山桐子健康和感病植株不同器官區細菌群落結構、多樣性與功能預測分析*
2022-12-23 09:04:24薛曉焱蔡齊飛耿曉東周慧娜李明婉王艷梅
林業科學 2022年8期
李 志 薛曉焱 劉 震 蔡齊飛 耿曉東 馮 建 周慧娜 張 濤 李明婉 王艷梅
(河南農業大學林學院 鄭州 450002)
山桐子(Idesiapolycarpa)是大風子科(Flacourtiaceae)山桐子屬落葉闊葉喬木,樹形優美,木材松軟,果實艷麗,在不同立地環境均具有較好的適應性,是良好的新型用材樹種和觀賞樹種(陳珺肄等, 2019)。山桐子果實含油率高且營養物質豐富,其果肉和種子中不飽和脂肪酸含量可達到75%和85%以上,是優良的木本油料樹種,可作為開發高檔保健型木本食用油的原料,素有“樹上油庫”的美譽(代莉, 2014)。山桐子作為木本油料樹種,具有抗逆性強、適生地廣、“不與糧爭地”的優勢,在林業、食品、醫藥及生態等多個領域有巨大的開發前景(陳釗等, 2019; 房麗莎等, 2020)。近年來,國家正式制定頒布了山桐子、山桐子油、山桐子餅粕3個行業標準,并將山桐子納入到了國家儲備林建設的政策體系中。
在自然界,植物生長環境中有大量微生物,寄主在植物宿主體內或附著其表面,這些微生物群體統稱為植物微生物組。細菌是這些微生物群落中最豐富的類群(Mülleretal., 2016)。同種植物不同器官區微生物存在一定差異,李聰聰等(2020)比較了高寒草甸優勢植物葉內、根內、土壤原核微生物組的組成,發現植物器官對微生物具有強烈選擇性,只有部分土壤微生物能夠進入植物根內和葉內; Wu等(2020)研究結果表明柑桔(Citrusreticulata)葉片和韌皮部的微生物群落數量遠低于根系和土壤。細菌與植物形成復雜、動態的關聯,包括互利共生或致病,Zarraonaindia等(2015)的研究發現葡萄(Vitisvinifera)葉子和果實中的甲基桿菌(Methylobacteriumspp.)可以通過產生植物激素的生產來刺激植物的發育; 尹詩琳等(2021)的研究發現,芽孢桿菌屬(Bacillus)對桑樹(Morusalba)病原菌存在著某種程度的拮抗作用,而泛菌屬(Pantoea)可能對桑輪紋病的發生有協同或者伴生作用。
山桐子林木出現感病現象,部分植株根系變黑腐爛,樹皮出現褐色水漬病斑,葉片邊緣枯萎且葉面有褐色斑點,病害發生原因及致病因子尚未明確。目前,山桐子相關研究主要集中在栽培技術、油脂提取、開發利用等方面,而微生物病害方面的研究相對薄弱。 李佛生等(2021)對山桐子內生真菌進行分離,鑒定出4種山桐子組織來源共同的內生真菌為芽短梗霉(Aureobasidiumpullulans); 有研究報道了山桐子葉斑病的病原真菌鏈格孢菌(Alternariaalternata)(Sunetal., 2015)。 細菌是否為造成山桐子病害的關鍵因子還有待進一步探索。研究基于高通量測序技術,以健康和感病山桐子植株為對象,探究山桐子土壤、樹皮及葉片中細菌群落組成和多樣性,分析其差異,探討健康和感病山桐子不同器官區的優勢菌屬及其潛在功能,探究各器官區的關鍵菌屬與山桐子感病之間的關系,以期為山桐子病害防治及經營管理提供理論基礎。
1 材料與方法
1.1 試驗地概況
研究試驗地位于河南農業大學林業試驗站,屬于暖溫帶大陸性氣候,年平均氣溫14.2℃,極端最高氣溫43℃,極端最低氣溫-17.9℃,年平均降水量650.1mm,全年日照時數約2 400 h。試驗地0~20cm土壤的基礎化學性質為: pH 8.0,土壤全碳(TC)17.76 g·kg-1,全氮(TN)0.81 g·kg-1,有效磷(AP)2.43 mg·kg-1,有效鉀(AK)75.78 mg·kg-1。
1.2 樣品采集
2020年5月,在樣地內采集健康山桐子植株和感病山桐子植株的根際土壤、樹皮、葉片樣品,以及樣地非根際土壤樣品。土壤樣品采集時,每樣株隨機選取3個樣點,去除植株基部表層土壤及雜物,取5~20cm土層側根或須根表面0.4cm左右的土壤作為根際土壤,空地同土層隨機取3個樣點作為非根際土壤,將每個樣點采集的土壤分別裝入無菌自封袋中充分混勻,共計9份土樣,土壤樣品編號為健康植株根際土壤(G)、感病植株根際土壤(BG)、非根際土壤(FG)。用無菌剪刀和鑷子采集植物樹皮和葉片,每樣株2個部位分別采3份,共6份皮樣、6份葉樣,編號分別為健康植株樹皮(P)、感病植株樹皮(BP)、健康植株葉片(Y)和感病植株葉片(BY)。采集好的樣品用冰盒保存帶回實驗室后,置于-80℃冰箱保存,供檢測使用。
1.3 測定指標及方法
對健康、感病山桐子植株根際土壤、樹皮、葉片及非根際土的總DNA進行提取,提取的DNA用1.2%瓊脂糖凝膠進行電泳,用紫外分光光度計檢測其質量和濃度。選擇引物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′),對樣品細菌16 S rRNA 基因的V3-V4 區進行PCR 擴增(Claessonetal., 2009)。采用的25μL擴增體系包括5×reaction buffer 5μL,5×GC buffer 5μL,dNTP(2.5mmol·L-1)2μL,Forwardprimer(10μmol·L-1)1μL,Reverseprimer(10μmol·L-1)1μL,DNA Template 2μL,ddH2O 8.75μL,Q5 DNA Polymerase 0.25μL。擴增參數為98℃ 2min,98℃ 15 s,55℃ 30 s,72℃ 30 s,72℃ 5min,30個循環,10℃保存。Illumina miseq測序平臺進行高通量測序,測序由上海派森諾生物科技有限公司完成。
1.4 數據處理
利用QIIME2(2019.4)軟件調用DADA2進行質控、去噪、拼接、去嵌合體,采用97%的序列相似度作為OTU(operational taxonomic unit,OTU)的劃分閾值,獲得的OTU代表序列用于細菌群落組成分析、α多樣性分析和β多樣性分析、菌群功能預測等; 運用R軟件等工具,繪制稀疏曲線; 根據ASV/OTU豐度表制作upsetR圖; 對樣本中相對豐度前20位的屬進行聚類分析并繪制熱圖; 根據樣本差異距離進行主坐標分析(Principal coordinate analysis,PCoA); 利用SPSS 24.0分析α多樣性指數在各樣本間的差異(P<0.05)。
2 結果與分析
2.1 微生物測序結果分析
對山桐子林地非根際土壤及健康、感病山桐子植株的根際土壤、樹皮、葉片進行高通量測序,21個樣本(3個重復/處理)中的細菌16 S rRNA V3-V4序列過濾后,共獲得1 193 103條高質量序列。稀疏曲線可以用來比較測序數量不同的樣本物種豐富度,也可說明樣本的取樣大小是否合理。根據樣本的測序數據,繪制對應的稀疏曲線(圖1)。結果表明,各樣本的稀疏曲線均趨于平緩,說明該測序深度包含了樣本中絕大多數細菌類型,測序深度充足,可以反映真實的細菌群落生物多樣性。
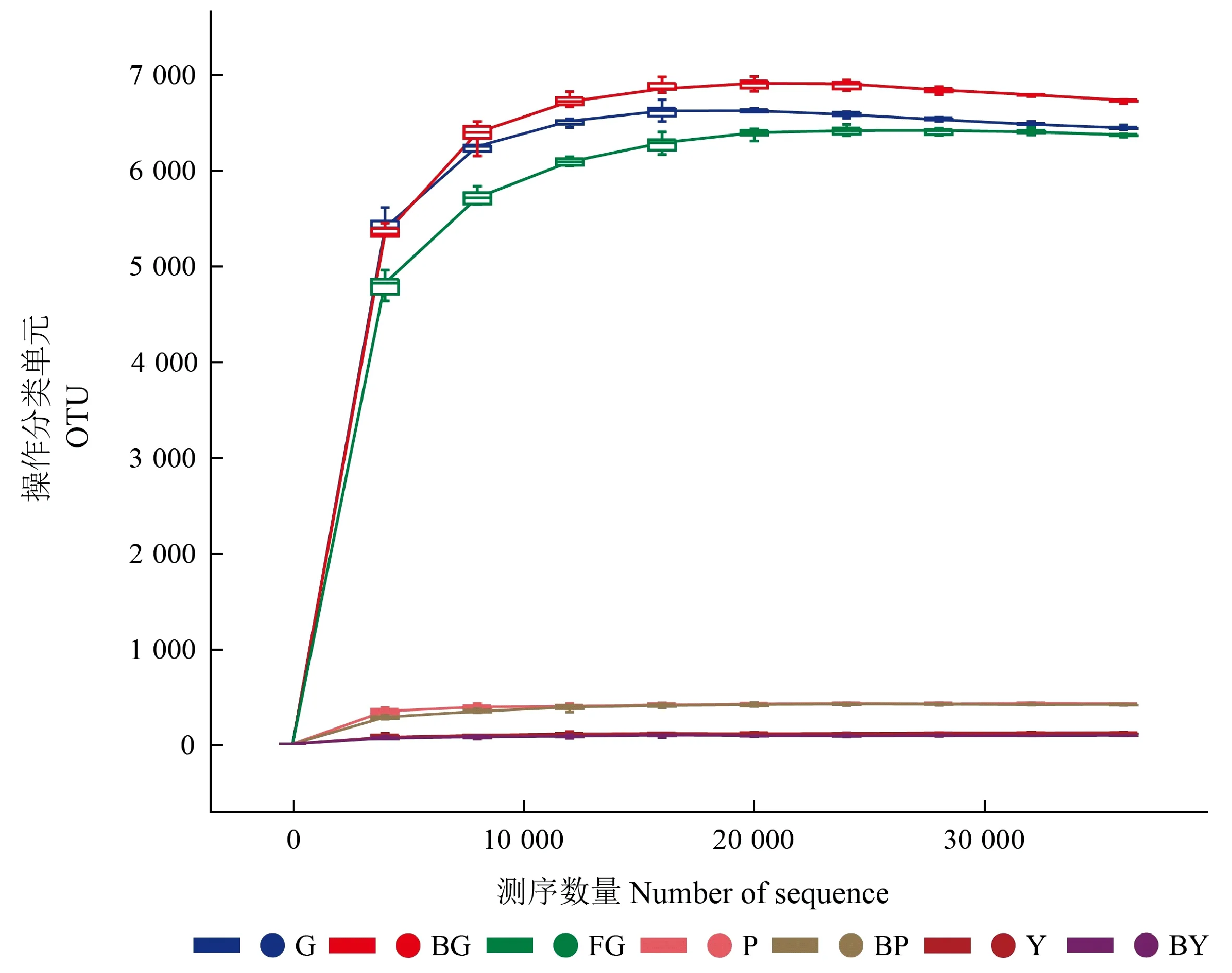
圖1 各區域樣本中細菌稀釋曲線
2.2 山桐子健康、感病植株不同器官區細菌群落結構
2.2.1 山桐子健康、感病植株不同器官區OTU數 使用upset圖分析山桐子不同器官區細菌的物種組成(圖2)。土壤樣本健康植株根際土壤感病植株根際土壤(G)、感病植株根際土壤(BG)、非根際土壤(FG)中包含的OTU 數目分別是17 236、17 403、15 461 個,3組共有OTU 數量為1 100個,各自特有OTU 數目分別是14 517、14 577、12 776個,根際土壤樣本特有的OTU 數均高于非根際土壤,預示著根際土壤中含有較多的特有微生物種類; G和BG兩組特有OTU分別占其總OTU的84.22%和83.76%。樹皮樣本P、BP中的特有OTU數量為737、857個,分別占其總OTU的74.29%和80.77%。葉片樣本Y、BY中特有OTU數量為181、156個,分別占其總OTU的69.35%和70.91%。以上數據表明,健康植株根際土壤和樹皮中特有OTU數量少于感病植株,葉片特有OTU數量多于感病植株,7組樣本中特有OTU數量均占較大比例。
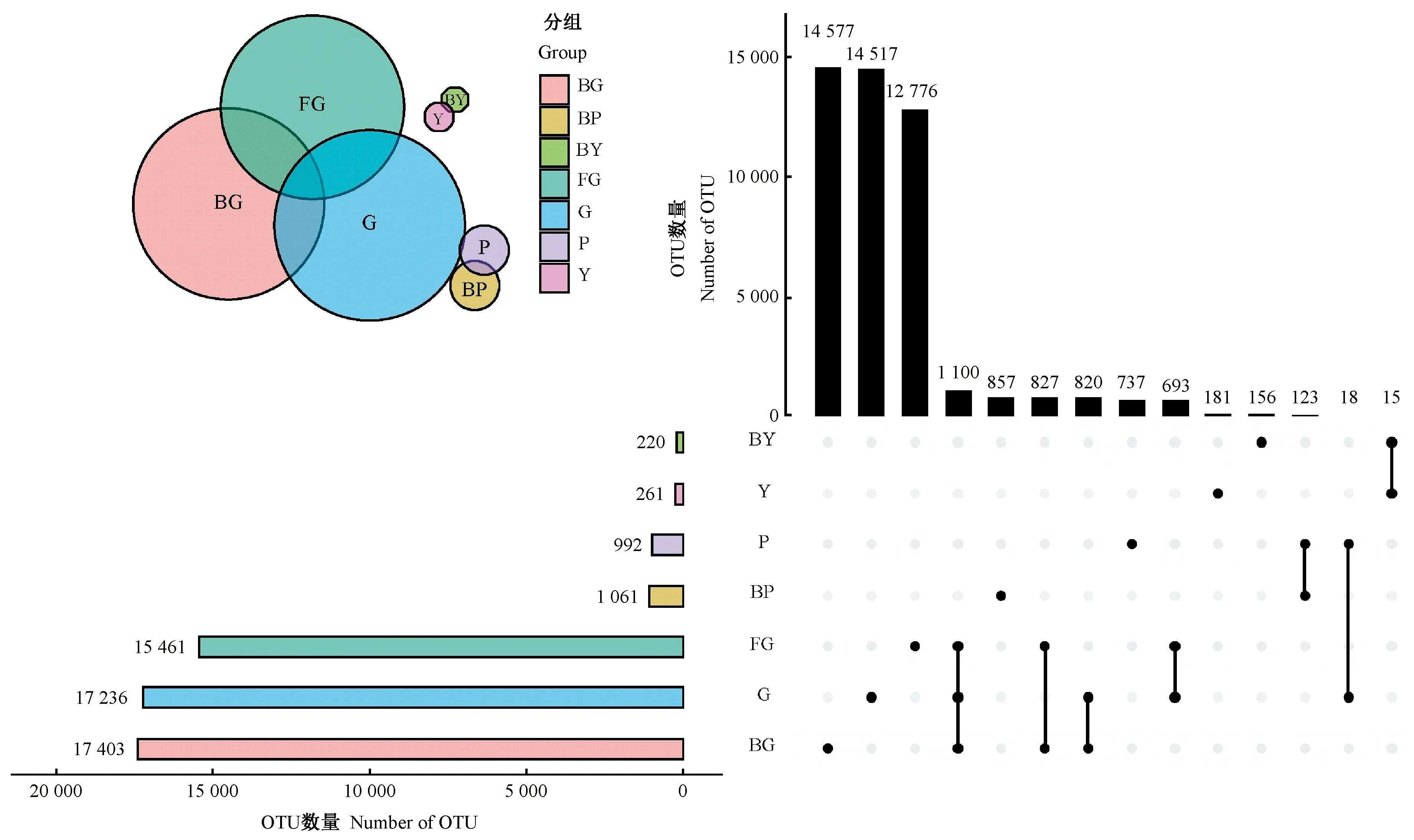
圖2 各器官區細菌群落
2.2.2 山桐子健康、感病植株不同器官區細菌門、綱、目、科水平群落結構 對各樣本中得到的OTU序列進行分類注釋,所有樣本的細菌OTU隸屬于37個門,118個綱,291個目,511個科,1051個屬,2772個種。
各分類水平上物種相對豐度前10位的細菌分布見圖3。在門水平上(圖3a),土壤樣本的優勢菌門為變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria); 其中變形菌門在G、BG和FG中所占比例為BG>G>FG,在根際土壤中相對豐度較大; FG中的酸桿菌門和綠彎菌門(Chloroflexi)占比數量顯著高于根際土壤(P<0.05),且綠彎菌門為FG的優勢菌門。樹皮、葉片樣本的優勢菌門為藍細菌門(Cyanobacteria)和變形菌門。在樹皮樣本中,變形菌門在P、BP中的相對豐度分別為30.52%、62.26%,P的相對豐度顯著低于BP; 藍細菌門在P、BP中的相對豐度分別為61.57%、28.39%,P的相對豐度顯著高于BP(P<0.05)。在葉片樣本中占比最多的是藍細菌門84.58%(Y)、88.42%(BY),其次是變形菌門15.16%(Y)、11.33%(BY),2個門的細菌占總數量的99.7%以上。
在綱水平上(圖3b),變形菌門中α-變形菌綱(Alphaproteobacteria)、γ-變形菌綱(Gammaproteo-bacteria)和放線菌門中放線菌鋼(Actinobacteria)在G和BG中占比較為接近,但均高于FG,其中放線菌綱在根際與非根際土壤中差異達到顯著水平(P<0.05); 而放線菌門中嗜熱油菌綱(Thermoleophilia)在FG中的相對豐度顯著高于根際土壤。在樹皮樣本中,藍細菌門中產氧光細菌綱(Oxyphotobacteria)在P(61.57%)中的相對豐度顯著高于BP(28.39%),γ-變形菌綱在BP(43.82%)中的相對豐度顯著高于P(5.15%)。綱水平上,葉片樣本主要綱為產氧光細菌綱和α-變形菌綱,兩者在Y和BY中占比接近。
在目(圖3c)、科(圖3d)水平上,產氧光細菌綱中葉綠體(Chloroplast)是樹皮、葉片的主要組成,在P、BP、Y、BY中占比分別為61.56%、28.39%、84.54%、88.41%; α-變形菌綱中立克次氏體目(Rickettsiales)及其下屬線粒體(Mitochondria)的相對豐度在P、Y、BY 3組樣本中占比分別為14.03%、14.06%和10.76%; γ-變形菌綱中腸桿菌目及其下屬腸桿菌科(Enterobacteriaceae)在樹皮樣本BP(41.03%)中的相對豐度顯著高于P(1.02%)。
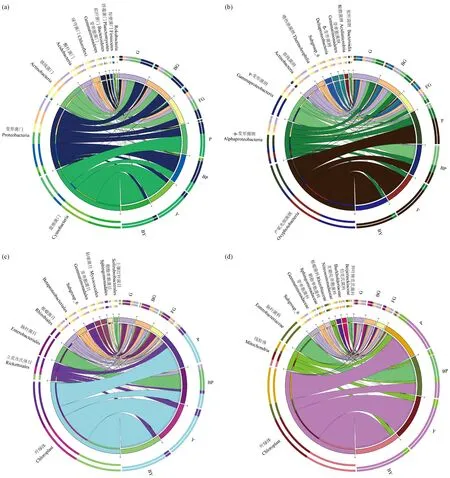
圖3 各分類水平上相對豐度前10物種在各樣本的分布和豐度
2.2.3 山桐子健康、感病植株不同器官區細菌屬水平群落結構 對3組土壤樣本中相對豐度排名前20位的細菌屬進行UPGMA聚類分析,結果表明(圖4),根際土壤G、BG具有相似的細菌群落結構,聚為一類; FG與根際土壤差異較大,單獨為一類。20個細菌屬可以分為3組(Ⅰ、Ⅱ和Ⅲ)。FG中Ⅰ組細菌屬豐度較高,主要包括變形菌門中的bacteriap25、Haliangium、MND1; 放線菌門中的Gaiella、MB-A2-108、67-14、IMCC26256; 綠彎菌門中的A4b、KD4-96; 酸桿菌門中的Subgroup_17、RB41; 以及浮霉菌門的OM190,Rokubacteria的Rokubacteriales,共13個菌屬。G中Ⅲ組細菌屬豐度較高,主要包括放線菌門中的類諾卡氏菌屬(Nocardioides)、芽球菌屬(Blastococcus)、土壤紅桿菌屬(Solirubrobacter); 酸桿菌門中的Subgroup_6; 變形菌門的鞘氨醇單胞菌屬(Sphingomonas),共5個菌屬。BG中Ⅱ組細菌屬豐度較高,主要包括變形菌門中Dongia以及放線菌門中的鏈霉菌屬(Streptomyces),共2個菌屬。
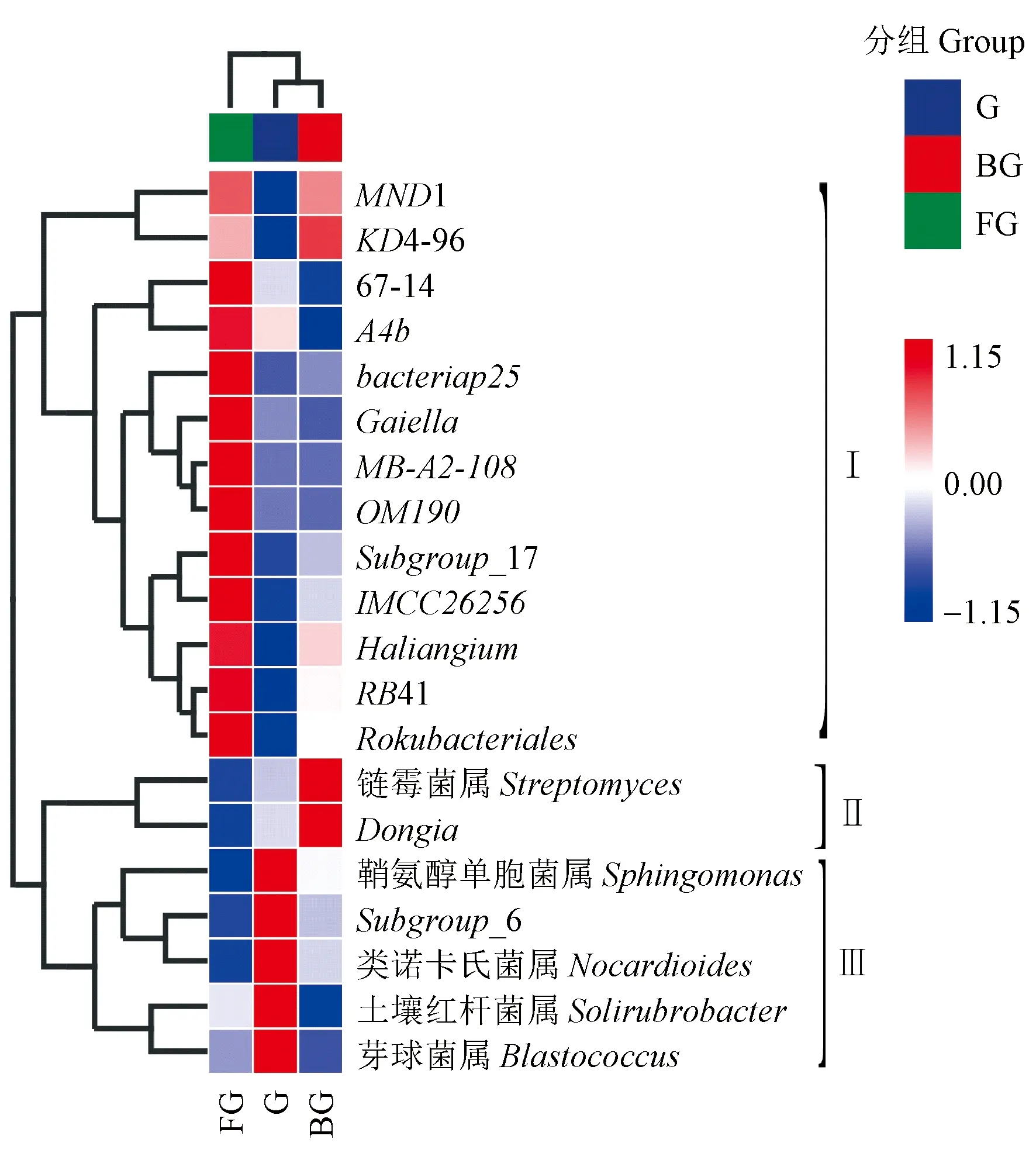
圖4 土壤在屬水平上的細菌豐度聚類熱圖
選取2組樹皮樣本中豐度排名為前20位細菌屬進行UPGMA聚類,繪制物種豐度聚類熱圖(圖5)。由圖5可知,2組樹皮中的細菌群落在屬水平上差異明顯。P中豐度較高的細菌屬,主要包括變形菌門中的鞘氨醇單胞菌屬、不動桿菌屬(Acinetobacter)、Aureimonas; 放線菌門中的Quadrisphaera、貧養桿菌屬(Modestobacter)和擬桿菌門的薄層菌屬(Hymenobacter),共6個菌屬。BP中豐度較高的細菌屬,主要包括變形菌門中的Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium、果膠桿菌屬(Pectobacterium)、甲基桿菌屬(Methylobacterium)、假單胞菌屬(Pseudomonas)、小坂菌屬(Kosakonia)、Brenneria、Hafnia-Obesumbacterium、玫瑰單胞菌屬(Roseomonas); 厚壁菌門中的類芽胞桿菌屬(Paenibacillus)、丹毒絲菌屬(Erysipelothrix)、Saccharibacillus; 放線菌門的芽球菌屬,共12個菌屬。
對樹皮樣本中的細菌進行優勢菌屬分析,在健康和感病山桐子樹皮細菌中,細菌豐度10%以上且存在顯著差異(P<0.05)的屬有2個,分別是Brenneria和Hafnia-Obesumbacterium,在健康山桐子樹皮中相對豐度僅為0.79%和0,顯著低于感病山桐子樹皮中Brenneria(18.33%)和Hafnia-Obesumbacterium(17.5%)的相對豐度。
選取2組葉片樣本中豐度排名為前20位細菌屬進行UPGMA聚類(圖6)。由圖6可知,Y中豐度較高的細菌屬,主要包括變形菌門中的鞘氨醇單胞菌屬、Burkholderia-Caballeronia-Paraburkholderia、短波單胞菌屬(Brevundimonas)、不動桿菌屬、慢生根瘤菌屬(Bradyrhizobium)、腸桿菌屬(Enterobacter)、Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium、假單胞菌屬、貪銅菌屬(Cupriavidus)、草螺菌屬(Herbaspirillum); 以及綠彎菌門的KD4-96,共11個菌屬。BY中豐度較高的細菌屬,主要包括厚壁菌門中的魏斯氏菌屬(Weissella)、芽孢桿菌屬、乳桿菌屬(Lactobacillus)、鏈球菌屬(Streptococcus); 以及變形菌門中的Escherichia-Shigella、Phreatobacter、Pusillimonas,共7個菌屬。
2.3 不同器官區細菌群落多樣性分析
2.3.1 α多樣性分析 對山桐子植株不同器官區細菌多樣性指數進行差異分析(表1)。采用Chao1指數反映物種豐富度; 采用Shannon指數和Simpson指數反映細菌群落的多樣性; Goods coverage指數反映群落覆蓋度。山桐子植株不同器官區細菌的Chao1、Shannon、Simpson指數值都表現為土壤樣本最高,其次是樹皮樣本,最后是葉片樣本,說明土壤細菌群落豐富度和多樣性是最高的。土壤各組的Chao1指數由大到小排列依次為: BG>G>FG,感病植株根際土壤細菌群落豐富度最高; Shannon指數和Simpson指數由大到小依次為: G>FG>BG,說明健康植株根際土壤細菌群落多樣性最高。葉片中Y組的Chao1、Shannon、Simpson指數均高于BY,表明健康植株葉片細菌群落豐富度和多樣性高于感病植株葉片。但健康植株與感病植株土壤、葉片細菌群落的Chao1指數、Shannon指數和Simpson指數差異均未達到顯著水平(P>0.05)。樹皮細菌群落Chao1指數表現為P高于BP,而Shannon、Simpson指數均表現為BP高于P,其中Simpson指數差異達到顯著水平(P<0.05),說明健康植株樹皮細菌群落豐富度較高,但細菌群落多樣性低于感病植株樹皮。

圖5 樹皮在屬水平上的細菌豐度聚類熱圖
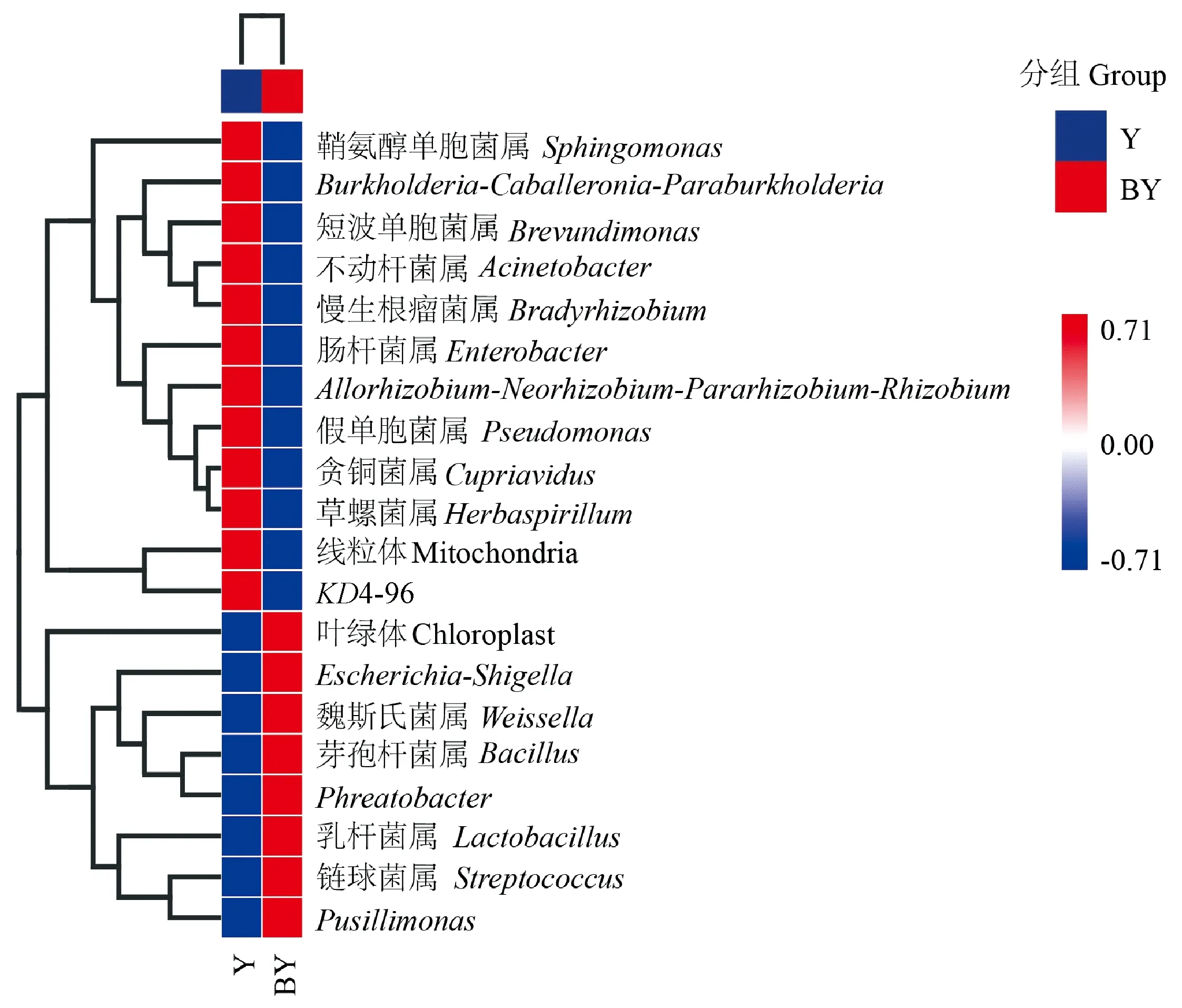
圖6 葉片在屬水平上的細菌豐度聚類熱圖
2.3.2 β多樣性分析 為進一步明晰樣本間在群落物種組成上的差異性,采用PCoA分析山桐子不同器官區細菌群落物種組成的相似度。如圖7所示,PCo1和PCo2分別解釋變量方差的42.9%、8.6%,兩者累計貢獻率達51.5%。PCo1可以將土壤的細菌群落與樹皮、葉片明顯區分開,土壤分布在PCo1負方向,樹皮和葉片主要分布在正方向,說明樹皮和葉片與土壤細菌群落差異較大。不同健康狀態山桐子的土壤和樹皮細菌群落結構存在差異性,其中健康、感病樹皮細菌組成差異明顯。而Y和BY相聚較近,表明健康和感病山桐子葉片細菌組成相似度較高。
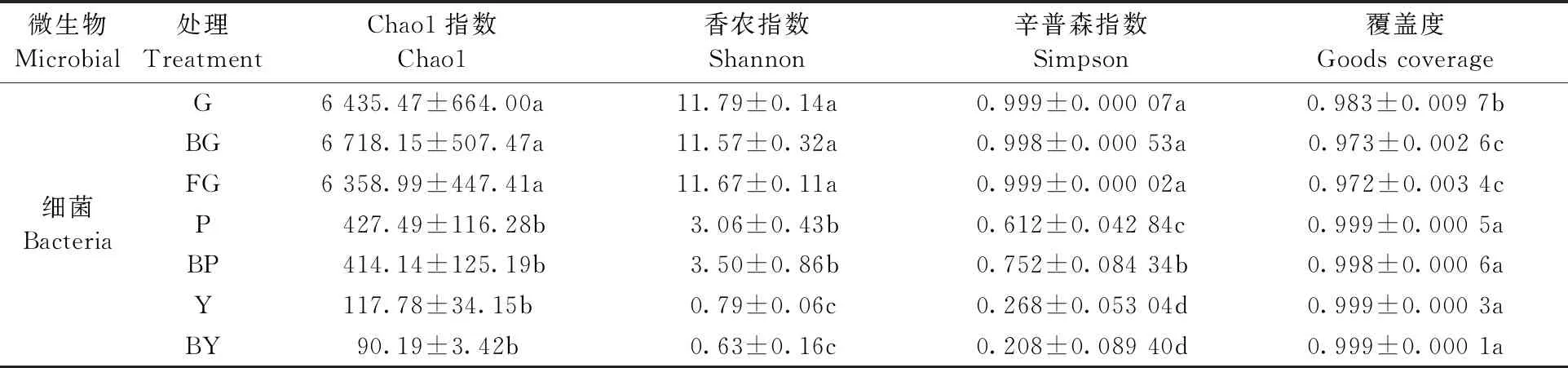
表1 不同器官區細菌Alpha多樣性指數①

圖7 不同器官區細菌PCoA分析
2.4 菌群代謝功能預測分析
使用PICRUSt2軟件對健康、感病山桐子中不同器官區的細菌微生物群落進行代謝功能預測。通過KEGG數據庫比對,結果共注釋到6條一級代謝通路,包括生物進程(cellular processes)、環境信息處理(environmental information processing)、遺傳信息處理(genetic information processing)、人類疾病(human diseases)、代謝(metabolism)和生物體系統(organismal systems); 一級代謝通路中代謝、遺傳信息處理為主要代謝通路,占全功能分類的80.68%~82.24%、11.26%~13.48%。在二級代謝通路中共注釋到34種代謝通路子功能,選取豐度占總豐度比值大于1.00%的子功能進行統計(表2)。山桐子不同器官區的微生物群落功能相似,但在子功能通路豐度上存在較大差異; 感病、健康山桐子的代謝功能,在土壤和葉片樣本中組內差異較小,樹皮樣本組內差異較大。細胞進程中的細胞運動(0.64%~2.72%)和環境信息處理中的膜運輸(1.58%~2.32%),在子功能通路豐度上呈P低于BP。遺傳信息處理中復制和修復(4.73%~4.87%)、折疊、分類和降解(3.08%~4.46%)、翻譯(2.52%~3.22%)為主要子功能,在子功能通路豐度上呈P高于BP。與代謝有關的主要子功能有11種,在各樣本中豐度均較高,其中5種子功能通路豐度為土壤>樹皮>葉片,包括碳水化合物代謝(12.01%~13.21%)、氨基酸代謝(11.63%~13.13%)、萜類化合物和聚酮類化合物的代謝(7.02%~9.72%)、外源生物降解與代謝(4.49%~7.74%)、脂質代謝(4.06%~7.25%); 輔助因子和維生素的代謝(11.4%~16.58%)、能量代謝(5.01%~9.51%)2種子功能通路豐度表現為葉片高于土壤和樹皮; 氨基酸代謝、能量代謝、聚糖生物合成與代謝、輔助因子和維生素的代謝、其他氨基酸代謝在子功能通路豐度上呈P高于BP; 脂質代謝、外源生物降解與代謝在子功能通路豐度上呈G高于BG,P低于BP。

表2 KEGG 數據庫主要代謝通路豐度
對已有代謝通路的豐度數據進行分析,使用metagenomeSeq方法對樣本組進行兩兩比較,分析相同器官區、不同樣本之間在功能上的差異(圖8),篩選條件為P<0.05。分析結果表明,在土壤中,BG組12,14和16元大環內酯的生物合成(Biosynthesis of 12-, 14-and 16-membered macrolides)、植物晝夜節律(Circadian rhythm-plant)、軸突導向(axon guidance)、倍半萜生物合成(sesquiterpenoid biosynthesis)、Ⅱ型聚酮主鏈的生物合成(biosynthesis of type II polyketide backbone)、Ⅱ型聚酮產品的生物合成(biosynthesis of type II polyketide products)、糖胺聚糖降解(Glycosaminoglycan degradation)和溶酶體(Lysosome)通路相關功能基因對照G組顯著增加,而萘降解(Naphthalene degradation)通路相關功能基因對照G組極顯著降低。在樹皮中,BP組的D-精氨酸和D-鳥氨酸代謝(D-Arginine and D-ornithine metabolism)和金黃色葡萄球菌感染(Staphylococcus aureus infection)通路相關功能基因對照P組極顯著增加,而類胡蘿卜素生物合成(Carotenoid biosynthesis)、弓形體病(Toxoplasmosis)和志賀菌病(Shigellosis)相關功能基因對照P組顯著降低。
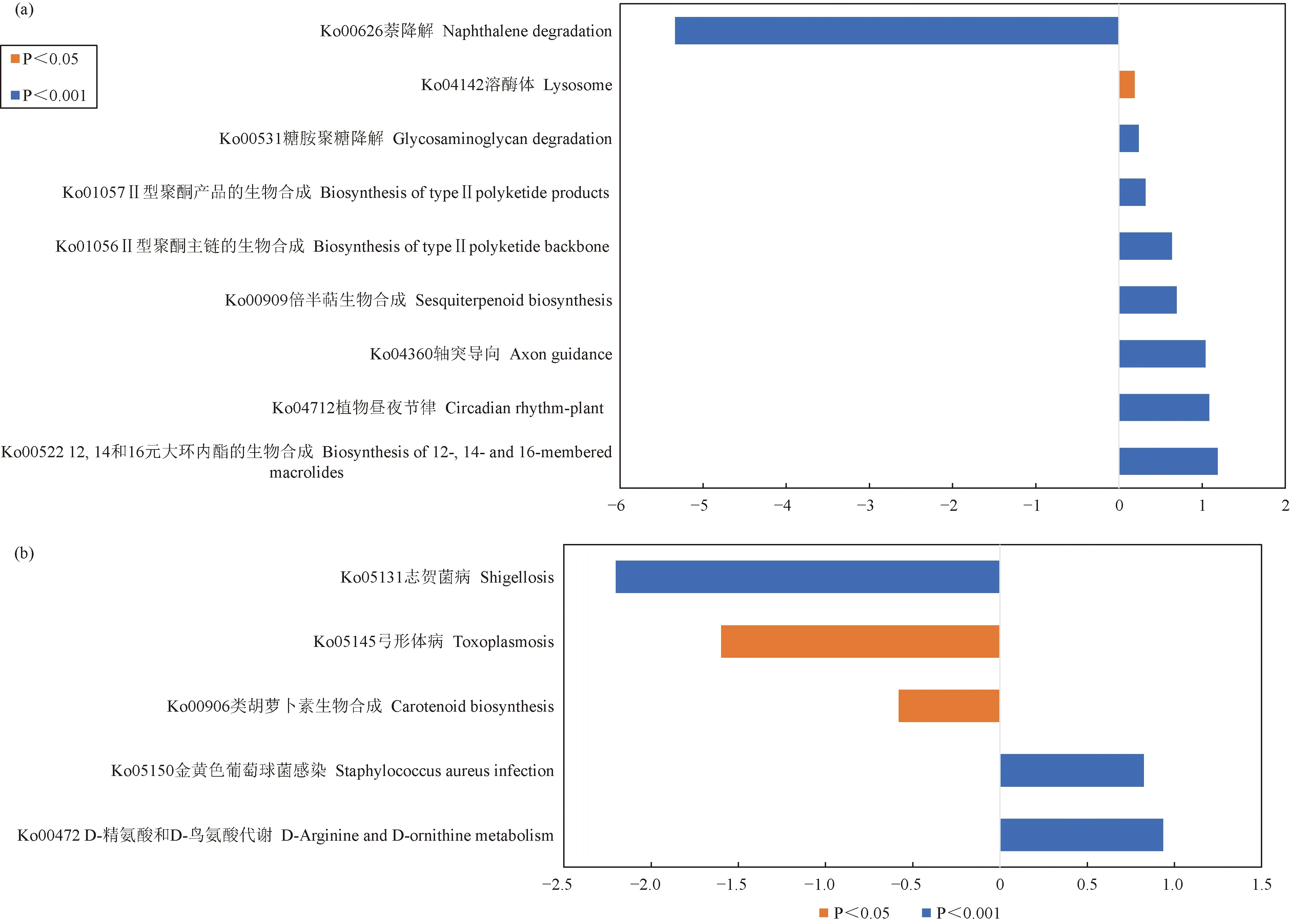
圖8 兩兩組間代謝通路存在顯著差異的功能分類
3 討論
3.1 健康和感病山桐子各器官區細菌群落結構特征
本研究采用Illumina miseq高通量測序技術分析健康和感病山桐子土壤、樹皮、葉片細菌群落結構和多樣性的差異。結果表明,山桐子不同器官區細菌群落結構存在明顯差異,OTU數量由高到低為土壤>樹皮>葉片,土壤相較于植物組織含有較多的OTU數量,這與其他林木研究結果相似(劉海燕等, 2019; 楊多等, 2022),一定程度上也反映了不同器官區環境的特性。進一步的物種分析注釋發現,山桐子不同器官區細菌群落的主要成分存在差異,在分類學上不同種類細菌的相對豐度發生變化。山桐子根際土壤細菌群落主要由變形菌門、放線菌門和酸桿菌門構成,這些優勢菌門在根際環境中富集,與其較強的環境適應性有關(王安寧等, 2019; Lietal., 2020)。其中變形菌門包括多種病原菌和固氮菌,因為其外膜主要由脂多糖組成可以保護其內部遺傳物質不受外部干擾,可以在大多數環境中生存并繁衍成為優勢菌(葛藝等, 2021; 翟婉璐等, 2017)。山桐子樹皮和葉片細菌群落主要由藍細菌門和變形菌門構成,這一結果與前人的研究發現相似(暴家兵等, 2020; 李慧娟等, 2021)。藍細菌門在山桐子葉際細菌中處于優勢地位,相對豐度范圍為84.58%~88.42%,產氧光細菌綱是其主要的細菌類群,有研究表明,藍細菌是地球上廣泛存在的一種自養型細菌,種類繁多,有些種類能夠進行產氧的光合作用,有些種類在暗環境下也能生長,營養要求低,對極端環境適應性很強(王文曉等, 2020)。山桐子樹皮和葉片中的微生物群落與土壤微生物群落差異較大,可能與不同器官區域的周邊環境有關,藍細菌能夠為山桐子樹皮、葉片區域提供更適宜生長的微環境,促進山桐子的生長。
3.2 健康和感病山桐子各器官區細菌群落差異菌屬
健康和感病山桐子各器官區細菌在屬水平有顯著差異。本研究中鞘氨醇單胞菌屬是健康山桐子各器官區共有的優勢菌屬,不動桿菌屬是健康山桐子樹皮、葉片的優勢菌屬,與抑病能力密切相關。鞘氨醇單胞菌屬具有植物促生特性(陳兆進等, 2021),鞘氨醇單胞菌在降解有毒物質、抵抗病原菌(秦立金等, 2018)、促進植物生長(葛藝等, 2021)等方面的功能。不動桿菌屬能產生生物表面活性劑和一些酶類物質,具有較高的促生、抗菌和降解有機污染物性能(程淺, 2020; 顧美英等, 2021)。從核桃根際土壤中分離到的2種不動桿菌屬菌株,經研究發現對核桃細菌性疫病具有明顯拮抗作用,可以抑制病原菌生長,進而提高植物的抗病性(涂昌等, 2020)。本研究從感病山桐子樹皮中篩選出的優勢菌屬為腸桿菌科的Brenneria和Hafnia-Obesumbacterium,在感病山桐子樹皮中的相對豐度均顯著高于健康山桐子樹皮。Brenneria是重要的林木病原細菌,該屬細菌侵染林木樹皮后會引起不規則的潰瘍病斑,對經濟林木生長有嚴重的威脅(Frutos, 2010)。國內外研究表明,Brenneria細菌會引起許多林木病害,例如Brennerianigrifluens是核桃樹皮潰瘍病的病原菌(Morettietal., 2007);Brenneriasalicis是柳樹水紋病的病原菌(Martineetal., 2009);Brenneriaquercina和B.corticissp. nov.是中國歐美楊潰瘍病的致病菌(楊莉, 2013; Lietal., 2019);Brenneriarubrifaciens可引起核桃深層樹皮潰瘍病(Amirsardarietal., 2017)等。Brenneria可能對山桐子樹皮區域病害的發生有協同或伴生作用,但其是否為山桐子感病的致病菌仍需進一步的研究。Hafnia-Obesumbacterium近年來已經被證明某些細菌具有較高的耐鹽性(王改萍等, 2021),而Qing等(2020)研究發現Hafnia-Obesumbacterium在感病林蛙腸道菌群中的相對豐度較高,可能與動物腹瀉相關。但是,對于Hafnia-Obesumbacterium在植物致病性方面的報道還較少,有待進一步探究。
3.3 健康和感病山桐子各器官區細菌群落多樣性
本研究對不同器官區山桐子細菌群落多樣性分析可知,山桐子各器官區細菌群落多樣性和豐富度由高到低均表現為土壤>樹皮>葉片,這與Wu等(2020)的研究結果類似,表明山桐子不同器官區細菌群落在多樣性和豐富度方面存在顯著差異。本研究中,感病山桐子樹皮細菌多樣性高于健康植株,樹皮細菌豐富度低于健康植株,這與林馬水等(2019)以健康和干腐病山核桃植株為研究對象,得出感病山核桃(Caryacathayensis)樹皮多樣性及豐富度指數均低于健康植株的結果不同,這可能是由于病菌在寄主體內存在時間的長短,對樹體生態平衡造成的影響不同; 同時,植株發病時間的長短也可能是造成感病山桐子樹皮組內細菌多樣性差異較大的主要原因。
3.4 健康和感病山桐子各器官區細菌功能特征
菌群代謝功能預測結果表明,山桐子中的細菌主要涉及代謝、遺傳信息處理等6條一級代謝通路和34種子功能,表現出功能上的豐富度。山桐子不同器官區樣本中所含的功能基因種類大致相同,說明山桐子相關細菌群的功能基因具有一定相似性。對山桐子不同器官區樣本細菌功能基因相對豐度進行分析,發現代謝通路在山桐子不同器官區中相對豐度均最大。在代謝通路中,代謝功能注釋相對豐度大多表現為土壤微生物組含有更多的碳水化合物代謝、氨基酸代謝、萜類化合物和聚酮類化合物的代謝、外源生物降解與代謝和脂質代謝,這可能與土壤益生菌的含量有關(馬欣等, 2021),其中碳水化合物代謝調控著生物體內碳水化合物的代謝形成、分解和互相轉化,有益于植物的氮、磷循環(馬欣等, 2021; 丁鈺珮等, 2021); 氨基酸代謝與碳代謝和氮代謝相關(南鎮武等, 2021); 萜類化合物代謝與植物抗病性和抗逆性有關(叢微等, 2021),間接說明土壤細菌代謝功能活躍有利于植物生長發育。而輔助因子和維生素的代謝、能量代謝在葉片微生物組中注釋豐度明顯高于土壤和樹皮,這2種代謝功能可能在維持植物葉際環境中發揮重要功能。通過代謝通路差異分析可知,感病山桐子根際土壤和樹皮中相關生物合成與氨基酸代謝的功能基因富集,說明感病后山桐子感病區域微生物群落的結構及其功能已做出了有益于生長發育的響應。而感病山桐子樹皮中金黃色葡萄球菌感染功能基因富集,造成代謝紊亂,降低山桐子抗病性,進而導致山桐子病害的發生。因PICRUSt功能預測分析只是初步預測相關細菌的功能,未來的研究將需要進一步驗證山桐子的致病菌,明晰引起山桐子病害的機制,更好地為山桐子病害防治及高質量經營提供理論支撐。
4 結論
健康和感病山桐子植株根際、樹皮及葉片細菌結構組成、多樣性和功能存在差異。山桐子土壤主要優勢菌門為變形菌門、放線菌門和酸桿菌門,樹皮和葉片主要優勢菌門為藍細菌門和變形菌門。健康山桐子各器官區中主要的優勢菌屬為鞘氨醇單胞菌屬和不動桿菌屬,感病后群落結構發生改變,健康樹皮與感病樹皮細菌群落結構差異顯著,感病山桐子樹皮的主要優勢菌屬為腸桿菌科的Brenneria和Hafnia-Obesumbacterium。功能預測表明山桐子各器官區細菌生態功能較為豐富,健康和感病山桐子的土壤及樹皮區域代謝功能有明顯差異。
