危害白蠟的斂片葉蜂屬(膜翅目: 葉蜂科)一新種*
2022-12-23 09:04:28牛耕耘王青華閆家河付懷軍劉瑞霞魏美才
林業科學 2022年8期
牛耕耘 王青華 閆家河 付懷軍 劉瑞霞 姜 敏 魏美才
(1.江西師范大學生命科學學院 南昌 330022; 2.中國林業科學研究院森林生態環境與自然保護研究所 北京 100091; 3.山東商河縣林業保護和發展中心 商河 251600; 4.北京市植物園 北京 100093; 5.山東省平邑縣林業發展中心 平邑 273300)
近年來,葉蜂類昆蟲對林木的危害性日趨嚴重,逐漸成為危害性僅次于鱗翅目的食葉害蟲。不僅傳統上常見的知名葉蜂害蟲類群中出現了一些新的危害種類,如松葉蜂科的青海新松葉蜂(NeodiprionqinghaiicusWei & Wu, 2021)(武星煜等, 2021)、扁蜂科的黃腹阿扁蜂(AcantholydaxanthogasterWu & Xin, 2014)(武星煜等, 2014)和煙翅腮扁蜂(CephalciainfumataZhang & Wei, 2018)(張寧等, 2018)分別在祁連山東段和賀蘭山嚴重危害青海云杉(Piceacrassifolia),葉蜂科突瓣葉蜂亞科的中華厚爪葉蜂(StauronematussinicusLiu, Li & Wei, 2018)在中國東部平原地帶廣泛危害楊樹(Populusspp.)(劉萌萌等, 2018)、白榆突瓣葉蜂(NematuspumilaLiu, Li & Wei, 2019)在華東和華北一帶嚴重危害榆樹(Ulmuspumila)(劉萌萌等, 2019)、錘角葉蜂科的萌萌丑錘角葉蜂(ZaraeamengmengYan, Li & Wei, 2020)在華東地區危害稠李(Prunuspadus)(晏毓晨等, 2020)、中華唇錘角葉蜂(LabriocimbexsinicusWei & Yan, 2019)在江南一帶危害櫻桃(Prunuspseudocerasus)(Yanetal., 2019),而且一些原來十分稀見的葉蜂類群中也陸續出現了在局部地區危害嚴重的林木害蟲,如: 多種巨棒蜂(Megaxyelaspp.)在中國多地局部危害胡桃科植物(Blanketal., 2017; 牛耕耘等, 2021),多種枝膜葉蜂(Cladiuchaspp. )(牛耕耘等, 2021)和巨基葉蜂(Megabelesesspp.)(李澤建等, 2021)在中國南方嚴重危害不同木蘭科植物,多種櫻實葉蜂(Analcellicampaspp.)在多地嚴重危害櫻桃屬植物(Niuetal., 2019; 牛耕耘等, 2021)。
白蠟(Fraxinusspp.)是中國東部地區常見的木犀科梣屬植物。近年來除了發現白蠟外齒莖蜂(StenocephusfraxiniWei, 2015)在國內廣泛危害白蠟外(Weietal., 2015; 閆家河等, 2018),過去2年中在山東、河北、天津和北京一帶多個地方,幾乎同時發現一種新的害蟲取食多種白蠟樹葉,嚴重危害其生長。山東泰安市徂徠山林場、平邑縣的蒙山山區多個林場白蠟葉蜂種群極大。近5年來,幾乎每年都有很多白蠟被吃成光桿或嚴重吃殘吃花,危害嚴重。蒙山的下山路上,成蟲隨處可見,個別地段的地面落滿了成蟲。該種害蟲經比較形態研究和線粒體基因組測序分析,確認是以前記載、偶爾危害桂花樹(Osmanthusfragrans)的葉蜂科斂片葉蜂屬(Tomostethus)一個新的物種。
1 材料與方法
1.1 材料
新種模式標本分別采集于山東蒙山、北京門頭溝和北京市植物園,保存于中國南昌亞洲葉蜂博物館(ASCN)。
1.2 術語和縮寫
構造描述術語采用Ross(1945)和Niu等(2010)。
1.3 圖片拍攝和處理
成蟲整體和細節照片采用NikonD700數碼相機拍攝多幅原始圖像,然后使用Helicon Focus(HeliconSoft?)軟件進行圖片合成,獲得多焦點合成的成蟲圖片,最后在Photoshop? CS6軟件上進行調光、去雜污點等簡單處理,獲得最終圖片。雄蟲和雌蟲外生殖器圖片分別使用Moticam? 5000和 motic? BA400獲得原始圖像,然后在Photoshop? CS6軟件上進行調光、去雜污點等簡單處理,獲得最終圖片。卵粒照片采用奧林帕斯CX31顯微鏡拍攝,聚集為害的幼蟲采用NIKON 5300相機拍攝,寄主植物和成蟲生態照片使用Canon G9相機拍攝。
1.4 線粒體基因組測序和分析
選取2個樣本進行了2代基因組測序。成蟲(雄蟲: CSCS-Hym-MC0323)采集自山東蒙山,幼蟲(CSCS-Hym-MC0380)采集自北京平谷,測序殘體均保存于中國南昌亞洲葉蜂博物館(ASCN)。
成蟲樣品取胸部肌肉組織,幼蟲取體壁。采用Qiagen試劑盒(DNeasy Blood & Tissue Kit)提取基因組DNA。使用Illumina Hiseq 4000 platform進行2代測序。原始數據經質檢、修剪后獲得clean reads。分別利用GetOrganelle(Jinetal, 2020)進行無參組裝,利用Geneious(https:∥www.geneious.com)軟件使用近緣種作為參考序列進行組裝,對無參組裝結果進行檢驗。利用MITOS Web Server(http:∥mitos.bioinf.uni-leipzig.de/index.py)對tRNA進行注釋,根據與近緣種的比較研究,確定蛋白質編碼基因(PCG)和rRNA的起止位置。
1.5 cox1序列和基因組比較分析研究
采用MEGA7軟件(Kumaretal., 2015)對斂片葉蜂屬(Eutomostethus)內3個種及近緣屬真片葉蜂屬的條刻真片葉蜂(E.vegetus)的cox1序列進行基于密碼子的多序列比對。比對后有效序列長度1542 bp。使用K2P(Kimura-2-parameter)模型(Kimura, 1980)計算序列遺傳距離。另對本種內來自2個不同種群的線粒體基因組的全部13個蛋白質編碼基因統計P距離。
1.6 基于線粒體基因組的種級系統樹構建
選擇條刻真片葉蜂作為外群,對斂片葉蜂屬(Tomostethus)內3個種的7條cox1序列,使用TranslatorX(Abascaletal., 2010)進行比對,并用IQtree軟件(Nguyenetal., 2015)構建最大似然樹,參數設置為默認。
2 結果與分析
2.1 白蠟斂片葉蜂 Tomostethus fraxini Niu & Wei, sp. nov.(圖1—3)
雌蟲: 體長8.5mm(圖1); 體和足黑色,腹部2—8背板后緣狹邊和2—5腹板后緣狹邊灰白色; 體毛銀色,鋸鞘毛黑褐色; 翅深煙灰色。翅痣和翅脈黑褐色。
唇基具明顯的細小稀疏刻點; 額區具極微弱刻紋,雜以細小稀疏刻點(圖2a); 上眶和單眼后區大部光滑,具極稀疏的細小刻點(圖2b); 后眶刻點稍細密; 觸角基部2節較光滑,鞭節表面刻紋密集(圖2k); 前胸背板刻點較明顯,中胸背板前葉和側葉刻紋微弱,刻點不明顯,小盾片表面光滑,刻點細小、清晰,附片高度光滑; 中胸前側片上半部稀疏具毛小刻點,刻點間隙光滑,下半部光滑,具很寬的無毛裸區(圖2h); 后側片大部光滑; 腹部背板具明顯的微細刻紋,腹板刻紋不明顯。
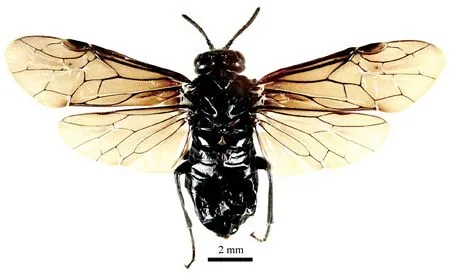
圖1 白蠟斂片葉蜂成蟲背面觀,新種,正模(雌)
頭部前面觀寬大于高; 復眼大,內緣向下明顯收斂,間距稍寬于眼高; 顎眼距線狀(圖2a); 觸角窩上突明顯隆起; 中窩十分寬深,側窩明顯較小; 額區五角形,額脊鈍,前緣脊明顯低于側脊; 單眼中溝寬點狀,前臂深長; 單眼后溝深,中部稍前突,中單眼前凹圓形,明顯小于單眼; 單復眼距∶側單眼距∶ 單眼后頭距=14∶13∶11; 單眼后區顯著隆起,稍高于單眼頂面,寬長比約等于2,具較淺但明顯的中縱溝; 側溝十分寬且深直,互相平行; 背面觀后頭兩側弧形收窄,短于復眼長的1/3(圖2b); 后眶圓鈍,后眶溝深長(圖2c),無后頰脊,顎眼距下側具短斜脊; 觸角粗短,等長于頭寬,第1節主體明顯傾斜,內側弧形彎曲,第2節長稍大于寬,第3節微長于第4、5節之和(10∶9),第7、8節長約等于寬(圖2k)。小盾片附片寬大; 胸腹側片寬平(圖2h); 后胸淡膜區寬,間距等長于淡膜區長徑1/2; 前足脛節內距端部分叉(圖2i); 后足脛節與跗節長度比為9∶7; 基跗節稍長于其后2節之和; 爪小型,無內齒(圖2j)。前翅2r脈位于2Rs室端部1/5處,2 m-cu位于2Rs室內側1/3處,1 m脈與1 m-cu脈向翅痣明顯收斂,2Rs室等長于1R1+1Rs室之和,cu-a脈中位,2A+3A脈長且彎曲,末端靠近但不接觸1A脈; 后翅M室封閉,臀室柄微短于cu-a脈(圖1)。腹部第1背板中部膜區小三角形; 產卵器明顯短于后足跗節,側面觀鞘端等長于鋸鞘基,腹側和端緣弧形彎曲(圖2e),鋸鞘刷狹窄,兩側弧形彎曲(圖2f); 背面觀鋸鞘端端部圓鈍,鋸鞘毛伸向后側(圖2 d); 鋸腹片窄長,亞端部微弱膨大,18刃,鋸刃微弱突出,中部鋸刃具8個內側亞基齒、3個外側亞基齒,節縫刺毛帶密集,互相連接(圖2m)。
雄蟲(副模): 體長7.5mm; 體色和構造類似雌蟲,但復眼后頭部更短,下生殖板長約等于寬,端部圓鈍; 抱器長大于寬,端緣中部明顯突出,內緣下部1/3具明顯缺口,副陽莖窄高(圖2l); 陽莖瓣頭葉窄長,背腹緣近似平行,端部鈍截,端位橫刺突三角形,十分顯著(圖2g)。
量度和變異: 雌蟲體長8~8.5mm,雄蟲體長5~7.5mm; 雌蟲鋸腹片中部鋸刃具8~9個內側亞基齒、3~4個外側亞基齒; 體色無變化; 單眼后區中縱溝的深度稍有變化。

圖2 白蠟斂片葉蜂,新種,副模
正模: ♀,北京植物園,40°04′17″N,116°12′4.82″E,海拔78 m, 2020-04-01,付懷軍,周達康; 副模: 20♀10,采集信息同正模; 5♀1,山東蒙山九龍潭(海拔580mm,117°50′40″E,35°32′26″N)至山頂(海拔1030 m,117°51′9″E,35°33′42″N); 1♀,北京門頭溝, 2012-05-01,李濤; 3♀9,天津薊州區國營林場黃花山林區, 2021-04-08,張慧超。
詞源: 本種以其寄主植物命名。
分布: 河北(承德)、北京(門頭溝、海淀、平谷)、天津(薊州)、山東(蒙山、徂徠山)。
寄主: 白蠟樹(F.chinensis)和花曲柳(F.chinensissubsp.rhynchophylla)等。
鑒別特征: 本種單眼后區強烈隆起,寬長比約等于2; 觸角第3節微長于第4、5節之和; 體和足全部黑色,體毛銀色; 中胸前側片下部具寬闊裸帶; 腹部背板具微細刻紋,鋸鞘毛黑褐色,伸向后方等,與本屬其余種類不同(參見下文分種檢索表)。

圖3 白蠟斂片葉蜂卵、幼蟲和成蟲
生物學概要: 1年1代。以老熟幼蟲在表土層5cm左右綴土粒作土繭,以預蛹越夏越冬。翌年4月上旬成蟲羽化,羽化期隨海拔差異有一定變化。成蟲在白蠟嫩芽附近飛行和交配(圖3g)。卵產于白蠟葉片邊緣的葉肉中,排列緊密,葉緣明顯鼓脹(圖3a)。幼蟲孵化前,黑色的頭部清晰可見(圖3e)。幼蟲剛蛻皮時體表光亮,頭部黑色,胸部和腹部末端黃色,其余部分乳白色,瘤突短小,近透明。老熟幼蟲體表刺突顯著,黑色,每個體節具2排瘤突(圖3c、3h)。幼蟲食葉,發生數量較大時有聚集現象(圖3f)。在山東的蒙山,4月中旬至5月底或6月上旬為幼蟲危害期。本種詳細的生物、生態學特性將另文總結報道。
2.2 斂片葉蜂屬世界種類及其檢索表
斂片葉蜂屬的主要鑒別特征是: 前足脛節內距端部分叉,爪無內齒; 鋸鞘短于中足脛節,端部具短耳形側突和側毛刷; 觸角短,約等于或稍長于頭部寬,顯著短于頭胸部之和,第3節長1.8~2.2倍于第4節; 前翅1 m脈與1 m-cu脈向翅痣顯著收斂,2A+3A脈長,約0.8倍于臀室基柄長,端部靠近1A脈; 后翅臀室柄很短,長約0.5~1.0倍于cu-a脈; 陽莖瓣頭葉窄長,端部具小而尖銳的刺突。這些特征可以很容易地將本屬與藺葉蜂亞科各屬鑒別。
斂片葉蜂屬是一個小屬。Taeger等(2010)在世界葉蜂目錄中的斂片葉蜂屬下列出了8種,其中TomostethusvelesKonow, 1900(分布于東西伯利亞)和T.kirbyiKonow, 1904(分布于南美)2種,根據原始描述(前翅1 m脈與1 m-cu脈互相平行)可以確定他們不是本屬成員,而是真片葉蜂屬的成員。其余6種中,1種分布于北美,5種分布于歐亞大陸。Taeger等(2018)在斂片葉蜂 屬下還列出了T.testaceusNiezabitowski, 1899。但根據原始描述(前翅1 m脈與1 m-cu脈互相平行),該種應是真片葉蜂屬的成員。
新種白蠟斂片葉蜂和本屬其他已知種類可以使用以下檢索表進行鑒別。
2.3 斂片葉蜂屬分種檢索表
1 腹部黃褐色,第1背板和鋸鞘黑色。歐洲…
…黃腹斂片葉蜂T.melanopygus(Costa)
腹部黑色,最多背板后緣狹邊白色
2
2 雌蟲前胸背板大部和翅基片、雄蟲翅基片部分淡色或足大部分黃褐色
3
胸部和足完全黑色
4
3 雌蟲前胸背板大部和翅基片、雄蟲翅基片部分淡色; 足黑色,股節端部、脛節基部黃白色,跗節褐色; 觸角第3節不長于4、5節之和。北美
黃肩斂片葉蜂T.multicinctus(Rohwer)
兩性胸部全部黑色; 足黃褐色,基節、轉節和股節的基部黑色,跗節暗褐色; 觸角第3節長于第4、5節之和。西亞
黃足斂片葉蜂T.orientalisZirngiebl
4 單眼后區寬長比約等于2,側溝長約1.5~2倍于單眼直徑
5
單眼后區寬長比約等于3,側溝寬短點狀,約等長于單眼直徑; 后眶溝深; 背面觀后頭兩側不膨大; 腹部背板具明顯細刻紋。中國、日本、韓國
淡毛斂片葉蜂T.sp.
5 體毛暗褐色或黑褐色; 腹部背板光滑; 背面觀雌蟲后頭兩側明顯膨大; 后眶溝不明顯發育,極淺弱; 胸部細毛長于單眼直徑,胸部側板中下部無裸帶
6
頭胸部體毛銀色; 腹部背板具微細刻紋; 背面觀雌蟲后頭兩側明顯收縮; 后眶溝明顯發育,深溝狀; 胸部細毛短于單眼直徑,中胸前側片中下部具寬闊的光裸無毛帶
………白蠟斂片葉蜂T.fraxinisp. nov.
6 觸角第3節長于第4、5節之和; 單眼后區寬長比等于2; 雌蟲體長6mm; 寄主為Juglans。韓國
胡桃斂片葉蜂T.juglansTakeuchi
觸角第3節稍短于第4、5節之和; 單眼后區寬長比小于2; 雌蟲體長約8mm; 寄主為Fraxinus
7
7 前翅透明; 頭胸部細毛暗褐色。西亞、南歐、北非
淡翅斂片葉蜂T.claripennisEnslin
前翅煙褐色,端部稍淡; 頭胸部細毛黑褐色。歐洲、日本
黑毛斂片葉蜂T.nigritus(Fabricius)
2.3 基于線粒體基因組的白蠟斂片葉蜂山東和北京種群差異
本種目前已經發現在北京、天津、河北、山東4省(市)有分布。對山東蒙山種群和北京海淀種群的線粒體基因組蛋白質編碼基因進行無校正遺傳距離統計分析(表1)。結果表明,2個種群十分近似,線粒體編碼基因蛋白質編碼基因遺傳距離均值為0.004 6,其中遺傳距離最大的是nad2(0.008 6),atp8和nad4l遺傳距離為0。據此可以確定兩地種群屬于同一物種。

表1 白蠟斂片葉蜂山東蒙山種群和北京海淀種群的線粒體PCG遺傳距離
2.4 基于線粒體基因組的斂片葉蜂屬種間關系和遺傳距離
中國北京和山東種群的白蠟斂片葉蜂線粒體全基因組是本屬報道的第1、2條線粒體全基因組。除此以外,本屬世界上還有另外2個確定種報道了5條cox1序列。對這3種7條序列,進行了cox1基因的K2P遺傳距離統計分析。結果表明本屬已知3種的種間遺傳距離為0.070 3~0.141 7,其中北美分布的T.multicinctus與歐亞大陸種類之間的遺傳距離最大,達到0.118 1~0.141 7。本屬3種與近緣屬的代表種Eutomostethusvegetus的遺傳距離為0.119 9~0.166 6(表2)。
選擇斂片葉蜂屬的近緣屬種E.vegetus作為外群,基于cox1基因序列信息,應用最大似然法方法推斷了斂片葉蜂屬3種之間的系統發育關系(圖4),結果表明亞洲的白蠟斂片葉蜂與歐洲的黑毛斂片葉蜂是姊妹種,北美的T.multicinctus與歐亞大陸分布的白蠟斂片葉蜂和黑毛斂片葉蜂關系較遠。
需要注意的是,coxI基因序列的K2P距離和系統發育關系樹均顯示歐洲的黑毛斂片葉蜂的南歐和北歐種群之間已經存在明顯的分化,K2P距離超過了3%,而各自的種群內差異僅分別為0和0.22%,黑毛斂片葉蜂的南歐和北歐種群,有可能已經分化為獨立的種系。但因為參與分析的樣本很少,并且僅有coxI序列數據可以使用,這個發現有待后續基于更多區域、更多樣本的進一步研究進行判斷。

表2 幾種斂片葉蜂屬cox1基因序列K2P距離(下三角)和標準差(上三角)

圖4 基于cox1基因序列的斂片葉蜂屬3種之間的系統發育關系
