馬尾松非結構性碳庫大小及分配的緯向變化*
2022-12-21 11:04:00倪妍妍簡尊吉曾立雄阮宏華肖文發李邁和
林業科學 2022年8期
關鍵詞:生長
倪妍妍 簡尊吉 徐 瑾 曾立雄, 阮宏華 雷 蕾 肖文發, 李邁和
(1.中國林業科學研究院森林生態環境與自然保護研究所 國家林業和草原局森林生態環境重點實驗室 北京 100091; 2.南京林業大學南方現代林業協同創新中心 南京 210037; 3.瑞士聯邦森林、雪與景觀研究所 蘇黎世 8903; 4.東北師范大學地理科學學院 長白山地理過程與生態安全教育部重點實驗室 長春 130024)
非結構性碳水化合物(non-structural carbohydrates, NSC)是植物生存和生長的物質基礎(Chapinetal., 1990; Martínez-Vilaltaetal., 2016)。NSC庫由可溶性糖(蔗糖為主)和淀粉組成。樹木光合同化形成的碳優先供給呼吸和生長后,將剩余的碳貯存形成穩定的NSC庫(Barbarouxetal., 2002; Goughetal., 2009; Rochaetal., 2009)。NSC的動態變化可作為評估碳“源-匯”關系的指標(K?rner, 2003),NSC庫大小與物候節律、樹木個體大小及所處環境等密切相關(倪妍妍等, 2017; Schoonmakeretal., 2021)。此外,Richardson等(2013)通過美國通量網Howland定位站長期觀測發現,NSC的長期逐漸增加量與木質生物量的增加比例基本一致,表明NSC庫不僅影響樹木個體生長,還在生態系統碳循環中發揮重要作用。然而,迄今關于樹木體內NSC的儲存與分配特征的研究相對較少。
樹木器官NSC庫的大小可表征林分地上和地下生產力的分配情況(Lietal., 2018)。樹木不同器官的NSC含量差異顯著(Boldingh, 2000; Lietal., 2001; Smithetal., 2017),因此,利用單個器官NSC含量無法準確估計整個植株的NSC存儲量。此外,樹木不同器官對植株生長和代謝過程的作用與貢獻不同(Lietal., 2001; 2002; Hochetal., 2003; Palacioetal., 2014)。例如,常綠樹種葉片既是植株碳水化合物生產的源(Kozlowski, 1992; Hartmannetal., 2016),也是儲存碳水化合物的庫(Lietal., 2001; 2002); 樹干除了作為碳水化合物的高消耗器官外,同時承擔了輸導功能和存儲功能(Barbarouxetal., 2003; Hochetal., 2003); 根系含有較大比例的射線和軸向薄壁組織細胞,具備相對較高的存儲能力,且這些儲存主要被用于次年新枝生長(Lensetal., 2000; Zhuetal., 2012)。但Furze等(2019)研究發現,溫帶5個樹種的枝條全年儲存了足夠多的淀粉,導致根系并不是主要儲存器官,而且根系的儲存并沒有用于支持春季生長。因此,探究不同器官NSC庫的動態變化,有利于深入了解樹木的生長動態及其對環境變化的響應。
NSC的儲存是對不同時空尺度下碳水化合物供應與需求非同步性的一種緩沖,具有明顯的時空效應(Hochetal., 2012)。樹木NSC含量高低和儲存大小除與物種生物學特性和自身生長機制密切相關外(Congetal., 2018),也與生長環境密切相關(Wangetal., 2018)。樹木NSC含量隨緯度、海拔的變化特征有諸多研究(Martínez-Vilaltaetal., 2016; Liuetal., 2018; Liuetal., 2019; 杜建會等, 2020)。如Martínez-Vilalta等(2016)發現從寒溫帶針葉林到熱帶雨林,NSC及其組分在樹干和根系中的含量隨緯度無顯著變化,而在葉片中的含量隨緯度增加而增加。目前,在大空間尺度上探究某單一樹種不同器官NSC儲存量變化的研究較少(Liuetal., 2019)。
馬尾松(Pinusmassoniana)是我國重要的鄉土用材樹種,在亞熱帶濕潤區域廣泛栽植,是探究跨地域尺度分布物種生存策略的理想材料。因此,本研究以馬尾松成熟林為研究對象,在其分布區內沿緯向梯度在不同緯向梯度選擇9個典型人工林,研究馬尾松NSC存儲及分配的緯向變化規律,并探討生長環境對其影響作用,為準確理解和模擬氣候變化背景下樹木生長和碳存儲提供依據。
1 材料與方法
1.1 研究區概況
根據馬尾松在我國的分布和生長情況,在不同緯度(23.0°—33.5°N)選擇9個馬尾松人工林純林,其中馬尾松蓄積量不低于林分總蓄積量的65%。采樣點從南到北依次為廣東肇慶、廣西賀州、廣西桂林、湖南會同、湖南永順、湖北宣恩、重慶忠縣、四川萬源和陜西漢中。樣地年均溫度和年均降水量從南到北遞減,地帶性土壤類型主要為紅壤、黃壤和黃棕壤。依據國家標準《森林資源連續清查技術規程GB/T38590-2020》(國家林業和草原局, 2020)中馬尾松人工林的齡組劃分,優先選擇林齡30~35年段的成熟林,其次選擇25~30年段的近熟林。各地氣候特征和林分結構存在差異,具體信息詳見表1。

表1 馬尾松人工林樣地的地理位置、氣候和林分特征
1.2 野外調查及樣品處理
2018年10—12月,從北到南依次采集樣品。每個樣點選擇3~4塊樣地(20 m × 30 m), 同一樣點每2塊樣地間的間距不小于1 km。在每塊樣地內選擇10~15個點,用土鉆收集表層土(0~20cm),混裝成1個樣品,用于測定土壤養分。每木檢尺記錄樣地內所有胸徑大于4cm的馬尾松的胸徑和樹高,確定5~7株標準木用于收集根、莖、枝、葉等器官樣品。對每一株標準木,在樹高2/3~3/4處選擇2個陽生枝收集針葉和枝條; 利用生長錐在樹干1.3 m胸徑處鉆取樹干樣品用來測定樹木年齡并測定NSC含量; 利用軍工鏟收集細根。植物器官樣品按照樣地進行混裝。在進入實驗室預處理前,樣品放置在保鮮箱。
在實驗室內,將枝、莖、根去皮。所有植株樣品用水清洗干凈后用蒸餾水沖洗2次,于105℃下殺青15min,在65℃下烘干至恒質量,粉碎、過篩(0.15mm孔徑)后儲存在磨口瓶中待用。土壤樣品置于陰涼處風干,過篩(0.07mm孔徑)后儲存待用。
1.3 NSC及土壤養分含量測定
NSC含量為同一樣品中可溶性糖和淀粉之和。采用蒽酮比色法測定樣品可溶性糖和淀粉含量(Lietal., 2018)。
土壤樣品(1.2~1.5 mg)用專用錫箔紙包裹后置于元素分析儀(Euro EA, Hekatech Gabh, 德國)上測定全碳、全氮元素含量。樣品先用酸消解(HNO3和HClO4)后用離子發射光譜儀(IRIS Intrepid II XSP, Thermo, 美國)測定全磷和全鉀含量。
1.4 植物器官生物量估算和NSC及其組分儲存量計算
用異速生長方程模型估算樣地馬尾松各器官的生物量:
M=aDbHc。
式中:M為各器官生物量;D為胸徑;H為樹高;a,b,c為參數,具體數值參考林業行業標準《立木生物量模型及碳計量參數——馬尾松LY-T 2263-2014》(國家林業和草原局, 2014)。根據不同區域的異速生長方程模型求算樣地馬尾松的各器官生物量。
與分布區北界相比,馬尾松在南界生長快、生長季長而輪伐周期短(表1)。為了標準化地表達NSC儲存,采用樹木各器官生物量及整株總生物量的年平均值。馬尾松各器官的NSC庫是利用器官NSC含量乘以器官年均生物量計算獲得。樹木NSC庫為各器官庫總和。
1.5 氣象因子
從中國氣象數據網(http:∥www.data.cma.an/)下載馬尾松分布區內氣象站點逐年數據。本研究選擇1979—2018年間氣候平均值數據。用Arcgis v10.0 進行克里金空間插值, 生成分辨率1 km的氣象柵格圖層,最后根據地理坐標提取各樣地的氣象數據。氣象變量包括年平均溫度、年均降雨總量、年均最冷月最低溫、年均最熱月最高溫和帕默爾干旱強度指數。
1.6 數據分析
數據分析和作圖均由R4.0.5軟件(http:∥www.r-project.org/)完成。所有數據通過正態分布(Shapiro-Wilk檢驗)和方差齊性檢驗(Bartlett檢驗),滿足數據進一步分析條件。利用線性混合效應模型,將樣地和樹齡作為隨機因子,分析器官、緯度及其交互作用對馬尾松生物量和NSC及其組分的含量、庫大小以及庫分配比例的影響。利用簡單線性回歸模型分析植株整體和各器官的NSC及其組分的濃度含量、庫大小以及庫分配比例隨緯度變化的線性趨勢。通過皮爾遜相關分析,分析馬尾松整株NSC及其組分庫大小與環境因子的關系,再按氣象因子和土壤因子將顯著的因子分類進行冗余分析,獲得各因子的解釋率。
2 結果與分析
2.1 器官生物量與非結構性碳水化合物含量的緯度變化
如表2所示,器官、緯度以及兩者間的交互作用對馬尾松年均生物量、可溶性糖和NSC含量影響顯著(P<0.01),緯度對淀粉含量作用顯著(P<0.001)。馬尾松各器官(根、干、枝和葉)的年均生物量均隨緯度升高而降低(圖1),干中NSC、可溶性糖和淀粉含量也隨緯度增加而降低,但在其他器官中無顯著趨勢(圖1)。
2.2 非結構性碳水化合物存儲的緯度變化
馬尾松整株NSC庫、可溶性糖庫和淀粉庫的大小均受緯度影響顯著(P<0.001, 表3),且均隨緯度增加而降低(圖2)。器官、緯度及其兩者間的交互作用對NSC庫、可溶性糖庫和淀粉庫的大小影響顯著(P<0.001, 表3)。各緯度的枝、干和根的淀粉庫顯著高于可溶性糖庫(表4),且枝和根中的可溶性糖庫、所有器官中的淀粉庫和枝、干、根中的NSC庫均隨緯度增加而降低(圖3)。

表2 器官、緯度及其交互作用對馬尾松的生物量和NSC含量的影響(F值)①
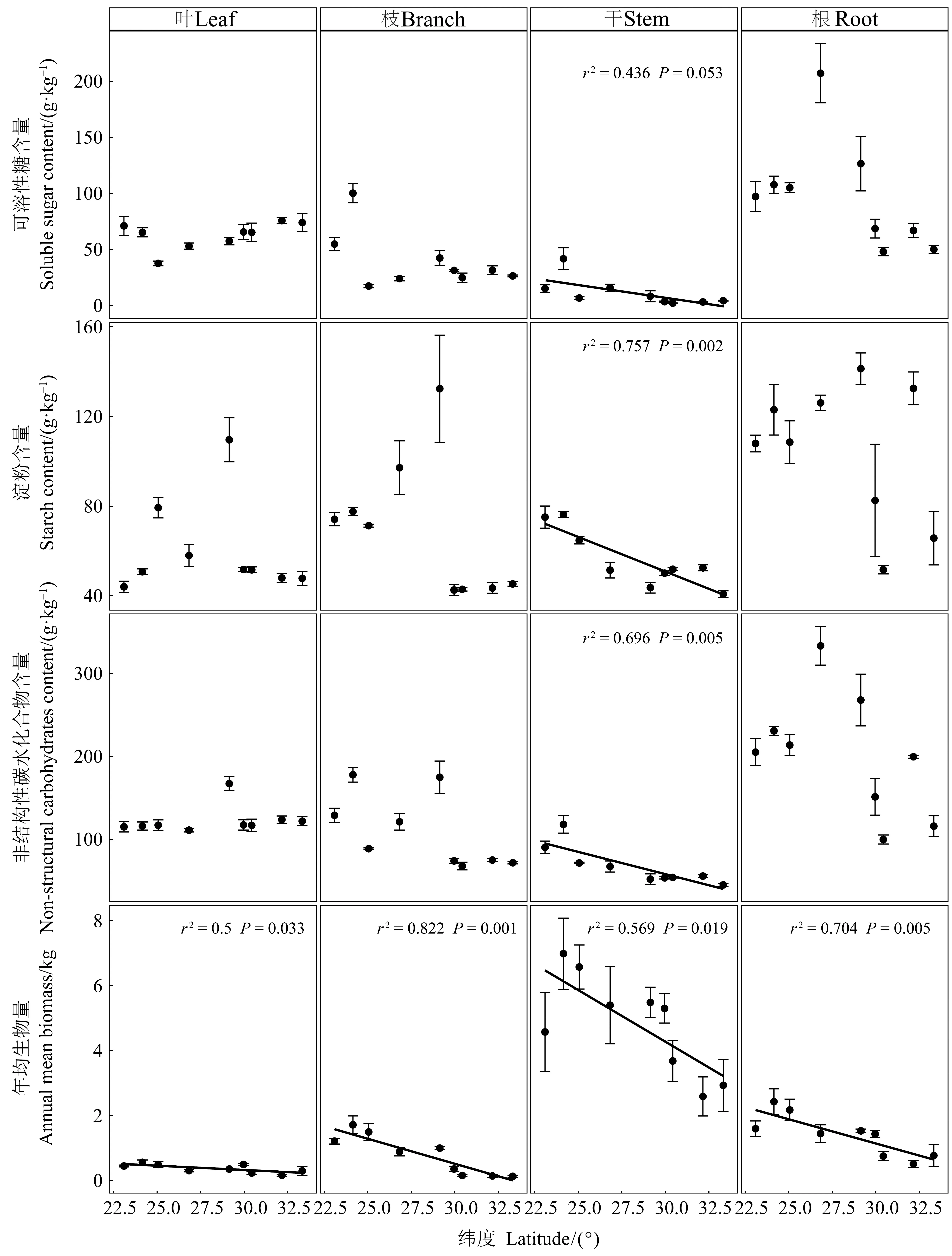
圖1 馬尾松不同器官年均生物量和非結構性碳水化合物含量的緯向變化

表3 器官、緯度及其交互作用對馬尾松的NSC庫大小的影響(F值)
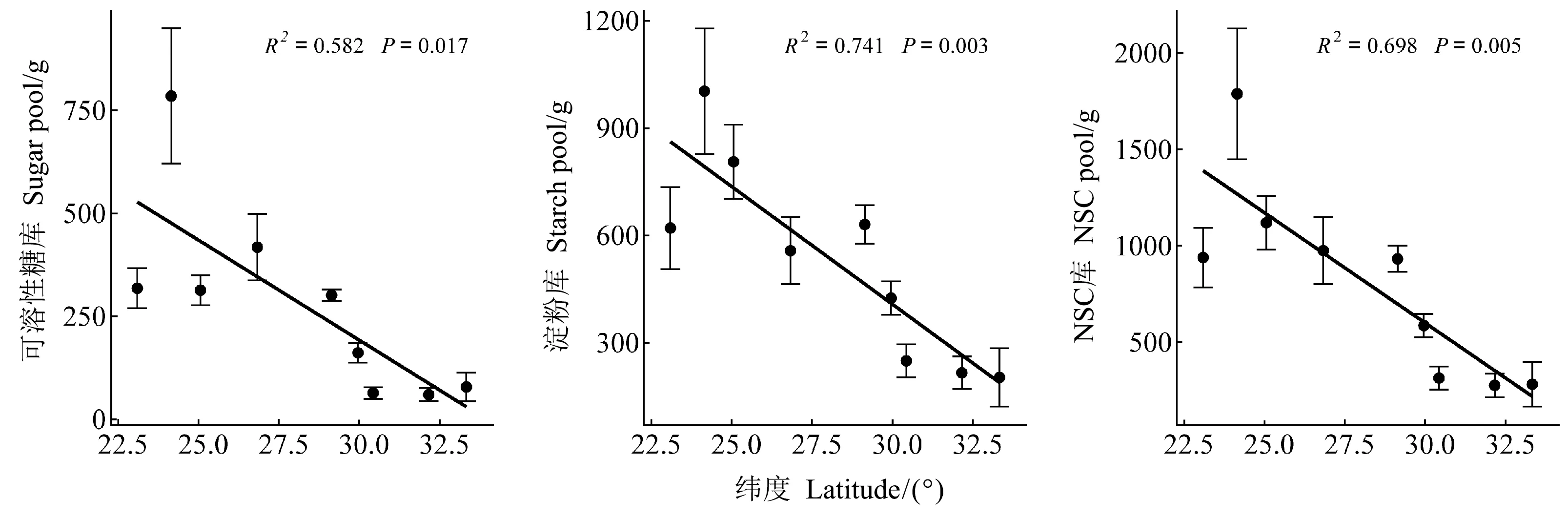
圖2 馬尾松整株NSC及組分庫的緯向變化

表4 馬尾松不同器官NSC及組分庫的大小(g)和分配比例(%)

圖3 馬尾松不同器官NSC及其組分庫大小的緯向變化
2.3 非結構性碳水化合物庫在各器官中的分配比例
可溶性糖庫和淀粉庫平均大小分別占全樹NSC庫的31.1%和68.9%,表明馬尾松NSC庫組成整體以淀粉為主。馬尾松整株的可溶性糖庫占NSC庫的比例隨緯度增加而降低,淀粉庫所占NSC的比例隨緯度增加而增加(圖4)。由表4和圖5可知,NSC庫、可溶性糖庫和淀粉庫在根、干、枝和葉中的分配比例差異均顯著,但均主要儲存在干和根中。隨緯度增加,馬尾松整株的NSC庫、可溶性糖庫和淀粉庫在枝中的分配比例降低,可溶性糖庫和NSC庫在葉中的分配比例增加(圖5)。
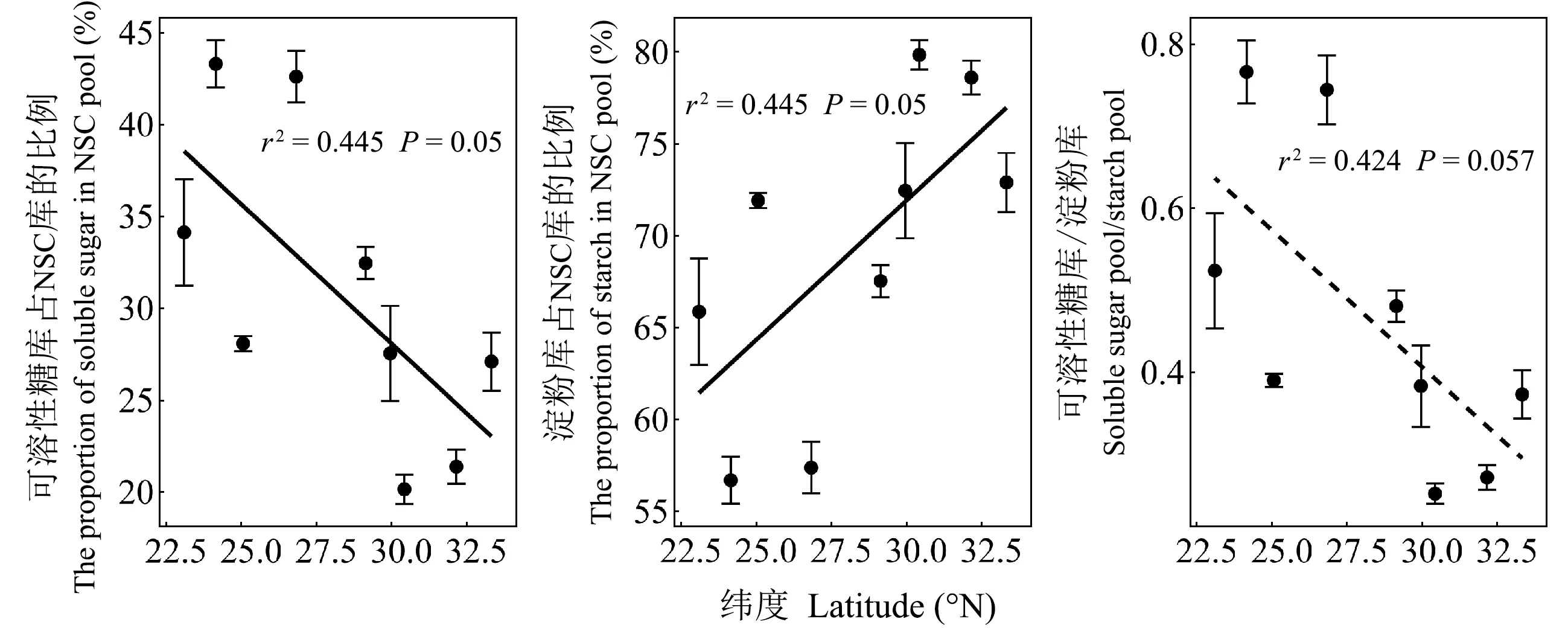
圖4 馬尾松整株可溶性糖庫和淀粉庫占NSC庫的比例以及可溶性糖庫和淀粉庫比值的緯向趨勢

圖5 馬尾松整株的NSC及其組分庫在根、干、枝和葉中分配比例的緯向趨勢
2.4 非結構性碳水化合物庫與環境因子的關系
由表5可知,馬尾松整株的NSC庫、可溶性糖庫和淀粉庫的大小與氣象因子年平均溫度、年均降雨量、年均最冷月最低溫、年均最熱月最高溫、土壤全氮含量和土壤氮磷比顯著正相關,但與帕默爾干旱強度指數、土壤全磷含量、土壤氮鉀比和土壤磷鉀比不相關,可溶性糖庫的大小與土壤全鉀含量正相關(表5)。將顯著的影響因子按氣候(年平均溫度、年均降雨量、年均最冷月最低溫和年均最熱月最高溫)和土壤(全氮、全鉀和氮磷比)分類進行冗余分析方差分解,發現所有因子對NSC庫、可溶性糖庫和淀粉庫的大小變化解釋率高達70.5%,其中氣象因子和土壤因子解釋率分別為26.2%和7.6%,共同作用解釋率為36.7%(圖6)。

表5 馬尾松NSC及其組分庫與環境因子間的相關性

圖6 氣候與土壤對NSC及其組分庫影響的方差分解結果
3 討論
3.1 馬尾松非結構性碳水化合物庫的大小及分配差異
馬尾松淀粉庫的大小平均約占NSC庫的68.9%,顯著高于可溶性糖庫(圖2),表明馬尾松植株整體NSC庫的組成以淀粉庫為主。本研究探討的是生長季節末的NSC儲存情況。在休眠季,NSC以淀粉儲存為主,這與Richardson等(2015)研究中紅櫟(Quercusrubra)整株儲存結果一致。目前,大多數研究側重于器官含量而不是樹木個體NSC庫,發現尤其在枝邊材(Hochetal., 2003)和干邊材(Hochetal., 2003; Richardsonetal., 2013)淀粉含量高于可溶性糖含量。
植物體內NSC的儲存大小和地上地下生物量的積累緊密相關(Sch?nbecketal., 2018)。本研究發現馬尾松整株的NSC、可溶性糖和淀粉都主要儲存在干和根中(圖3, 5),表明樹干和根是馬尾松最重要的NSC儲存器官,也反映出在非生長季植株多余的碳通過枝往樹干和根中進行存儲。樹木器官根據自身各功能需求,體內NSC可以短期或者長期被分配存儲在不同器官中(Richardsonetal., 2013),從而支持自身在呼吸大于光合的情況下(如春季發芽)或在不可預測的壓力(干旱、極端氣候等)下能夠生存(Hochetal., 2003; Kleinetal., 2014)。另外,處于主林層的樹木受光脅迫的概率較小,樹體較大,多年積累致使NSC總量較多(Kobe, 1997; Myersetal., 2007),因此樹干具有最高水平的NSC儲存。
此外,馬尾松NSC庫在地上和地下器官的存儲比例存在差異(表4, 圖5),樹干主要儲存淀粉,而樹根主要儲存可溶性糖。這一差別可能因為生長季末地上部分已停止生長,地上部分的大多數可溶性糖轉化為淀粉,致使樹干淀粉儲存最多。另外,因土壤溫度高于大氣溫度,地下根系生長仍未停止,根系仍需大量可溶性糖用于呼吸消耗。與樹干和根相比,馬尾松葉和枝的儲存比例相對較低(表4, 圖5),這主要和它們的生物量相對偏低有關。此外,枝存儲的NSC主要用來抵御冬季嚴寒和供給來年新葉生長(Kleinetal., 2014)。而常綠樹種的葉則具有碳水化合物生產的“源”和“庫”的雙重功能,即在生長季節因其光合作用是源,在非生長季節則因其儲存大量資源成為庫(Lietal., 2002)。
3.2 馬尾松非結構性碳水化合物庫的緯向特征
馬尾松整株NSC庫、可溶性糖庫和淀粉庫的大小隨緯度增加而降低(圖2),在器官(枝、干、根)的大小也呈現相同趨勢(圖3),與我國不同區域森林碳儲備空間格局的研究結果一致(周玉榮等, 2000; Lietal., 2004; Wenetal., 2016)。本研究表明隨著緯度增加,馬尾松碳供應不足,限制樹木生長。葉片作為植物碳水化合物的合成器官,高緯度的植株通過增加葉片光合物質積累能力來應對相對較短的生長季,進而減弱其NSC存儲的緯向趨勢。然而,可利用性碳庫的大小與樹木生長之間的消長關系一直存在爭議。如倪妍妍等(2017)發現,栓皮櫟幼苗根NSC庫的大小隨緯度增加而增加,并認為樹木通過主動調節體內NSC含量并維持在一定含量范圍,而不影響植株生長(Lietal, 2018)。筆者認為這種差異主要由物種間碳的積累與分配方式(Liuetal., 2018)、物種年齡階段(Guoetal., 2014)以及統計調查空間尺度的大小不同(Wenetal., 2016)造成。此外,本研究還發現,馬尾松整株可溶性糖庫占整個NSC庫的百分比隨緯度增加而降低,導致可溶性糖庫與淀粉庫的比值隨緯度增加而降低(圖5)。這可能是因為相對低溫的環境致使植物活性降低,較高的淀粉儲存比例利于生長在分布北界的樹木將自身的淀粉水解成可溶性糖來抵御冬季嚴寒(Dangetal., 2015)。
3.3 環境對非結構性碳水化合物存儲的影響
溫度和降水對NSC的含量和儲存具有顯著影響(K?rner, 2003; Hochetal., 2012)。在本研究中,馬尾松整株NSC庫、可溶性糖庫和淀粉庫的大小與年平均溫度、年均最冷月最低溫和年均最熱月最高溫正相關(表5)。NSC的儲存大小隨溫度增加而增加,表明低溫直接限制碳活性并影響結構組織的生長,進而限制碳水化合物的積累。樹木儲存的可溶性碳水化合物會用來彌補和緩沖由低溫導致較低的光合產物無法維持生長的不足(Lietal., 2002)。NSC存儲與年均降雨量正相關,但與帕默爾干旱強度指數無顯著相關性(表5)。這表明,干旱脅迫降低森林碳儲備,但植物通過主動調節體內NSC含量并維持在一定的含量范圍來保證植物存活與生長(Lietal., 2016)。如Würth等(2005)發現,熱帶森林維持較高水平的碳供應確保樹木在干旱季節生長不受碳限制。在大地理尺度上,植物往往由磷限制生長逐漸過渡到氮限制狀態(Reichetal., 2004)。本研究發現,NSC庫的大小隨土壤的全氮含量和氮磷比增加而增加,而與全磷含量無關(表5)。這可能是因為馬尾松生長環境土壤養分相對貧瘠,對額外資源采取更積極的利用方式,支持更高的生長率(Zadwornyetal., 2016)。除此之外,本研究發現氣候因子對NSC庫大小的影響高于土壤因子(圖6)。相比于土壤,氣候是樹木生長空間變化的最強驅動力,并認為氣候變化可能對森林生產力和固碳產生重大影響(Toledoetal., 2011),這支持本研究結果。本研究還發現,土壤對馬尾松碳儲存的影響主要通過與氣候共同作用影響(圖6)。在不同空間尺度上,土壤養分與氣候條件的變化可能對樹木生長產生復雜的相互作用(Gorissenetal., 2004)。如Baker等(2003)發現,在水分可利用性較高的情況下,半落葉性樹木生長高于常綠森林,但這種差異僅在足夠的降雨量情況下才明顯。除氣候和土壤外,林分結構、物種組成和植物多樣性等因子均影響樹木NSC的儲存與分配(Tanakaetal., 2016; Chlumskaetal., 2022),但在本文中并未進行探討,有待后續探索。
4 結論
通過在南北樣帶(23.0°—33.5°N)上對9個馬尾松人工林的樹木NSC及其組分庫的大小估算認為,馬尾松植株NSC庫以淀粉儲存為主,NSC庫、可溶性糖庫和淀粉庫分別在干和根中存儲最多; 整株和不同器官中的NSC庫、可溶性糖庫和淀粉庫均呈現出顯著的緯向變化格局; 氣候對馬尾松NSC存儲及分配的影響大于土壤因子。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
