寧夏賀蘭山斑子麻黃種群的性比及雌雄空間格局
2023-01-13 07:58:54劉萬弟高佳慧李小偉
生態學報 2022年24期
關鍵詞:關聯
趙 玲,楊 博,劉萬弟,馬 龍,高佳慧,李小偉
寧夏大學農學院,銀川 750021
性比和空間分布格局對雌雄異株植物種群的生存潛力、遺傳多樣性維持、個體繁殖以及對環境響應等均具有重要意義[1—2]。性比是指雌雄異株種群中所有個體或某一齡級上的雌雄個體數目的比例,反映了種群產生后代的潛力,在一定程度上影響著種群發展動態。一般來說,雌雄異株植物自然種群的性比為1∶1,但大量研究結果表明,雌雄異株植物種群普遍存在性比偏倚的現象[3—5]。導致種群性比偏離的原因有植物自身的因素和環境因素,比如:雌雄植株受精卵構成比例、花粉粒的存活率、花粉管的存活率[6]、性別表達的遺傳決定性[7]、種子發育、萌發率[8]、死亡率、生理生化特性[9]以及雌雄植株壽命差異等均會導致種群性比偏離。此外,環境因子也是造成性比偏離的重要原因[10]。雄株通常比雌株更有抗逆性,環境壓力的增大會導致種群中雄株比例明顯增加,如土壤養分[3]、海拔高度變化[4]等環境變化都會顯著的影響雄樹比例[11]。雌雄株對環境需求、性比格局、相鄰植株敏感程度的差異[3,12]以及雌樹的分布對種子散布的決定作用,都會影響空間分布格局,進而可能會影響種群動態。研究雌雄異株植物的性比結構以及與性別相關的空間格局有助于深化對種群動態的認識[13—14]。
斑子麻黃(Ephedrarhytidosperma)為中國特有種,國家二級保護植物,主要分布于寧夏賀蘭山,生于山前洪積扇或礫石質山坡,具有耐貧瘠、抗旱、抗寒等優良特性,在維持荒漠生態系統多樣性和生態平衡的方面具有不可代替的作用。近年來,隨著賀蘭山東麓的不斷開發和利用,棲息生境遭到破壞,種群數量銳減,瀕臨滅絕。有研究表明:未來氣候變化的過程中斑子麻黃將從適宜的分布區消失[15—16]。由此可見,對斑子麻黃的保護已迫在眉睫,急需開展試驗研究,以便提出有效的保護和管理措施。然而目前國內外相關研究匱乏,本文通過對寧夏賀蘭山4個不同海拔斑子麻黃種群調查與數據分析,旨在回答以下問題:(1)沿不同海拔分布的斑子麻黃種群的性別比例是否偏離1∶1?(2)沿海拔梯度的斑子麻黃種群的雌雄比是否有差異?(3)沿海拔梯度的斑子麻黃種群雌雄植株的空間關聯性是否相關?本研究結果可為國家二級保護植物斑子麻黃的保育提供基礎數據。
1 材料與方法
1.1 研究區域概況
賀蘭山位于阿拉善荒漠東南緣,地貌形態呈東仰西傾。本區域屬于干旱大陸性氣候,全年干旱少雨,寒暑變化強烈,日照強,無霜期短。年均溫8.5℃,年均降雨量200—400 mm,年平均蒸發量2000 mm以上,年日照時數3040 h。海拔1900m以下的洪積扇地區為山體灰鈣土,生長著耐旱的短花針茅(Stipabreviflora)、酸棗(Ziziphusjujubevar.spinosa)、蒙古扁桃(Amygdalusmongolica)、紅砂(Reaumuriasoongarica)、貓頭刺(Oxytropisaciphylla)、斑子麻黃(Ephedrarhytidosperma)、旱榆(Ulmusglaucescens)等疏林和灌叢植被。
1.2 調查方法
1.2.1樣地調查
2021年5月,在賀蘭山東麓選取了4個具有代表性斑子麻黃群落作為樣地(表1)。每個樣地設置了1個30 m × 30 m的樣方,記錄每株斑子麻黃的二維坐標、海拔、性別、株高、冠幅等信息,總計2012株,能夠確定性別有1770株,占87.97%[17—18]。不能確定性別的242株(S1:4株,S2:1株,S3:23株,S4:214株),且性別未定242株中植株d≤20 cm高達218株(d為植株體積的立方根,單位cm),占90%。(S1:3株,S2:1株,S3:22株,S4:192株)20
生長在干旱環境中的灌木,其冠幅和株高之間的關系與灌木的發育階段呈顯著關系[19—20]。本研究參照宋垚彬等[18, 21—23]采用植株體積(株高×冠幅)的立方根(d表示,單位cm)并結合野外斑子麻黃植株形體大小分布頻次及植株是否具有開花能力劃分為3個等級:齡級Ⅰ(幼年),d≤20 cm;齡級Ⅱ(成年),20
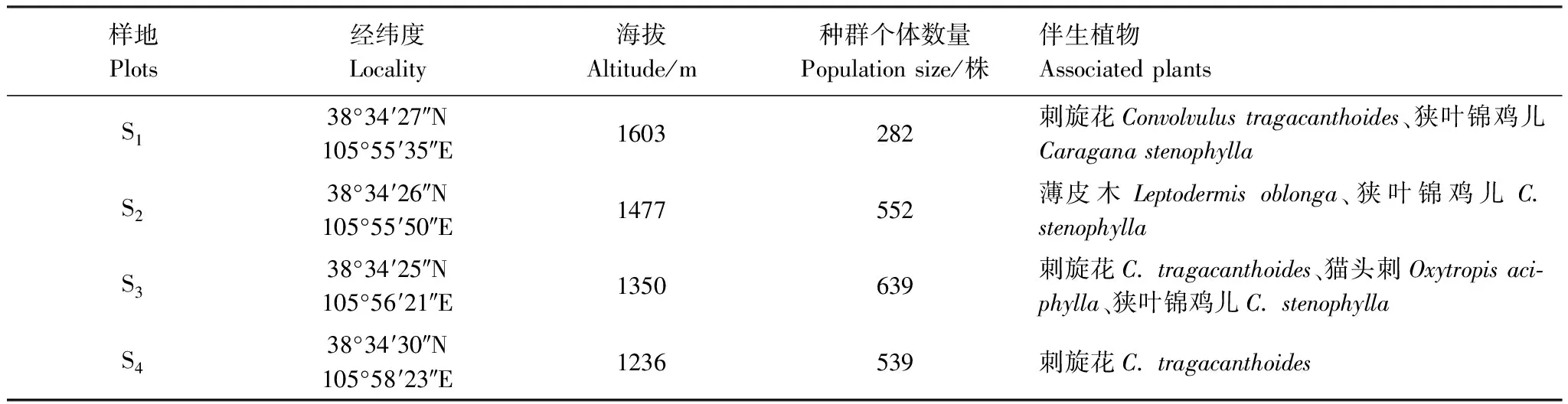
表1 各樣地種群概況和伴生植物
1.2.2土壤采集與測定
分別在S1、S2、S3、S4樣地內,利用土鉆在樣方的中心和四角各取0—40 cm土層樣品,將土壤充分混合,重復3次,自然風干后進行養分的測定,有機質采用重鉻酸鉀加熱法,全氮用凱氏定氮儀(全自動凱氏定氮儀K- 360,BUCHI Labortechnik AG,Switzerland),全磷用NaOH熔融-鉬銻抗比色法,土壤含水量用烘干法,pH采用酸度計測定法(水土比為2.5∶1),土壤容重采用環刀法。
1.3 數據分析
1.3.1形態特征及性比分析
采用SPSS 21.0軟件對數據進行處理;χ2檢驗性比(雄/雌)偏倚1∶1零假設的顯著程度,獨立樣本T檢驗用于比較各樣方中雌雄群體株高、蓋度的差異。Origin 2020、Excel 2010進行作圖。
1.3.2空間格局、空間關聯分析
本研究分別采用點格局分析中Ripley′sK函數與成對相關函數g(r)進行種群整體空間分布與其種內關聯性分析[24—26]。分析4個不同海拔種群所有斑子麻黃雌雄個體雙變量點格局,采用隨機標簽零假設(Randomlabeling model),通過計算以對象1的每一個體為圓心、距離r為半徑的指定環寬的圓環區域內對象2的個體數量,以此來判定雌雄個體在不同尺度下的空間關聯性。沒有開花的植物,無法判定植株的性別,在數據統計分析時未包括性別未定植株。
數據分析采用空間點格局分析軟件Programita 2014完成。空間尺度大小為各樣地最短邊長的一半,Monte-Carlo檢驗模擬循環199次,建立99%的置信區間。對于雌雄雙變量統計分析,某一尺度上,當g(r)高于置信區間上限,表示雌雄在該尺度上呈正相關關系,反之低于置信區間下限則為負相關關系。若在置信區間之間,則表示雌雄間的空間關聯性不顯著,趨于相互獨立[27]。
2 結果與分析
2.1斑子麻黃群體分布
從圖1可以看出,4個不同海拔種群斑子麻黃性別分布圖(30 m ×30 m),S1種群植株最少282株(131∶147∶4,雌株∶雄株∶性別未定,下同);S2種群552株(271∶280∶1);S3種群植株最多639株(269∶347∶23);S4種群539株(134∶191∶214)。空間分布如圖1。
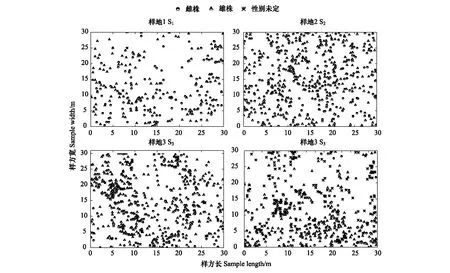
圖1 4個斑子麻黃種群空間分布圖Fig.1 Spatial distribution map of E. rhytidosperma in four populations
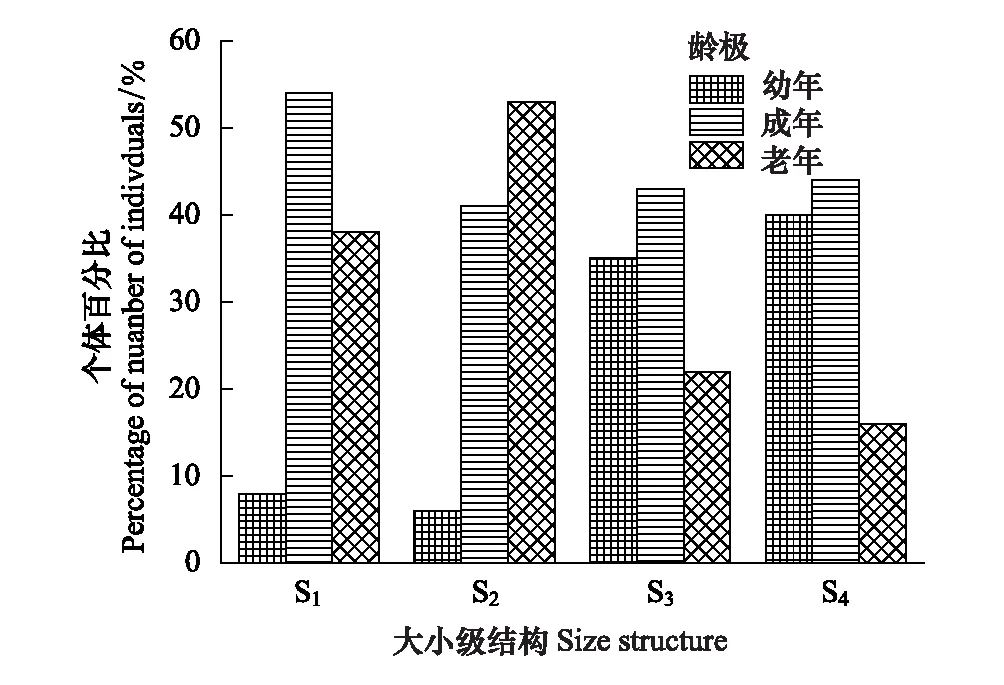
圖2 斑子麻黃種群大小級結構 Fig.2 The size structure of four populations of E. rhytidosperma 齡級Ⅰ(d≤20 cm)代表幼年,齡級Ⅱ,(20 cm
2.2 斑子麻黃種群的大小級結構
從(表1)中可以看出,S2、S3、S4的斑子麻黃種群個體數量較多;S1種群個體數量較少。4個種群中(圖2),S1、S3、S4種群的個體數量均隨d值的增大呈先增后減的“單峰型”, S3、S4種群個體數多度主要集中在幼樹和成年樹兩個等級,兩者總和分別占各自種群個體總數的78%、84%,表明幼樹和成年樹居多。說明這2個種群自然更新良好,在一定時期內表現出增長趨勢。S1、S2種群個體主要分布在成樹和老年兩個等級,其總和分別各自種群個體總數的92%和94%,說明種群缺乏幼體,更新較差,表現出一定的衰退趨勢。
2.3 斑子麻黃種群的性比格局及形態特征
4個不同海拔斑子麻黃種群雌雄植株的數量分布如表2所示。4個不同海拔種群中雄株總計965株,雌株805株,整體上偏雄性,且性比極顯著偏倚1∶1(P<0.01)。其中,S4種群(雄/雌= 1.43)是4個種群中偏雄性最高的種群,S2種群(雄/雌= 1.03)是4個種群中偏雄性最低的種群。S3(雄/雌= 1.29)、S4(雄/雌=1.43)兩個種群有顯著的性別偏倚現象(P<0.05)。
4個不同海拔斑子麻黃雌雄植株共1770株,雌雄蓋度差異極顯著(P<0.01)、株高無顯著差異(P>0.05)。S2種群雌雄株間株高、蓋度均差異極顯著(P<0.01);其次是S1種群雌雄株高(P<0.05)差異顯著、蓋度(P<0.01)差異極顯著,從均值來S1、S2看兩個種群中雄株株高、蓋度均大于雌株;S1、S2種群中雌雄植株的株高和蓋度遠遠大于S3、S4種群的植株。
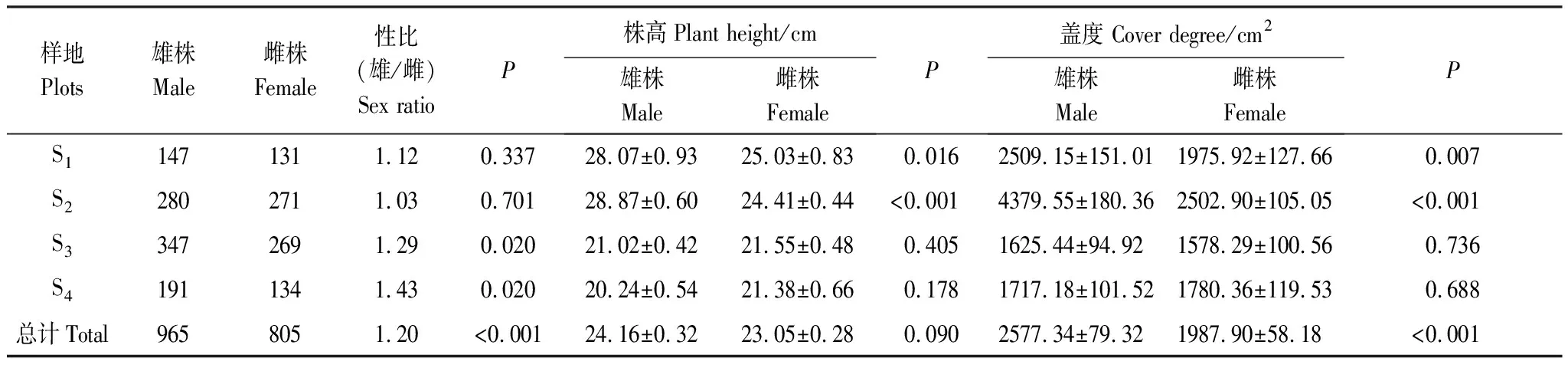
表2 4個種群中雌雄比及雌雄植株的形態特征
2.4 斑子麻黃種群不同徑級的性比格局
為了解斑子麻黃種群內具體的性比格局,對不同徑級上的雌雄植株數量進行統計并檢驗性比(雄/雌)偏離1∶1零假設的顯著程度(χ2檢驗)。對S1、S2、S3、S4種群進行統計檢驗分析,結果如(圖3)所示:S1種群在老年階段偏雄性,性比顯著偏倚1∶1;在幼年階段偏雄性,成年階段偏雌性,兩齡級性比均不顯著偏倚1∶1。S2種群在幼年、成年階段偏雌性;老年階段偏雄性;三齡級性比均顯著偏倚1∶1。S3、S4種群在不同徑級上的性比格局相同,三齡級都偏雄性,且在幼齡性比顯著偏倚1∶1。
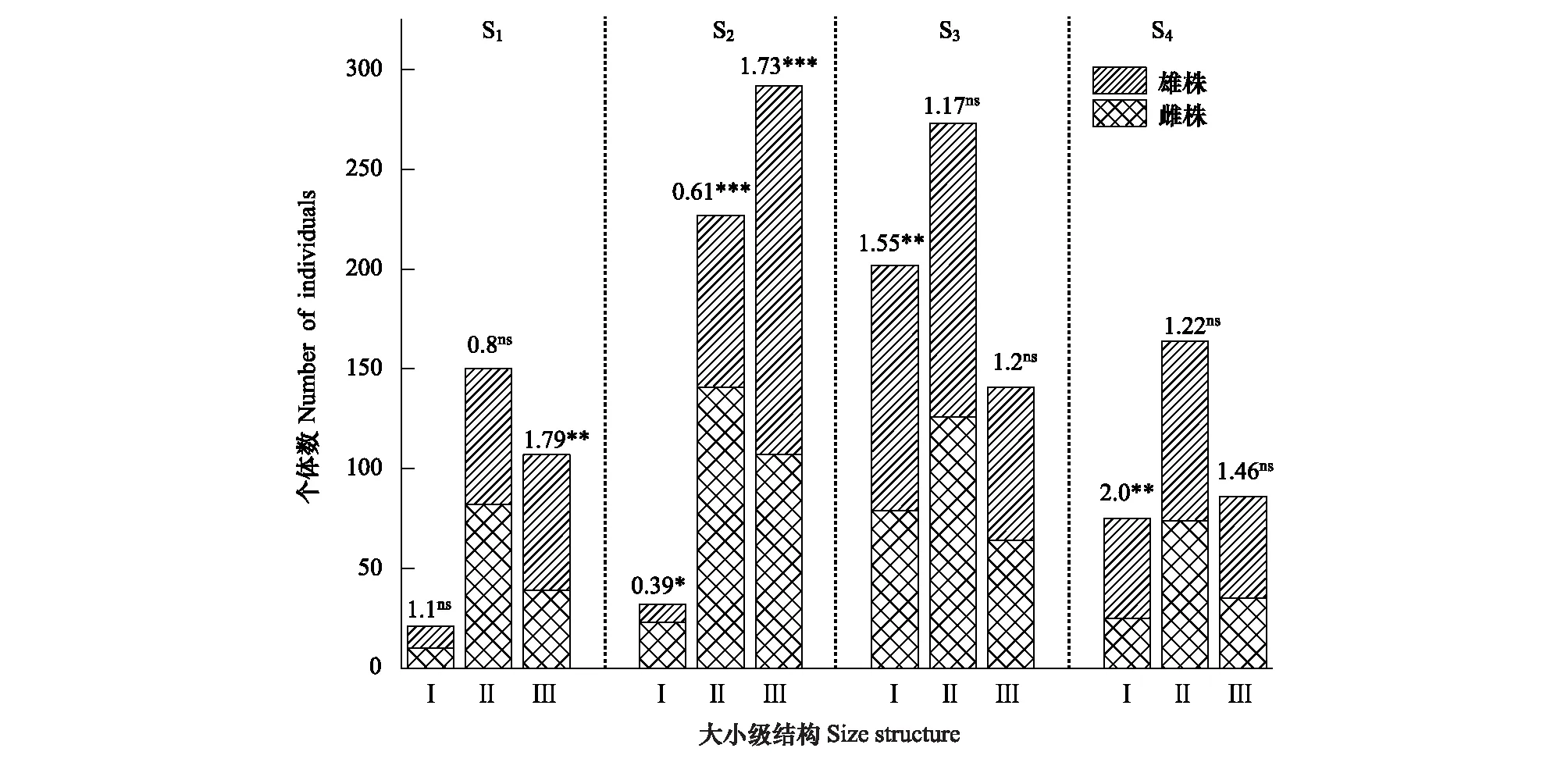
圖3 斑子麻黃種群不同徑級的性比格局Fig.3 Sex ratios for size structure of basal diameter in E. rhytidosperma populations*和**表示性比(雄/雌)性比顯著偏離1∶1(P<0.05和P<0.01),***表示性比(雄/雌)顯著偏離1∶1(P<0.001),ns表示性比(雄/雌)不顯著偏離1∶1(P≥0.05)(χ2檢驗)
2.5 斑子麻黃雌雄植株的空間關聯性
雙變量空間關系分析表明(圖4),4個種群雌雄植株間的相關性不完全相同,其中S1種群內,雌雄植株在0—0.45 m尺度上正相關,0.45—1.2 m尺度上彼此相互獨立,1.2—1.5 m尺度上彼此輕微正相關,其余尺度上彼此相互獨立,整體趨于不相關。整體尺度變化趨勢大致為:“正關聯→無關聯”的特點;S2種群內,雌雄植株在全部尺度內均為獨立或相關性極小。整體尺度變化趨勢大致為:“無關聯”的特點;S3種群內,雌雄植株在0—3 m尺度上為正相關,3—4.5 m尺度上為相互獨立或輕微正相關,4.5—6.3 m尺度上為彼此相互獨立,6.3—9 m尺度上為相互獨立或輕微負相關,9—15 m尺度上雌雄植株為相互獨立。整體尺度變化趨勢大致為:“正關聯→無關聯→負關聯→無關聯”的特點;S4種群內,雌雄植株在0—9m尺度上為正相關,9—10.5 m尺度上為相互獨立或輕微正相關,10.5—15 m尺度上彼此相互獨立。 整體尺度變化趨勢大致為:“正關聯→無關聯”的特點。
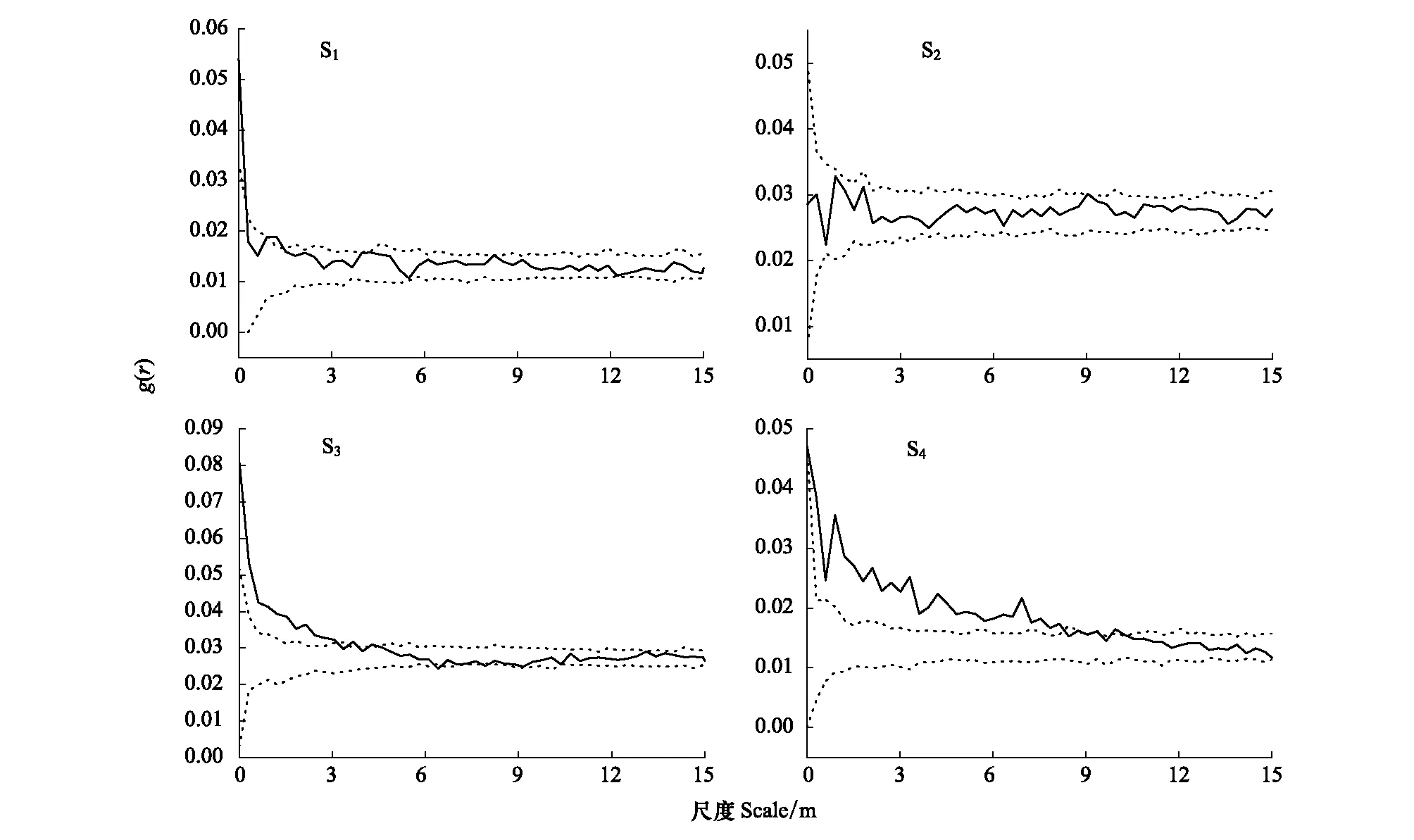
圖4 斑子麻黃種群雌雄植株間的空間關聯Fig.4 Spatial associations between male and female plants of E. rhytidosperma populations 實線為g(r)值,虛線為99%置信區間
2.6 斑子麻黃不同樣地的土壤指標
4個種群的土壤理化性質如表3所示,土壤含水量均值高海拔種群(S1、S2)高于低海拔種群(S3、S4),低海拔S4種群土壤含水量最低,S2與 S4之間差異顯著;土壤容重低海拔種群(S3、S4)高于高海拔種群(S1、S2),土壤容重均值S3最大,S3與S1差異顯著、S3與S2差異顯著;全氮高海拔種群(S1、S2)高于低海拔種群(S3、S4);土壤有機質含量高海拔種群(S1、S2)高于低海拔種群(S3、S4),S1最高。土壤全磷含量高海拔種群(S1、S2)高于低海拔種群(S3、S4),S1、S2、S3、S4之間差異不顯著;土壤pH值S3最大,S3與S1之間差異顯著。
3 討論
種群的空間分布、大小結構反映了種群的發展動態及趨勢,并在一定程度上揭示了種群的更新策略、地位以及與環境的相互關系[28—29]。4個斑子麻黃種群徑級分析顯示:S3、S4種群不同徑級的發展趨勢很相似,在成年階段呈現明顯的“波峰”現象,均有一定數量的幼體和成體植株,兩者總和占各自種群個體總數的78%、84%,幼樹和成樹居多,說明這2個種群自然更新良好,說明S3、S4種群在一定時期內表現出增長趨勢。S1、S2種群個體主要分布在成樹和老年兩個等級,其總和分別各自種群個體總數的92%和94%,說明種群缺乏幼體,導致種群天然更新力較差,表現出一定的衰退趨勢。在S1、S2種群中,成年、老年植株比例明顯上升,說明成年和老年階段的斑子麻黃成活率明顯提高了,生存能力更強。這與Bin研究的成年、老年植株個體生活史策略可能相對保守、生長相對緩慢,死亡率較低結果一致[30]。
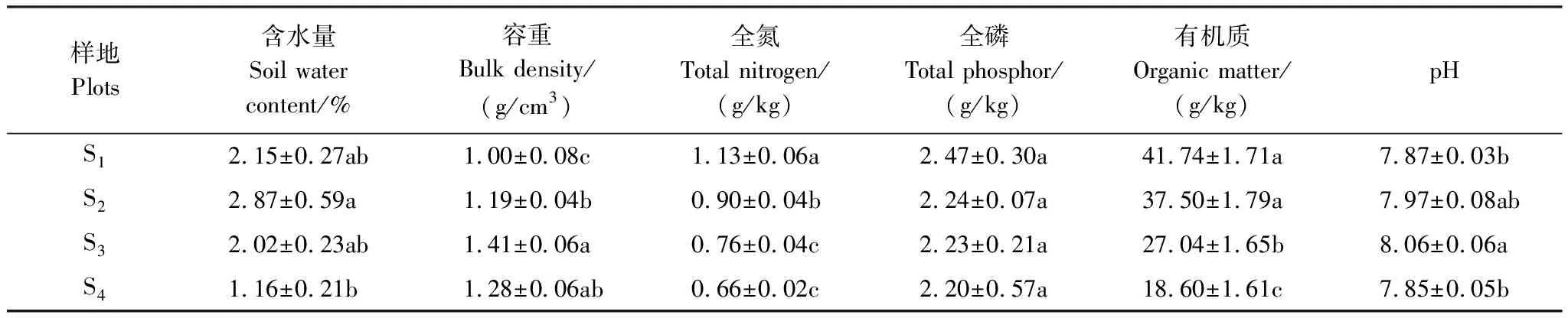
表3 各樣地土壤指標
雌雄異株植物普遍存在性別依賴的繁殖差異,雌株付出的繁殖代價通常比雄株要高。雌株較高的繁殖投資將導致雌株產生高死亡率,以致種群性比偏雄性[31—32]。本次調查的4個不同海拔的種群性比格局相似。S1、S2、S3、S44個種群分別都偏雄性。4個種群的總植株數雄株多于雌株,且性比極顯著偏離1∶1(P<0.01)這與很多雌雄異株植物的性比格局結果一致[33—35]。很多研究表明,環境因子能夠影響雌雄異株植物的性比,一般干旱或少養分環境條件下,雄株占優勢;而在濕潤或高養分環境條件下,雌株占優勢[34]。本研究中S1、S2樣地土壤全磷、全氮、有機質及土壤含水量明顯高于S3、S4樣地,S1、S2兩個種群性比格局不顯著偏倚1∶1(P>0.05);S3、S4整體性比格局顯著偏倚1∶1(P<0.05);這與Dawson等研究雌雄異株植物性比格局與環境因子異質性的研究結果一致[34]。
除去性別未定植株外,不同徑級之間種群的雌雄分布,S1和S2種群土壤水分、養分條件優于S3和S4種群。調查結果發現S2種群在幼年、成年階段均偏雌性,性比均顯著偏倚1∶1(P<0.05,P<0.001)。S1種群在成年階段也偏雌性,雖然S1種群在幼年階段沒有明顯偏雌性,但是雌雄植株數間僅差一株;結果表明S1、S2在幼年成年偏雌性。這與Dawson等、Freeman等研究的在濕潤或高養分環境條件下雌株占優勢結果一致[34,36]。隨著年齡的增加,在老年階段,S1、S2種群均明顯偏雄性,且兩個種群性比均極顯著偏倚1∶1(P<0.01、P<0.001)這可能由于雌株較高的繁殖投資造成死亡率高于雄株,雌株的生命周期明顯短于雄株,性比由原來的偏雌性逐漸過渡到偏雄性,這與Rocheleau等研究雌雄植株不同的繁殖投資成本將導致其生命周期的差異結果一致[37]。土壤環境較差的S3、S4種群幼年、成年、老年階段均偏雄性,且在幼年階段性比極顯著偏倚1∶1(P<0.01、P<0.01)。說明在干旱少雨的環境幼小的雄株比雌株具備更強的生命力。由于雌樹需要具有足夠的資源儲備才能開始有性繁殖,而雄樹只需相對較少的資源儲備,這導致雄樹在相對雌樹較小年齡就開始有性繁殖。小徑級范圍內性比的極顯著偏倚也導致了整體水平上性比偏雄性[13]。這與趙亞洲等的雌雄異株植物黃連木種群性比和空間分布結論一致[38]。
對于斑子麻黃雌雄植株間的空間關聯研究發現,4個不同海拔分布種群的空間關聯性不一致:S1、S3、S4大致表現為不同尺度的“正關聯→無關聯”,但S1種群雌雄之間的正關聯(0—0.45 m)遠遠小于S3、S4種群雌雄之間的正關聯(0—3 m)、(0—9 m),以致S3、S4種群為增長型種群,S1種群為衰退型種群;由此可見,較大尺度上的雌雄正關聯有利于雌雄異株植物完成傳粉,有利于繁殖。而S2種群在整個尺度上都表現為“無關聯”,即為衰退種群。說明雌雄之間空間隔離不利于雌雄異株植物繁殖。S2種群雌雄株間可能為了能更好地適應微環境,產生雌雄植株在空間占有和資源利用上的差異性,一定程度上導致空間分離[38]。這種空間分離雖然有利于斑子麻黃雌雄植株對資源和空間的充分利用,但形成性別間的空間隔離。且不利于種群繁殖,表現為S2種群出現衰退現象。S1、S2種群這種衰退現象足以說明雌雄之間的空間關聯性不夠或雌雄之間無關聯不利于種群繁殖。
本研究結果揭示了斑子麻黃種群的發展動態與環境因子以及與植株的年齡結構、性比、雌雄空間分布格局的關聯性,將為斑子麻黃種群保護和恢復提供理論依據。
猜你喜歡
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:30
當代陜西(2021年17期)2021-11-06 03:21:36
原道(2020年2期)2020-12-21 05:47:06
當代陜西(2019年15期)2019-09-02 01:52:00
中國非營利評論(2018年2期)2018-06-18 10:48:50
學苑創造·A版(2018年11期)2018-02-01 06:29:20
自動化學報(2017年1期)2017-03-11 17:31:17
讀者(2017年5期)2017-02-15 18:04:18
西藏科技(2016年5期)2016-09-26 12:16:39
振動工程學報(2015年1期)2015-03-01 01:15:42
