不同致病力松材線蟲對紅松早期防御酶活性影響*
2022-12-23 09:04:08曹業凡王曦茁汪來發蘇勝榮
林業科學 2022年8期
曹業凡 王曦茁 汪來發 汪 祥 徐 明 蘇勝榮 郭 偉
(1.中國林業科學研究院森林生態環境與自然保護研究所 國家林業和草原局森林保護學重點實驗室 北京 100091; 2.江蘇省林業科學研究院 南京 211153; 3.黃山學院生命與環境科學學院 黃山 245041; 4.吉林省林業和草原局 長春 130000)
松材線蟲病又稱松樹萎蔫病,是一種以松材線蟲(Bursaphelenchusxylophilus)為病原,墨天牛屬(Monochamus)昆蟲為媒介的系統性病害(葉建仁, 2019; Futai, 2013),主要危害松科植物,目前該病害已蔓延擴散至中國、日本、韓國、葡萄牙和西班牙等多個國家(Wangetal., 2010;Abelleiraetal., 2011;Shinyaetal., 2013)。據國家林業和草原局2022年6號公告統計,目前我國大陸地區松材線蟲病縣級疫區數為731個,新增縣級疫區22個,病情由南向北不斷蔓延擴散,形勢嚴峻。松材線蟲的寄主植物分布廣泛,包括自然感病寄主與人工接種感病寄主在內共計106種松科(Pinaceae)植物,其中大部分屬于松屬(Pinus),主要包括黑松(P.thunbergii)、馬尾松(P.massoniana)、思茅松(P.kesiyavar.langbianensis)、油松(P.tabulaeformis)等(葉建仁, 2010; Shietal., 2012)。韓國于2006年最早發現紅松(P.koraiensis)松材線蟲病(Hanetal., 2006),經科赫氏法則驗證確認紅松為松材線蟲病新寄主,我國2017年后相繼發現紅松感染松材線蟲病(于海英等, 2018; 潘龍等, 2019)。
當植物處于逆境脅迫環境時,會產生過多活性氧(ROS)。當活性氧累積過量時,會導致植物受到過氧化傷害,因此植物在逆境脅迫下抗氧化系統會被激活并分泌大量抗氧化酶,進而抵御逆境帶來的氧化傷害(Cha-UmSetal., 2009)。超氧化物歧化酶(superoxide dismutase, SOD)、過氧化氫酶(catalase, CAT)、苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)與過氧化物酶(peroxidase, POD)是ROS水解過程中重要的4種氧化酶,參與活性氧清除及酚類、木質素和植保素等抗病相關物質的合成,能抵御活性氧及氧自由基對細胞膜系統的傷害,使植物抵抗病害的能力增強,是植物體內重要的防御酶(何利等, 2013)。謝婉鳳等(2018)研究發現,2年生馬尾松苗接種松材線蟲后SOD與POD活性下降,且相關基因的表達也受到影響。滕濤等(2013)通過對1年生不同抗性種源馬尾松進行線蟲接種并進行防御酶活性測定發現,抗性種源POD活性均大于易感種源,表明POD活性動態變化與不同地理種源馬尾松的抗病能力有關。秦世杰等(2021)通過對自然感病狀態下的油松進行防御酶活性測定發現,感病油松PAL與CAT活性與對照組差異極顯著,但目前對紅松感染松材線蟲后防御系統的變化研究鮮見報道。鑒于此,本研究通過對接種不同株系松材線蟲后4年生紅松幼苗的早期相關防御酶進行活性測定,分析早期防御酶活性變化與同株系松材線蟲致病力的相關性,以期了解紅松感染松材線蟲后的生理生化反應,為研究松材線蟲的致病機制及相應寄主抗病特征提供參考。
1 材料與方法
1.1 試驗材料
松材線蟲株系號及其來源: 1) QH-1:采自沈陽清原的感病紅松; 2) NM-1:采自南京溧水的感病馬尾松; 3) CM-1:采自重慶的感病馬尾松。按常規方法分離純化和培養松材線蟲。通過貝爾曼漏斗法從疫木中分離得到線蟲后,接種于長滿灰葡萄孢(Botrytiscinerea)的PDA培養皿內,于25℃黑暗條件下培養線蟲。待培養皿內取食完畢后,通過貝爾曼漏斗法分離線蟲得到線蟲懸浮液,將其線蟲含量調整為20條·μL-1。
本試驗所用幼苗為4年生紅松實生苗,培養于中國林業科學研究院內苗圃地,備用。
1.2 試驗方法
1.2.1 接種線蟲 接種方法參照Qiu等(2016)方法。每個株系接種21株紅松幼苗,空白對照組接種21株幼苗。用消毒解剖刀在4年生紅松幼苗中部偏下約2cm位置,由上到下割開樹皮形成約0.5cm的縱切口,撕開韌皮部后放入大小適宜的棉球,使用封口膜將棉球包裹呈漏斗狀,通過手持移液器對棉球注入100 μL線蟲懸浮液,對照組注入等量無菌水。用透明膠蓋住漏斗上部便于保濕,接種完成后置于室外培育,定期澆水保濕。
1.2.2 樣品采集及預處理 接種完成后,持續1周對接種紅松幼苗進行取樣,取樣部位為接種點附近的當年生嫩枝針葉,每個時間點取生物學重復3組。樣品采集完成后,立即將樣品儲存于10 mL離心管內,將離心管置于液氮中浸泡1min后,儲存于-70℃冰箱保存備用。SOD、CAT、PAL與POD活性測定試劑盒由蘇州科銘生物技術有限公司提供,測定方法參照試劑盒說明書。
1.2.3 致病性測定 致病性測定接種方法參照1.2.1,每株系接種紅松幼苗10株,接種量為每株2 000條,以無菌水接種為對照。接種后,持續觀察接種幼苗發病情況,統計松苗發病率,感病指數與死亡率計算參照徐福元等(1994)并稍作改進。接種35天后,統計各株系松苗發病率,對所有植株分別進行線蟲分離與計數統計。完成致病性測定后,對不同株系松材線蟲進行致病力統計。
發病率=發病株數/接種株數×100%。
感病指數=∑(各級株數×各級代表數值)×100/調查總株數×發病最重級的代表數值。
平均發病等級=∑(各級株數×各級代表數值)/調查總株數
1.2.4 防御酶活性變化幅度與致病力相關性分析 SOD、CAT、PAL與POD活性最大變化幅度測定方法參照1.2.2,致病力統計結果參照1.2.3中公式。對上述數據繪制折線圖,分析不同株系松材線蟲與不同防御酶活性變化幅度相關性。
1.3 數據處理
采用SPSS 13.0進行差異顯著性分析及Ducan多重性比較,繪圖軟件采用Sigmaplot 14.0。
2 結果與分析
2.1 SOD活性變化
SOD活性變化結果表明,接種2天后,除松材線蟲株系CM-1處理與空白對照組外,其他處理組紅松幼苗SOD活性均出現小幅度上升(圖1),隨后2~4天內,所有處理組SOD活性均有不同程度下降,其中QH-1處理組SOD活性下降幅度最大。接種第5天后,QH-1處理組SOD活性開始大幅度上升,隨后SOD活性持續下降直至接種7天后達到最小值,而NM-1與CM-1處理組SOD活性在接種5天后均呈下降趨勢,隨后NM-1處理組SOD活性呈先上升后下降趨勢,CM-1呈先下降后上升趨勢,無明顯變化。接種7天后,QH-1、NM-1和CM-1處理組與CK組相比分別下降36.7%、17.7%和12.1%。因此,紅松幼苗接種QH-1株系線蟲后SOD活性變化幅度最大。

圖1 接種不同株系松材線蟲后的紅松SOD活性變化
2.2 CAT活性變化
接種2天后,除QH-1處理組與CK組紅松幼苗CAT活性呈小幅上升趨勢外,其他處理組幼苗CAT活性出現小幅度下降(圖2)。隨后2~4天內,NM-1與CM-1處理組CAT活性均呈先上升后下降變化趨勢,而QH-1處理組CAT活性呈先下降后上升變化趨勢。接種第5天后,除CK組外所有線蟲處理組CAT活性呈小幅度下降。接種6~7天后,所有線蟲處理組CAT活性呈先上升后下降趨勢變化,其中QH-1處理組活性最低,與CK組相比下降幅度為52.1%,而NM-1、CM-1處理組CAT活性與CK組相比分別上升10.6%、7.5%。因此,紅松幼苗接種QH-1株系線蟲后CAT活性下降幅度最大。
2.3 POD活性變化
接種2天后,所有線蟲處理組紅松幼苗與CK組相比POD活性呈上升趨勢變化,這表明對松材線蟲感染有積極的響應作用。其中QH-1處理組POD活性最高(圖3),CM-1處理組POD活性達到最高值。接種3天后,QH-1與NM-1處理組POD活性達到最高值。接種4~6天內,所有處理組POD活性均呈先下降后上升趨勢變化,其中QH-1處理組POD活性下降幅度最大。接種第5天后,除CK組外所有線蟲處理組POD活性呈小幅度下升。接種6~7天后,所有線蟲處理組POD活性呈先下降趨勢變化,其中QH-1處理組活性最低。接種7天后QH-1、NM-1和CM-1處理組與CK組相比分別上升3.3%、21.9%和16.3%,與最大值相比分別下降12.8%,33.5%和10.1%。
2.4 PAL活性變化
接種2天后,所有線蟲處理組紅松幼苗PAL活性均高于與CK組,此時QH-1處理組PAL活性最高(圖4),NM-1處理組活性最低。接種3~6天后,NM-1與CM-1處理組PAL活性雖有波動但無明顯變化,而QH-1處理組PAL活性呈先下降后上升趨勢變化,且在接種第5天后達到最小值。接種7天后,所有處理組PAL活性均高于CK組,其中QH-1處理組與CK組相比上升111%,NM-1、CM-1處理組與CK組相比分別上升46.3%、46.9%。
2.5 不同株系松材線蟲對紅松的致病力
接種試驗表明,QH-1、NM-1與CM-1株系松材線蟲均能使4年生紅松幼苗枯萎發病(表1)。癥狀觀察結果表明,感病初期接種點附近針葉開始褪綠變黃,感病中期整株針葉變黃,其中接種點附近針葉開始變紅。感病后期整株針葉枯萎變紅并停止流膠,此時接種幼苗已枯萎死亡(圖5)。

圖2 接種不同株系松材線蟲后的紅松CAT活性變化
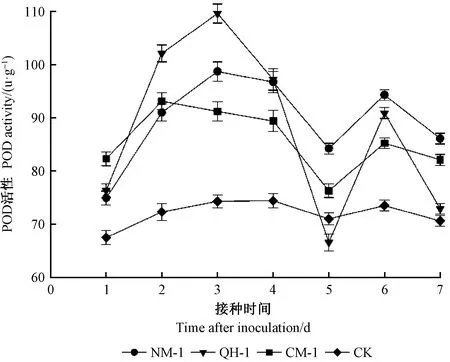
圖3 接種不同株系松材線蟲后的紅松POD活性變化
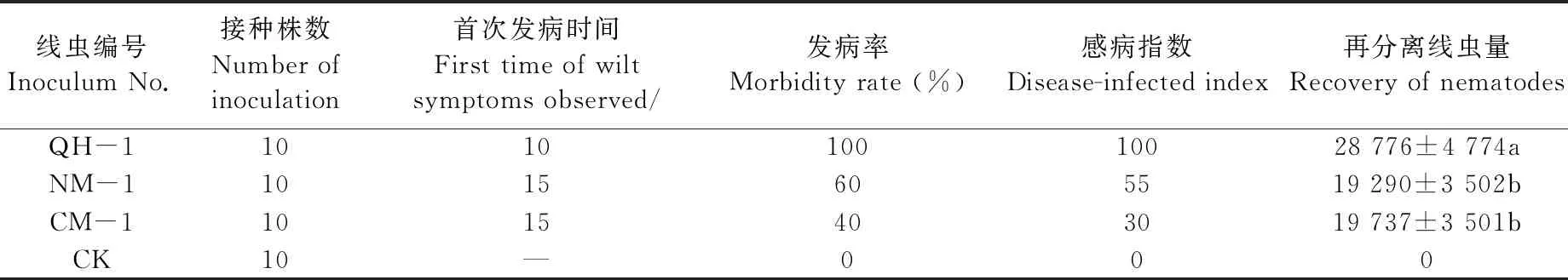
表1 4年生紅松幼苗接種松材線蟲的發病率、感病指數與再分離線蟲量(接種后35天)①
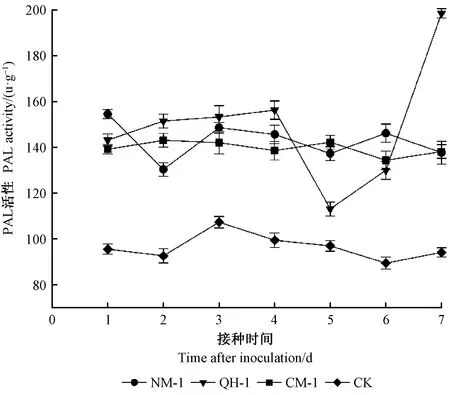
圖4 接種不同株系松材線蟲后的紅松PAL活性變化

圖5 接種松材線蟲后的紅松外部病狀
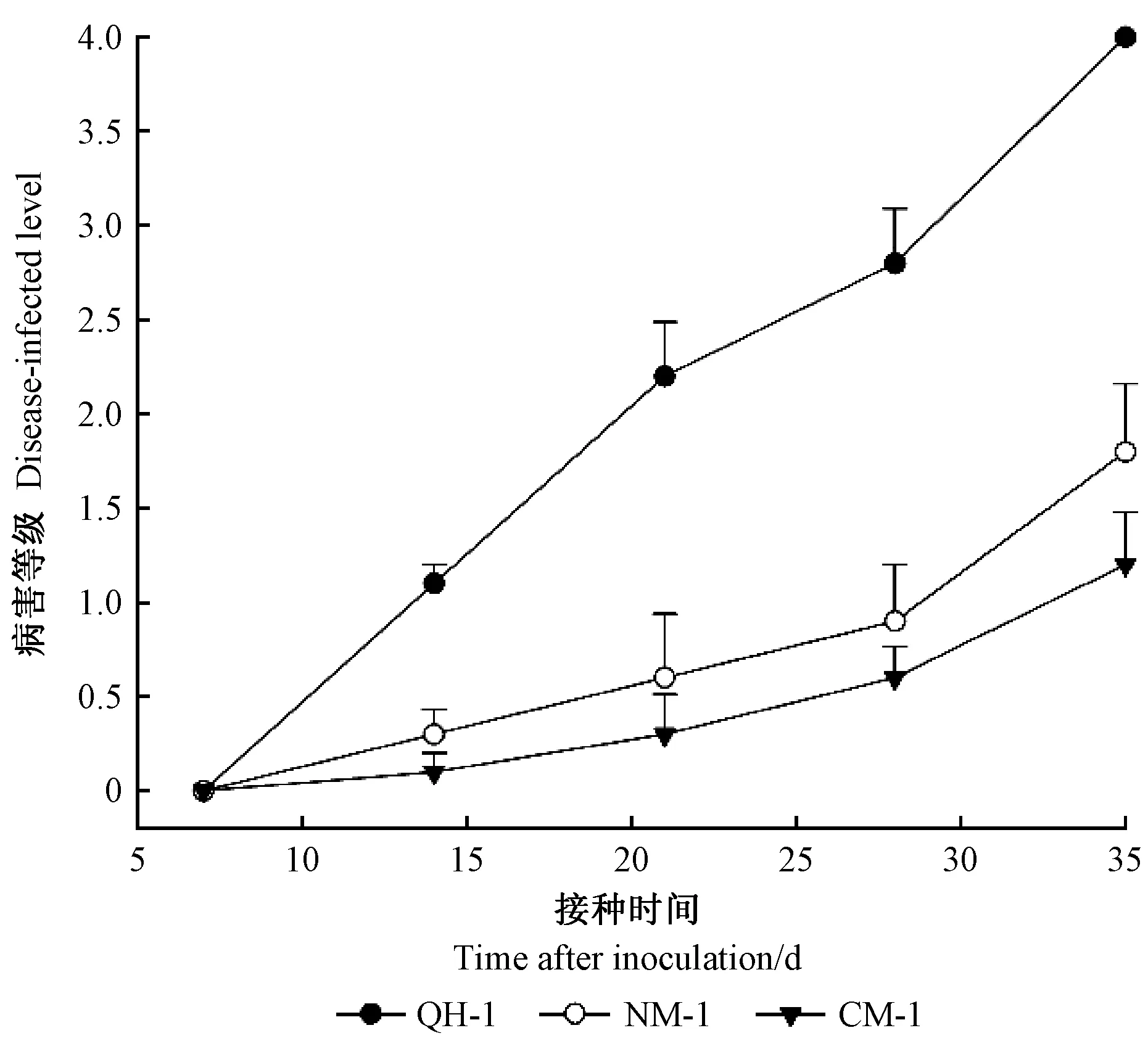
圖6 接種不同株系松材線蟲后紅松幼苗平均發病等級變化
病程記錄結果表明,不同株系松材線蟲接種紅松后的首次發病時間與致病進程存在明顯差異(表1)。接種QH-1的幼苗在10天時針葉開始出現褪綠黃變,于20天時出現針葉枯萎變紅,而接種NM-1和CM-1的幼苗于15天左右才開始出現針葉褪綠變黃癥狀,30天時出現針葉枯萎變紅(圖5)。發病率統計結果表明,接種5周后,接種QH-1的紅松幼苗發病率為100%,接種NM-1和CM-1的發病率分別為60%和40%。感病指數統計結果表明,接種QH-1株系松材線蟲2年生紅松幼苗感病指數為100,接種NM-1和CM-1的紅松幼苗感病指數分別為55和30。平均發病等級變化結果表明,接種35天后接種QH-1發病等級最高,此時接種QH-1紅松幼苗全部萎蔫死亡,而接種NM-1與CM-1紅松幼苗平均發病等級未達到4,尚未全部死亡(圖6)。
接種35天后萎蔫植株中的再分離線蟲量表明(表1),在接種QH-1、NM-1和CM-1后枯萎發病的紅松幼苗內均能分離到松材線蟲,在接種QH-1后枯萎的紅松幼苗平均每株分離的線蟲量為(28 776±4 774)條,而接種NM-1與CM-1后枯萎的紅松幼苗平均每株分離的線蟲量分別為(19 290±3 502)條和(19 737±3 501)條,數量差異顯著(P<0.1)。
致病力統計結果表明,QH-1、NM-1和CM-1株系松材線蟲對4年生紅松幼苗致病力存在差異,來自相同寄主的松材線蟲株系QH-1對紅松的致病力要大于來自馬尾松的松材線蟲株系NM-1和CM-1。
2.6 紅松接種不同株系松材線蟲后的酶活變化幅度
由圖7可知,紅松幼苗接種不同致病力株系線蟲后酶活性變化幅度存在差異。QH-1處理組防御酶活性變化幅度由大到小依次為PAL、CAT、SOD與POD,最大變化幅度為117%。NM-1處理組防御酶活性變化幅度由大到小依次為PAL、SOD、CAT與POD,最大變化幅度為46.3%。CM-1處理組防御酶活性變化幅度由大到小依次為PAL、POD、SOD與CAT,最大變化幅度為46.9%。所有線蟲處理組中PAL活性變化幅度最大,且接種強致病力松材線蟲株系QH-1后,感病紅松防御酶活性變化幅度最大。

圖7 接種不同株系松材線蟲后紅松幼苗防御酶活性變化幅度
3 討論
本研究發現,4年生紅松在接種不同株系松材線蟲后的防御酶活性變化存在明顯差異,結合后期致病性統計結果,發現接種強致病力株系松材線蟲后,接種幼苗防御酶活性變化幅度大于弱致病力處理組酶活性變化幅度。因此,通過測定接種植株的早期防御酶活性變化,可以從側面反映不同株系松材線蟲致病力強弱,致病力強弱與寄主感病早期防御酶活性變化存在一定相關性。
3.1 松材線蟲對接種紅松幼苗防御活性影響
SOD主要通過催化ROS歧化反應,使得ROS生成H2O2和O2,進而保護植物避免或減輕ROS損傷(夏民旋等, 2015)。Song等(2015)通過利用黃瓜花葉病毒(cucumber mosaic virus, CMV)侵染黃瓜和西紅柿(Solanumlycopersicum)葉片,發現兩者葉片中SOD的活性均上升,表明CMV通過激活黃瓜(Cucumissativus)防御酶系統,導致SOD活性上升。謝婉鳳等(2018)通過對接種松材線蟲后馬尾松進行SOD活力測定,發現SOD活力比對照組顯著下降。因此,當植物處于逆境脅迫時,SOD活性會出現不同程度的上升與下降,這可能與該植物抗病性與感病程度相關。本研究中,所有線蟲處理組SOD活性在接種7天后均顯著低于對照組,說明紅松幼苗感染線蟲后,SOD活性降低導致病害程度加深,且QH-1處理組活性下降程度最大,后續病害統計結果也顯示QH-1對4年生紅松致病力最強。因此,QH-1處理組SOD活力大幅下降可能是因為QH-1對接種植株致病力最強,進而加快接種植株發病進程。
CAT和POD是分解H2O2的主要氧化酶類。當植株由于ROS反應產生大量H2O2后,CAT和POD能夠將H2O2分解為H2O,防止因自身免疫反應造成細胞損傷。因此,CAT與POD在植物抗病過程中作用重大。陳玉蓮等(2015)通過測定接種松材線蟲后不同家系馬尾松防御酶活性發現,CAT與POD活性變化趨勢與松材線蟲病的感病指數相關,且活性在接種后都表現出先上升后下降的趨勢。劉洪劍等(2018)通過對不同抗性家系馬尾松接種松材線蟲并測定防御酶酶活性變化發現,CAT與POD變化趨勢與陳玉蓮等(2015)研究結果相似。本研究中所有線蟲處理組CAT與POD活性在接種線蟲1天后,均高于CK組,可能是由于體內高質量濃度的H2O2對酶促反應起到正調控的作用,此時接種植株抗病反應已被激活。接種7天后,QH-1處理組CAT活性大幅度下降,而其他處理組無明顯變化,可能是因為QH-1處理組接種幼苗此時因ROS過量導致細胞損傷,進而無法繼續產生CAT分解過量H2O2,這與SOD中的數據變化相似。后續致病力測定表明QH-1為強致病力松材線蟲株系,說明通過早期CAT活性變化能夠在一定程度上驗證接種致病力強弱。本研究中,所有線蟲處理組POD活性均呈先上升再下降的動態變化,接種1~3天內所有線蟲處理組POD活性均高于對照組,之后開始呈下降趨勢,表明此時接種植株抗病反應過度,導致POD活性下降。因此,在早期接種植株抗病過程中,通過CAT與POD活性測定能夠在一定程度上反映后期發病程度,做到病害預測,減少經濟損失。
當植物被病原物侵入時,寄主植物通過苯丙烷代謝途徑抵御病害,此時PAL含量升高。當PAL活性升高后,其代謝產物的不斷積累將會導致寄主植物的代謝紊亂,造成感病植株死亡(歐陽光察等, 1988)。陳玉惠等(2006)通過對1年半生馬尾松植株進行松材線蟲接種,并測定接種后植株防御酶活性,發現PAL活性高于對照,且活性高峰出現于外部病癥表現前,并在感病后期下降。本研究中,所有線蟲處理組在接種2天后,接種植株PAL活性均高于與CK組,與前人研究結果一致。接種7天后,所有處理組PAL活性均高于CK組,其中QH-1處理組與CK組相比活性上升幅度最大,且差異顯著,表明強致病力株系與弱致病力株系相比,更能破壞接種紅松內部生理活動,造成PAL活性升高。因此,通過測定早期PAL活性,更能反映感病松樹的抗病反應程度。在本研究中,所有線蟲處理組防御酶中,PAL活性變化幅度遠大于其他防御酶,也說明在接種紅松在發病初期,由PAL主導的生理生化反應響應最迅速,PAL活性變化幅度在一定程度上能夠及時預測病原物對寄主致病力強弱。
3.2 株系差異對松材線蟲致病力影響
不同株系松材線蟲致病性強弱主要與松材線蟲的地理來源、寄主植物、寄主樹齡與飼喂菌株有一定關系(胡凱基等, 1994; 張治宇等, 2002; 楊寶君等, 1993; 瞿紅葉等, 2009)。有研究發現,南方株系對黑松致病性強于北方株系,且北方株系對北方易感樹種油松致病性大于南方株系(Kongetal., 2021),與本研究致病性研究結果相似。本研究中所選松材線蟲其地理來源與寄主植物分別為清原-紅松(QH-1)、南京-馬尾松(NM-1)與重慶-馬尾松(CM-1),因此本研究中不同株系松材線蟲對4年生紅松幼苗致病性存在差異,可能與地理分布與寄主植物有關。結合相關防御酶活性變化結果表明,4年生紅松幼苗在接種強致病力松材線蟲后,與接種弱致病力株系松材線蟲的4年生紅松幼苗相比,酶活性變化幅度更大。通過接種線蟲并測定早期防御酶活性,可以預測接種松材線蟲致病力強弱,至于其致病力差異的產生機制,有待進一步研究。
3.3 不同病原物對寄主御酶活性的影響
不同寄主病原物對寄主防御酶的活性影響存在一定差異。蔡麗等(2011)通過對橡膠(Heveabrasiliensis)樹葉接種不同寄主來源的多主棒孢(Corynesporacassiicola),發現接種葉片防御酶活性變化存在差異,且原寄主為橡膠樹的多主棒孢更容易激發接種植株防御酶活性變化,變化幅度最高,其他寄主的多主棒孢侵染橡膠樹后,酶活性升高幅度小。雍道敬等(2014)通過對蘋果葉片分別接種2種不同致病性的蘋果樹腐爛病菌(Valsamalivar.mali),發現接種強致病力菌株葉片防御酶活性變化幅度最早,但感病后期接種弱致病力菌株葉片防御酶活性峰值顯著大于強致病力組,因此強致病力病原物能夠加速寄主抗氧化反應失效,導致活性氧大量積累,加速寄主死亡,使得防御酶活性降低。本研究通過接種不同致病力松材線蟲株系發現,強致病力株系QH-1導致4年生紅松幼苗防御酶活性變化最早,變化幅度最大。接種7天后,QH-1處理組SOD、CAT與POD活性均低于弱致病力NM-1與CM-1處理組,與雍道敬等(2014)研究一致。接種35天后強致病力株系QH-1處理組線蟲分離量顯著大于其他處理組,這可能是因為QH-1株系線蟲在早期侵入樹體階段,更早更快地破壞了接種幼苗樹體內部抗氧化系統,進而能夠大量繁殖。
4 結論
松材線蟲株系QH-1、NM-1和CM-1對紅松均有致病性,其中QH-1致病力最強,NM-1、CM-1無明顯差異。防御酶活性結果表明,接種QH-1后的紅松幼苗在發病早期SOD、CAT、POD與PAL變化幅度大于其他線蟲處理組,且致病力強弱與防御酶活性變化幅度有一定相關性。綜上所述,QH-1、NM-1與CM-1對4年生紅松存在致病力差異,且致病力差異與寄主早期防御酶活性變化幅度有關。
