AcrR 突變介導鼠傷寒沙門菌耐藥的調控
2022-12-09 11:19:50李小軍李勝利樊帥奇鄧翔杰孫亞偉
中國獸醫雜志 2022年10期
關鍵詞:耐藥
李小軍,李勝利,樊帥奇,鄧翔杰,孫亞偉
(1.河南省濟源市動物衛生監督所,河南 濟源 454650;2.河南科技學院動物科技學院,河南 新鄉 453003)
沙門菌是引發食物中毒的一類重要病原菌,每年該菌導致全球約9 380萬人感染,15.5萬人死亡[1]。鼠傷寒沙門菌是沙門菌一個重要血清型,是臨床沙門菌病的主要病原。目前,治療沙門菌病的首選藥物是 β-內酰胺類藥物和氟喹諾酮類藥物。然而,由于抗菌藥的不合理使用和誤用,多重耐藥鼠傷寒沙門菌的出現和廣泛傳播已使沙門菌病治療面臨更大的挑戰[2]。
多重耐藥泵是細胞膜內的一類膜蛋白,它可借助ATP酶水解和細胞膜兩側的質子反向轉運提供動力,把進入細胞內的藥物、去污劑和細胞內代謝產物排至細胞外。在鼠傷寒沙門菌體內,AcrAB-TolC是最重要的多重耐藥泵之一。當其在敏感鼠傷寒沙門菌SL1344中失活后,體外用環丙沙星不能篩選出氟喹諾酮類耐藥突變體[3]。當其在臨床多重耐藥鼠傷寒沙門菌BN10055和543SA98中失活后,突變體對萘啶酸、環丙沙星、氯霉素、氟苯尼考和四環素的最小抑菌濃度(Minimum inhibition concentration,MIC)降低8~64倍,表型由耐藥轉變為中介或敏感[4]。細菌多重耐藥產生過程中,AcrAB-TolC是組成性表達,其表達水平主要受RamRA、MarRA、SoxRS 和AcrR調控。其中,RamRA是最重要的調控體系。已證實,RamA能夠直接結合在acrA和tolC啟動子區提高該基因表達[5]。ramR基因位于ramA基因上游,其轉錄方向與ramA基因相反。臨床鼠傷寒沙門菌中,當RamR發生突變或RamR與外排泵底物(溴化乙錠、羅丹明6G和吲哚)結合后,RamR從ramA啟動子區解離,RamA表達,acrAB表達水平增加,細菌對藥物敏感性下降[6-7]。AcrR是AcrAB-TolC的一個負調控蛋白,其蛋白編碼序列位于acrAB開放閱讀框上游,轉錄方向與acrAB相反。目前,AcrR突變、缺失、轉錄提前終止,IS插入等已在大腸桿菌、肺炎克雷伯菌和流感嗜血桿菌染色體內發現,這些基因變化導致了acrAB表達水平增加,細菌對藥物敏感性下降[8-10]。盡管上述研究已證實了AcrR在臨床多重耐藥菌中的作用,然而它在沙門菌中的相關報道卻非常少。
2004年,Hsueh等從豬和人體內分離的環丙沙星耐藥霍亂沙門菌中首次發現AcrR突變(Gln78Stp),但鼠傷寒沙門菌的AcrR沒有發生變化[11]。另外,由鴿子分離的1株敏感鼠傷寒沙門菌 BN18,體外用環丙沙星篩選2株多重耐藥菌BN18/41和BN18/71,其AcrR序列在第75位異亮氨酸(Ile) 第76位谷氨酸(Glu)后出現1個75Ile-76Glu重復。用敏感菌BN18的acrR在該突變體內互補后,麻保沙星、環丙沙星、四環素和氯霉素對細菌的MICs下降4倍[12]。目前AcrR在臨床鼠傷寒沙門菌中是否發揮作用還未見報道。
為了闡明臨床鼠傷寒沙門菌多重耐藥機制,本試驗首先檢測多重耐藥泵抑制劑在4株臨床菌對不同藥物敏感性中作用,并通過實時熒光定量PCR(Real time RT-PCR)檢測細菌內主要多重耐藥泵基因相對表達水平;其次,失活細菌內主要多重耐藥泵及其調控蛋白,比較蛋白失活前后細菌MICs變化,確定該蛋白在細菌多重耐藥中作用;最后,測序分析細菌耐藥相關蛋白基因序列,選取發生突變的耐藥泵相關蛋白并在臨床菌中過表達,確定該蛋白在多重耐藥調控中的作用。
1 材料與方法
1.1 菌株和質粒 鼠傷寒沙門菌CVCC541(命名為ST),購自中國獸醫藥品監察所;臨床鼠傷寒沙門菌CST1、CST2、CST3和 CST4,分別從不同地區不同病雞的肝臟中分離,均通過生化及PCR測序鑒定為鼠傷寒沙門菌。大腸桿菌DH5α,本實驗室保存菌;原核表達質粒 pET-32a,購自Novagen公司;用于基因失活質粒:pKD4,pKD46和pCP20,均購自耶魯大學大腸桿菌菌種中心。
1.2 藥品及試劑 阿莫西林 (Amoxicillin,AMO)、頭孢噻呋鈉(Ceftiofur sodium,CF)、頭孢噻肟 (Cefotaxim,CTX)、鹽酸四環素 (Tetracycline hydrochloride,TET)、多西環素 (Deoxycycline,DOX)、氟苯尼考(Florfenicol,FLO)、環丙沙星(Ciprofloxacin,CIP)、恩諾沙星 (Enrofloxacin,ENR) 和加替沙星 (Gatifloxacin,GAT),均購自中國獸醫藥品監察所;多重耐藥泵抑制劑苯丙氨酸-精氨酸-β-萘胺(Phenylalanine arginine beta-naphthylamide,PAβN)和羰基氰化物間氯苯腙 (Carbonyl cyanide m-chlorophenylhydrazone,CCCP),均購自西格瑪奧德里奇(上海)貿易有限公司 (Sigma-Aldrich);質粒小量提取試劑盒和凝膠回收試劑盒,均購自天根生化科技(北京)有限公司;PCR 2×Taq預混體系和DNA Marker,均購自康為世紀生物科技股份有限公司;細菌總RNA提取試劑盒,購自美國普洛麥格公司(Promega);Superscript Ⅲ 反轉錄酶,購自美國英杰生命技術有限公司(Invitrogen);2×SYBR Green Supermix,購自寶生物工程(大連)有限公司;胰蛋白胨、酵母提取物、MH培養基和營養瓊脂,均購自青島高科技工業園海博生物技術有限公司;NaCl、Na2HPO4、KCl和KH2PO4,均購自中國醫藥集團有限公司。
1.3 主要儀器設備 低溫離心機(型號:7146)、微量核酸蛋白濃度測定儀(型號:NanoDrop),美國賽默飛世爾科技(Thermo Scientific);電轉儀(型號:Gene Pulser Xcell)、熒光定量PCR儀(型號:CFX96),美國Bio-Rad公司;梯度基因擴增儀(型號:Biometra Tone 96G),德國耶拿公司;生物安全柜(型號:BHC-1300IIA2),蘇州金凈凈化設備科技有限公司。
1.4 引物設計 鼠傷寒沙門菌相關引物均依據NCBI公布的SalmonellatyphimuriumLT2 (GenBank登錄號:NC_003197) 基因組序列,利用Primer primer 5軟件設計,由武漢金開瑞生物工程有限公司合成。試驗所用引物信息見表1。

表1 引物信息

注:下劃線:AcrR表達用引物攜帶的酶切位點
1.5 基因失活及噬菌體轉導 按照黃慧等[13]方法構建ST的acrB、ramA、marA和soxS基因失活菌,同源重組打靶DNA片段擴增引物和基因失活菌檢測引物均見表1。基因失活菌陽性PCR擴增產物純化后送武漢金開瑞生物工程有限公司測序。
通過 P22噬菌體將基因失活菌中失活目的片段按照已報道的試驗方法傳導給臨床菌[14]。(1)制備攜帶基因失活序列的噬菌體。將100 μL的P22噬菌體與10 mL對數生長期的基因失活菌混合,37 ℃、220 r/min振蕩培養6~8 h。細菌裂解后,加入100 μL 氯仿,繼續振蕩培養20 min。隨后,將菌液6 500 r/min離心20 min,棄去細胞碎片,吸取上清液以 0.22 μm濾膜過濾即得攜帶失活基因片段的噬菌體,4 ℃保存備用。(2)噬菌體轉導。無菌操作臺內吸取臨床鼠傷寒沙門菌過夜培養物200 μL分別加入到3個1.5 mL離心管內,分別加入1、5 μL和20 μL攜帶失活基因片段的噬菌體,同時設置1管不加噬菌體作為空白對照,1管僅加入20 μL攜帶失活序列的噬菌體作為陰性對照。將5個離心管37 ℃靜止20 min,隨后每個離心管加入LB液體培養基,使其總體積為1 mL。將5個離心管于37 ℃、220 r/min培養45 min,菌液6 500 r/min離心5 min 棄上清,用200 μL LB液體培養基懸浮沉淀,分別涂布于含有50 μg/mL 卡那霉素的LB平板上,37 ℃培養過夜。(3)陽性轉導子鑒定。挑取篩選平板的單克隆菌在含有卡那霉素(50 μg/mL)的LB液體培養基內過夜培養后,利用基因失活菌檢測引物 (表1)進行PCR檢測,隨后將陽性PCR純化產物送武漢金開瑞生物工程有限公司測序。檢測不同抗菌藥物對測序確證的acrB失活菌、ramA失活菌、marA失活菌和soxS失活菌的MICs。
1.6 MICs檢測 具體方法按照美國臨床和實驗室標準協會(Clinical and Laboratory Standards Institute,CLSI)標準[15]。挑取待測菌 (表2) 單克隆,由LB液體培養基培養至對數期,稀釋菌液使其在MH液體中的濃度為1×105~5×105CFU/mL。將稀釋菌液添加入96孔微量板中,將不同濃度的抗菌藥物溶液進行2倍倍比稀釋,放置于37 ℃培養16~18 h,觀察結果以眼觀無細菌生長所在孔的藥物濃度值即為該藥物的MICs。另外,檢測在多重耐藥泵抑制劑 PAβN(工作濃度,20 mg/L)或CCCP(工作濃度,70 mg/L)存在時藥物對ST、CST1、CST2、CST3和CST4的MICs。本試驗中每次檢測均以大腸桿菌ATCC25922為質控菌,每個試驗重復3次,任何2次試驗數值差異均不超過1個梯度,試驗結果取3次平均值。
1.7 多重耐藥泵基因表達水平檢測 取2 mL對數生長期的ST、CST1、CST2、CST3和CST4菌液,按照細菌總RNA提取試劑盒按照說明書步驟提取總RNA。獲得的總RNA經DNA酶消化后,使用16S rRNA引物(表1)通過PCR檢測總RNA提取物中DNA是否消化完全,并利用NanoDrop One/OneC微量核酸蛋白濃度測定儀測定總RNA濃度。
利用Superscript Ⅲ 反轉錄酶將1 μg 總RNA按照說明書步驟體外反轉錄為cDNA。隨后,取1 μL cDNA為模板,與熒光定量檢測引物和2×SYBR Green Supermix 在冰上避光混合,通過熒光定量PCR儀檢測cDNA中各基因信使RNA (mRNA) 的Ct值。擴增體系(25 μL):2×SYBR Green Supermix 12.5 μL,上、下游引物各1 μL,cDNA模板 1 μL,超純水 9.5 μL。擴增程序:預變性,95 ℃ 30 s;變性,95 ℃ 5 s;退火及延伸,59 ℃ 20 s,共40個循環;充分延伸,72 ℃ 10 min;降溫,16 ℃ 30 min。每個樣品3個重復,最少檢測3次。所獲Ct值通過 2-ΔΔCt方法計算臨床菌所測基因(表1)相對ST菌的變化倍數。
1.8 喹諾酮耐藥決定區和多重耐藥泵調控蛋白序列檢測 以臨床菌CST1、CST2、CST3和CST4菌液為模板,PCR擴增其各自喹諾酮耐藥決定區(gyrA、gyrB、parC和parE)和多重耐藥泵調控蛋白編碼區(ramRA、marRA、soxRS和acrR)基因序列。PCR擴增體系(50 μL):2×PCR Mixture 25 μL,上、下游引物各1 μL,菌液1 μL,ddH2O 22 μL。PCR擴增程序:預變性,95 ℃ 3 min;變性,95 ℃ 30 s;退火,30 s (退火溫度:gyrA、gyrB、parC和parE為55 ℃;ramRA、marRA和acrR為53 ℃;soxRS為59 ℃),共 30個循環;充分延伸,72 ℃ 10 min;降溫,16 ℃ 30 min。PCR產物純化后送武漢金開瑞生物工程有限公司測序。
1.9 AcrR表達載體構建及互補 以ST菌液為模板,按1.8方法擴增acrR基因序列。隨后,以純化的acrR序列為模板利用引物AR32F和AR32R(表1)擴增其開放閱讀框。純化后的PCR產物通過BamHⅠ和XhoⅠ雙酶切,其回收產物與同樣雙酶切的pET-32a載體連接。連接產物電轉DH5α感受態細胞,轉化子在含有100 μg/mL 氨芐青霉素的LB平板上篩選。篩選的轉化子通過PCR擴增測序確定陽性重組質粒。最后,將陽性重組質粒電轉入臨床菌CST2、CST3和CST4中并誘導表達,檢測AcrR過表達后所測藥物對上述細菌的MICs。
2 結果
2.1 多重耐藥泵抑制劑對MICs的影響 多重耐藥泵抑制劑PAβN和CCCP存在時,不同抗菌藥物對鼠傷寒沙門菌的MICs見表2。PAβN導致鹽酸四環素、多西環素、加替沙星和恩諾沙星對ST、CST1、CST2、CST3和CST4的MICs下降3.00~26.67倍;氟苯尼考對CST1和CST2的MICs分別下降3.00倍和5.19倍;環丙沙星對CST3的MICs下降6.25倍。另外,CCCP導致氟苯尼考對ST、CST1和CST2的MICs下降4.67~5.19倍;鹽酸四環素對ST、CST1和CST3的MICs下降6.00~13.32倍;多西環素對ST和CST1的MICs分別下降8.04倍和3.20倍;但CCCP 沒有導致所測藥物對CST4的MICs發生明顯改變。
2.2 多重耐藥泵基因相對表達水平 臨床菌相對ST多重耐藥泵基因表達水平結果見圖1。4株臨床菌acrA表達水平相對ST增加了3~9倍。CST2、CST3和CST4中,多重耐藥泵編碼基因acrF、mdfA和emrB表達水平也增加5~15倍。但外膜通道蛋白編碼基因tolC僅在CST1和CST4中表達水平增加3倍以上。mdtK僅在CST2中表達水平增加3倍以上。

圖1 臨床鼠傷寒沙門菌多重耐藥泵基因相對表達水平
2.3 AcrB及其調控蛋白在藥物敏感性中作用 MICs測定結果見表2。acrB缺失導致所測藥物除阿莫西林外對ST和CST1的MICs下降3.33~80.00倍;所測藥物除阿莫西林和環丙沙星外對CST4的MICs下降3.54~37.66倍;頭孢噻呋鈉、頭孢噻肟、鹽酸四環素和多西環素對CST2和CST3的MICs下降3.00~26.75倍。當ramA、marA和soxS基因在細菌中被分別失活后,所有藥物對基因失活前后細菌的MICs變化均沒有超過3倍。

表2 鼠傷寒沙門菌在多重耐藥泵抑制劑、多重耐藥泵AcrB和多重耐藥泵調控蛋白存在或缺失時MICs
2.4 喹諾酮耐藥決定區和多重耐藥泵調控蛋白靶位點突變 序列檢測結果見表3。4株臨床菌在GyrA均出現Gly133→Glu突變。CST2在ParC出現了Thr57→Ser突變,CST3在GyrA和ParC分別出現了Asp87→Gly和Thr57→Ser突變。另外,CST2、CST3 和CST4的AcrR也發現了突變。其中,CST2和CST3 的AcrR在第216位氨基酸發生了突變(Ser→Ala);CST4中acrR第235位堿基G缺失,導致其在第267位形成TAA終止密碼子,acrR基因轉錄提前終止。CST4中acrR基因序列經NCBI中BLAST對比,沒有發現100%同源序列,上傳至NCBI數據庫獲得序列號為ON614636。另外,AcrAB其他相關調控蛋白(RamRA、MarRA和SoxRS) 均未出現突變。
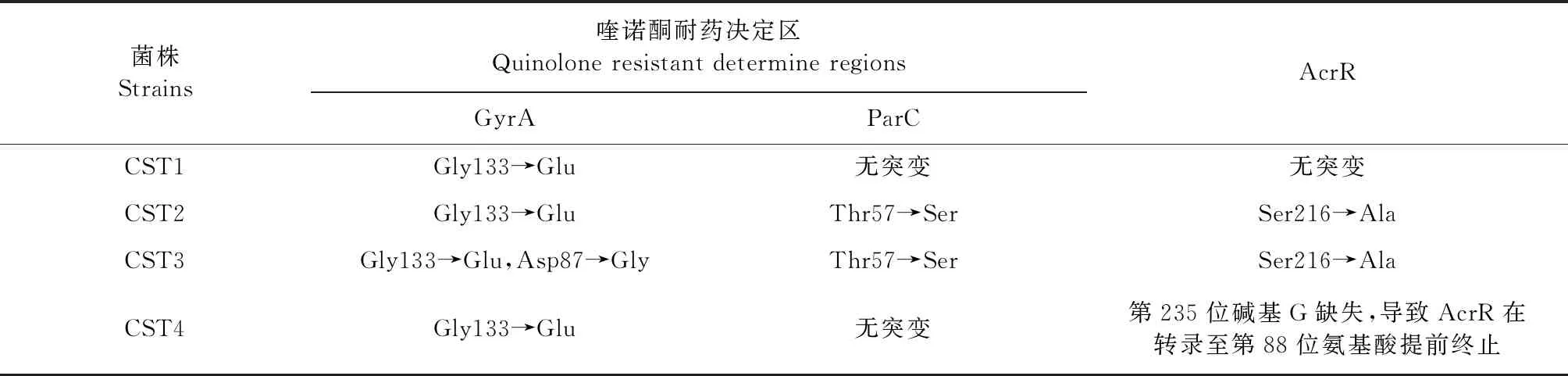
表3 臨床菌喹諾酮耐藥決定區和多重耐藥泵調控蛋白AcrR靶位點突變
2.5 AcrR表達載體構建及互補 ST菌acrR基因通過pET-32a載體構建重組質粒pET-32a-acrR,該質粒在臨床菌CST2、CST3和CST4中過表達后,鹽酸四環素對CST2的MICs下降3.00倍,頭孢噻呋鈉、鹽酸四環素和多西環素對CST3的MICs下降3.98~21.40倍;恩諾沙星和加替沙星對CST4的MICs分別下降4.00倍和3.00倍 (表4)。

表4 臨床菌及其過表達AcrR后MICs
3 討論
多重耐藥泵抑制劑PAβN能夠抑制AcrAB和AcrEF表達,導致多個藥物對細菌的MICs顯著下降[16]。本試驗發現,PAβN導致鹽酸四環素、多西環素、恩諾沙星和加替沙星對ST、CST1、CST2、CST3和CST4的MICs下降3.00~26.67倍;頭孢噻呋鈉和頭孢噻肟MICs沒有發生明顯改變。Nikaido等研究顯示,鼠傷寒沙門菌AcrB能夠外排 β-內酰胺類藥物且外排能力與該藥物含有親脂側鏈的比例有關[17]。試驗發現,當acrB在ST、CST1、CST2、CST3和CST4中失活后,頭孢噻呋鈉和頭孢噻肟對所有失活菌的MICs下降3.50~80.00倍。推測PAβN不能有效增加頭孢噻呋鈉和頭孢噻肟敏感性的原因是這2個抗菌藥與AcrB結合能力較強或它們與AcrB結合位點和PAβN與AcrB結合位點不同,故PAβN與AcrB結合后不影響2個抗菌藥物外排。Lamers等研究發現,10 mg/L PAβN很難有效增加 β-內酰胺類藥物對銅綠假單胞菌的敏感性,當PAβN濃度為25 mg/L 或 50 mg/L時可顯著增加該類藥物對銅綠假單胞菌的敏感性,但該增效作用是由于PAβN增加了細胞膜對 β-內酰胺類藥物的通透性而非抑制多重耐藥泵表達[18]。CCCP是 H+螯合劑,它通過結合H+降低細胞膜兩側離子濃度差,減少耐藥泵轉運藥物動力從而抑制細菌多重耐藥。在鼠傷寒沙門菌體內,MATE家族MdtK依賴Na+反向轉運提供的動力將藥物從細胞內排至細胞質間隙[19]。同時,MFS家族MdfA依賴H+和其他質子(Na+或K+)反向轉運提供的動力將藥物從細胞內排至細胞外,特別是在外排中性藥物(如氯霉素)時主要依賴Na+或K+反向轉運提供的動力[20]。本試驗臨床菌CST2、CST3和CST4中mdfA和CST2中mdtK表達水平相對ST顯著增加,這可能是CCCP對這3株臨床菌抑制作用減弱甚至消除的主要原因。
細菌對喹諾酮類的耐藥機制主要包括喹諾酮耐藥決定區靶位點突變、活性多重耐藥泵和質粒介導。本試驗中的臨床鼠傷寒沙門菌CST2在ParC發生了Thr57→Ser突變,CST3除上述的ParC突變外,在GyrA還發生了Asp87→Gly突變,這些突變在已報道的臨床鼠傷寒沙門菌都曾檢測到,它們可導致氟喹諾酮類藥物敏感性降低[21]。另外,4株臨床菌在GyrA都出現了Gly133→Glu突變,該突變在傷寒沙門菌中常見,而在鼠傷寒沙門菌中并不常見[22]。比利時對全國沙門菌的耐藥性調研結果顯示,1 200余株鼠傷寒沙門菌GyrA攜帶Gly133→Glu突變,該突變不導致鼠傷寒沙門菌對喹諾酮類藥物敏感性發生改變[23]。
AcrR是AcrAB的負調控蛋白之一,它以同源二聚體形式與所調控DNA結合,抑制基因表達。鼠傷寒沙門菌AcrR晶體結構學研究表明,每個AcrR單體由N端的DNA結合域(Met1-Lys55)和C端的底物結合域(Ser56-Ala210) 構成。其中,C端底物結合域第4個 α螺旋中的第67位谷氨酸(Glu)與第7個α螺旋中的第130位甘氨酸(Gln)可形成氫建,該氫鍵對維持AcrR底物結合位點的穩定性發揮重要作用。當AcrR與底物結合后,蛋白C端底物結合域空間結構發生改變,導致其N端的DNA結合域發生改變,AcrR從基因序列上解離,受抑制基因的表達水平增加[24]。對臨床多重耐藥大腸桿菌的研究發現,AcrR突變檢出率要高于MarA和SoxS,其序列與鼠傷寒沙門菌AcrR序列比較,2個序列N端209個氨基酸表現89%同源性,剩余的19個氨基酸僅有1個相同[25]。目前在臨床鼠傷寒沙門菌中AcrR突變很少被發現,其在多重耐藥鼠傷寒沙門菌中的作用還不清楚。本試驗在臨床鼠傷寒沙門菌CST2、CST3和CST4中發現AcrR突變,當ST的acrR基因在這3株菌中過表達后,鹽酸四環素對CST2,頭孢噻呋鈉、鹽酸四環素和多西環素對CST3,恩諾沙星和加替沙星對CST4的MICs均顯著下降,說明AcrR參與了臨床鼠傷寒沙門菌多重耐藥的調控,該結論豐富了臨床鼠傷寒沙門菌多重耐藥機制研究的數據。
總之,本試驗發現AcrB是臨床鼠傷寒沙門菌多重耐藥的主要原因,負調控蛋白AcrR突變導致其從acrAB啟動子區解離,acrAB表達水平增加,細菌多重耐藥性增強,這可能是臨床鼠傷寒沙門菌多重耐藥的主要分子機制。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
