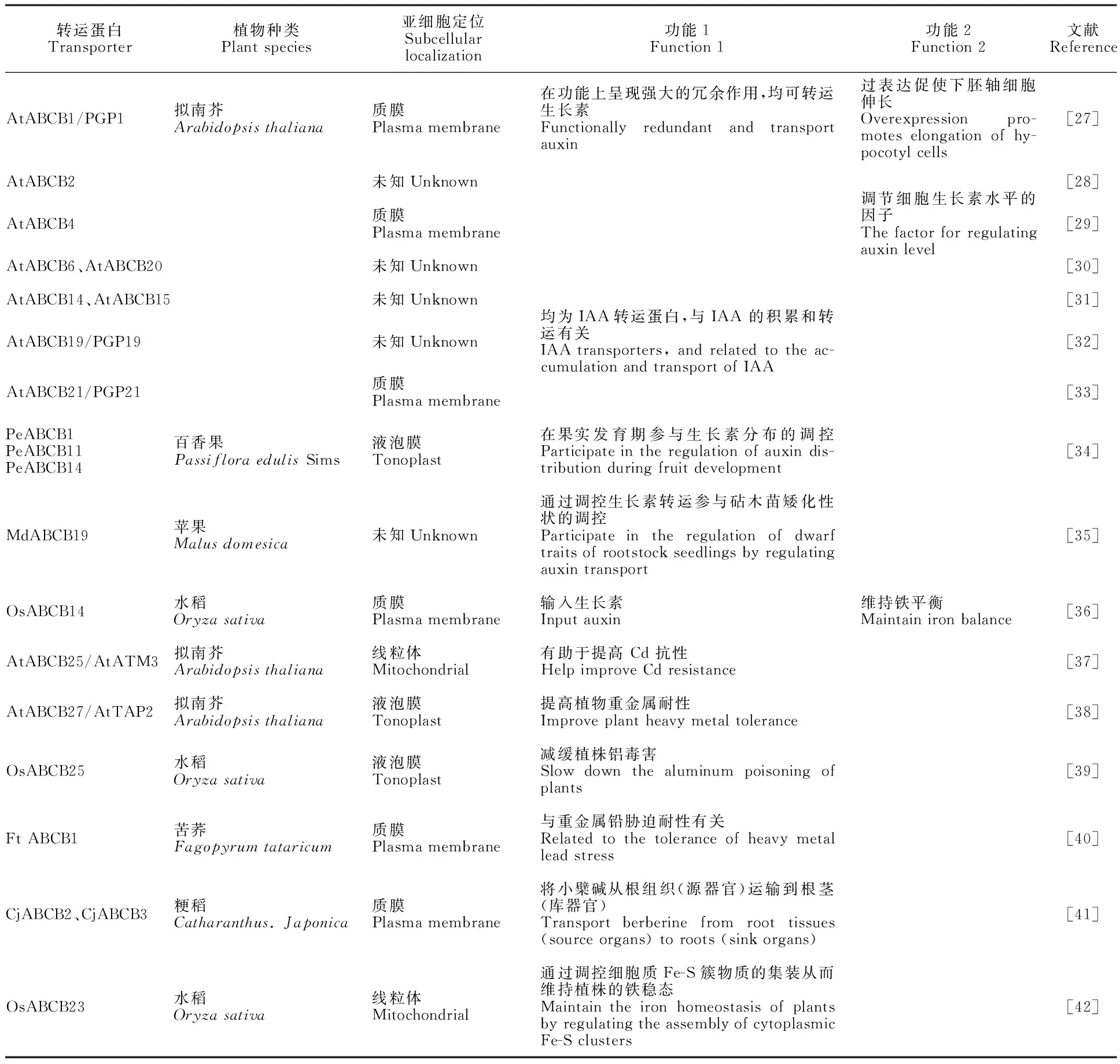
表1 植物ABCB轉(zhuǎn)運蛋白功能Table 1 Function of plant ABCB transporters
2.2 ABCB亞族
ABCB是僅次于ABCG的第二大亞族,擬南芥中發(fā)現(xiàn)29個ABCB成員[4]。據(jù)表1可知,已鑒定的大多數(shù)ABCB成員主要與激素類物質(zhì)(生長素)的運輸及調(diào)控有關(guān),且部分成員功能冗余,均能轉(zhuǎn)運生長素;其次,ABCB還參與離子及重金屬轉(zhuǎn)運,如在番茄中研究發(fā)現(xiàn)大多數(shù) SlABCBs 在根中高度表達(dá),這可能表明這些 SlABCB 參與根中的離子和重金屬運輸[11]。除上述功能外,ABCB還具有轉(zhuǎn)運生物堿和維持鐵穩(wěn)態(tài)的作用。綜上可知,還需加大對ABCBs功能的研究,以期為闡釋植物對激素和非生物脅迫響應(yīng)的生理機(jī)制奠定基礎(chǔ)。
2.3 ABCC亞族
根據(jù)國內(nèi)外研究報道發(fā)現(xiàn),C亞族目前已鑒定的功能之一是轉(zhuǎn)運代謝產(chǎn)物(花青素、植酸、葉酸等物質(zhì)),有研究表明,玉米ZmMRP3和葡萄VvABCC1均具有運輸液泡中花青素的功能[43-44];擬南芥的AtABCC2可能與其他的ABCC轉(zhuǎn)運蛋白共同介導(dǎo)營養(yǎng)組織中花青素和其他類黃酮的液泡運輸[45];還有研究表明,部分ABCC轉(zhuǎn)運蛋白具有多種功能,如擬南芥AtABCC5/AtMRP5[46]和水稻 OsABCC13[47]參與種子植酸的轉(zhuǎn)運和含磷產(chǎn)物的積累,其中AtABCC5/AtMRP5還是保衛(wèi)細(xì)胞脫落酸和 Ca2+信號轉(zhuǎn)導(dǎo)過程中保衛(wèi)細(xì)胞離子通道的中央調(diào)節(jié)器[48];而擬南芥AtABCC4/AtMRP4不僅參與葉酸轉(zhuǎn)運[49],還介導(dǎo)氣孔孔徑的復(fù)雜調(diào)節(jié)[50]。
二是具有解毒功能,擬南芥AtABCC1、AtABCC2和AtABCC3均定位于液泡膜,其中AtABCC1和AtABCC2被證明賦予對砷、鎘和汞的耐受性[51-53],是因為植物螯合素 (PC) 與Cd需要以 PC-Cd 復(fù)合物的形式運輸?shù)揭号葜衅鸬浇舛咀饔茫鳤tABCC3是一種PC-Cd 復(fù)合物的轉(zhuǎn)運蛋白,其活性受Cd 調(diào)節(jié)并與AtABCC1/AtABCC2 的活性協(xié)調(diào);此外,AtABCC6 在重金屬Cd脅迫下表達(dá)上調(diào),也參與重金屬解毒作用[54];苦蕎FtABCC9在重金屬Pb脅迫下表達(dá)上調(diào),說明此蛋白也響應(yīng)重金屬脅迫[40]。這些轉(zhuǎn)運蛋白為植物基因工程提供了有用的工具,可使植物增強對金屬耐受性和積累,這是植物修復(fù)的理想特征[52]。
2.4 ABCG亞族
在ABC轉(zhuǎn)運蛋白家族中,ABCG是其中最大的一個亞族,其功能廣泛,有全分子的PDR和半分子的WBC兩種類型。已知的擬南芥、亞麻、水稻、玉米、鐵皮石斛、毛果楊、二穗短柄草、葡萄、番茄、菠蘿及蜜柚中分別有43、85、50、54、37、74、44、71、70、42和22個ABCG轉(zhuǎn)運蛋白(表2),分別占ABC蛋白的33.08%、42.93%、39.06%、41.54%、42.05%、38.54%、33.08%、39.23%、45.45%、42%和44%,平均數(shù)量占ABC家族的40%以上,說明ABCG是ABC家族最為重要的成員。從已報道的研究來看,已鑒定ABCG成員的功能多樣,具體類別見表3。

表2 部分植物中ABC轉(zhuǎn)運蛋白數(shù)量Table 2 Number of ABC transporters in some plants
由表3可知,ABCG是植物體生命活動的重要參與者。首先,可轉(zhuǎn)運植物體內(nèi)的代謝物,包括脂類、烯類,生物堿類等,還調(diào)節(jié)花散發(fā)揮發(fā)性化合物的功能,除轉(zhuǎn)運內(nèi)源代謝物外,還可轉(zhuǎn)運外源物質(zhì)百草枯等除草劑。其次,介導(dǎo)激素類物質(zhì)的運輸(ABA),如AtABCG25、AtABCG31、AtABCG30、AtABCG40、AtPDR12/AtABCG40均具有轉(zhuǎn)運脫落酸(ABA)的作用。此外,ABCG參與重金屬脅迫/逆境脅迫響應(yīng),可提高植物對非生物脅迫的抗逆性。
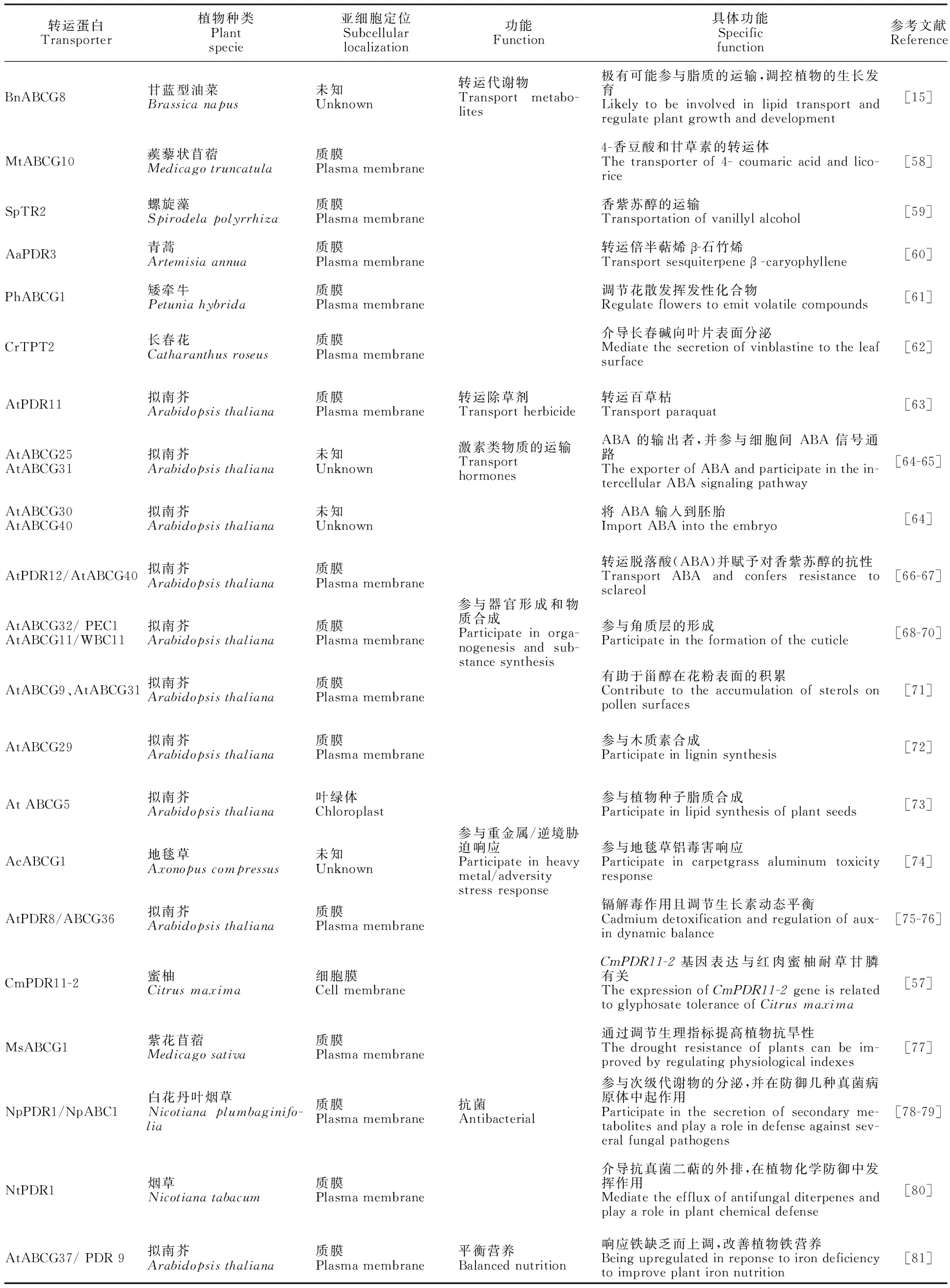
表3 植物ABCG轉(zhuǎn)運蛋白功能Table 3 Functions of plant ABCG transporters
植物角質(zhì)層不僅減少水分流失,還具有化學(xué)防御功能,研究發(fā)現(xiàn)AtABCG32/PEC1和AtABCG11/WBC11均參與角質(zhì)層的形成[68-70],ABCG9 和 ABCG31 通過促進(jìn)甾醇在花粉表面的積累而支持花粉外殼的成熟,從而增強花粉的適應(yīng)性[71];木質(zhì)素作為植物細(xì)胞壁的重要組成部分,其轉(zhuǎn)運機(jī)制還未可知,Alejandro等[72]研究發(fā)現(xiàn)AtABCG29參與擬南芥中木質(zhì)素合成,填補了人們對木質(zhì)素生物合成理解的關(guān)鍵空白,這可能為木質(zhì)素工程開辟新的方向。
此外,ABCG還具有抗菌方面的作用,如白花丹葉煙草(Nicotianaplumbaginifolia)中NpPDR1/NpABC1 通過參與次級代謝物的分泌,在防御真菌病原體中起作用[78-79];煙草NtPDR1通過介導(dǎo)抗真菌二萜的外排,在植物的化學(xué)防御中發(fā)揮作用[80],這為植物抗菌方面的進(jìn)一步研究奠定了基礎(chǔ)。
2.5 其余亞族
除亞族 B、C、G研究較多外,其他亞家族的報道相對較少,它們是否具有與其他亞族相似的功能未有明確的報道。根據(jù)模式作物擬南芥的研究,推測蔬菜作物番茄ABCD可能參與長鏈脂肪酸的過氧化物酶體輸入,ABCE可能在核糖體生物發(fā)生、翻譯控制和基因沉默調(diào)節(jié)中發(fā)揮作用,ABCI在果實發(fā)育期表達(dá),表明其可能在果實發(fā)育中有特定功能[11],擬南芥ABCF 亞家族成員 AtABCF3 參與根系生長發(fā)育[82];水稻OsABCI7可能涉及到多種葉綠體發(fā)育調(diào)控網(wǎng)絡(luò)[83],具體功能還有待驗證。
3 總結(jié)與展望
綜上發(fā)現(xiàn),植物ABC蛋白家族體系龐大,成員眾多,這些 ABC 蛋白存在于各類細(xì)胞器中,大多定位于質(zhì)膜和液泡膜、還有些定位于線粒體、葉綠體、內(nèi)質(zhì)網(wǎng)等細(xì)胞器上,其轉(zhuǎn)運底物形式多樣,有激素、離子、重金屬、代謝產(chǎn)物等,通過介導(dǎo)底物的轉(zhuǎn)運及平衡,隨時調(diào)控植物的生理狀態(tài)。就目前研究可知,ABCB參與生長素及重金屬轉(zhuǎn)運,ABCC參與轉(zhuǎn)運代謝產(chǎn)物(花青素、植酸、葉酸等物質(zhì))、還具有重金屬解毒功能,ABCG功能較多,參與代謝產(chǎn)物和激素轉(zhuǎn)運、植物器官形成和細(xì)胞發(fā)育、逆境脅迫、化學(xué)防御等過程,ABCA報道較少,初步表明其與脂質(zhì)代謝有關(guān),其余亞族功能還有待進(jìn)一步研究。總而言之,ABC是一類值得深入研究的轉(zhuǎn)運蛋白,研究其可解析生理機(jī)制,研究其可輔助植物育種。通過總結(jié)上述進(jìn)展的同時,也發(fā)現(xiàn)以下不足。
研究作物種類單一:目前研究主要集中在模式作物和個別作物中,如蔬菜僅有番茄ABC 轉(zhuǎn)運蛋白進(jìn)行了全基因組分析[11]。
研究深度不夠:大多研究只對某些成員做了初步的鑒定和表達(dá),其轉(zhuǎn)運底物尚不明確,具體功能還有待驗證[11,23]。
與應(yīng)用研究結(jié)合不緊密:如品質(zhì)育種已成為研究熱點,代謝產(chǎn)物是影響果實品質(zhì)的關(guān)鍵因素[84],而ABC家族中有許多成員參與轉(zhuǎn)運代謝產(chǎn)物,但未見到關(guān)于此類問題的研究報道。
基于上述研究現(xiàn)狀,筆者認(rèn)為未來研究,需從以下方向著手:(1)開展蛋白互作,挖掘未知功能。對目前功能未知的其他亞族建議開展蛋白互作研究,可有助于加快ABC轉(zhuǎn)運蛋白的研究進(jìn)程;(2)橫向研究,由此及彼。利用已有模式作物的研究基礎(chǔ),加快其他作物ABC蛋白的研究,如擬南芥AtABCA9過量表達(dá)可促進(jìn)種子三酰甘油 (TAG)的生物合成,從而增加油籽脂質(zhì)產(chǎn)量,這是一種源庫調(diào)控效應(yīng),若應(yīng)用于其他油料作物,可能有助于提高其品質(zhì);(3)深度研究,明確機(jī)制,加以應(yīng)用。針對目標(biāo)蛋白加深研究,掌握其生理功能,并延伸至實踐應(yīng)用,如NpPDR1/NpABC1在抗菌方面起作用, AtATH14 和 AtATH15響應(yīng)鹽脅迫,MsABCG1響應(yīng)干旱,若加以深度研究,則可應(yīng)用于植物抗逆育種;(4)多學(xué)科聯(lián)合,準(zhǔn)確解析具體功能。由于其結(jié)構(gòu)的相似性,有許多蛋白功能冗余[27-31],大多研究只是通過模式作物進(jìn)行推測,應(yīng)通過代謝組學(xué)、轉(zhuǎn)錄組學(xué)、生物信息學(xué)及結(jié)構(gòu)生物學(xué)等多學(xué)科聯(lián)合,準(zhǔn)確具體的鑒定其功能。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
體育科技文獻(xiàn)通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計與研究(2019年4期)2019-05-21 07:21:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
