膠東地區(qū)設(shè)施栽培奧尼爾和米斯蒂藍(lán)莓花色苷代謝研究
2022-11-17 06:28:02劉笑宏張麗麗王春燕慈志娟于海軍李公存顧亮趙玲玲
山東農(nóng)業(yè)科學(xué) 2022年10期
劉笑宏,張麗麗,王春燕,慈志娟,于海軍,李公存,顧亮,趙玲玲
(1.山東省煙臺(tái)市農(nóng)業(yè)科學(xué)研究院,山東 煙臺(tái) 265500;2.中華全國供銷合作總社濟(jì)南果品研究所,山東 濟(jì)南 250000;3.海陽市果業(yè)發(fā)展服務(wù)中心,山東 海陽 265100)
藍(lán)莓是我國主要生態(tài)經(jīng)濟(jì)林漿果,營養(yǎng)物質(zhì)含量豐富,風(fēng)味獨(dú)特,被評為“人類五大健康食品之一”[1],列入世界第3代水果行列[2]。花色苷是藍(lán)莓重要的品質(zhì)性狀,其生物合成受內(nèi)部因素(合成途徑關(guān)鍵酶活性及相關(guān)基因表達(dá)量等)和外界環(huán)境因子(溫度、空氣濕度、光照等)的共同影響[3-5]。研究表明,苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)是花色苷合成過程的起始酶,起限速作用;查耳酮合酶(chalcone synthase,CHS)可催化生成柚皮素查爾酮;查耳酮異構(gòu)酶(chalcone isomerase,CHI)可顯著提高花色苷合成速率;黃烷酮3-羥化酶(flavanone 3-h(huán)ydroxylase,F(xiàn)3H)是花色苷合成途徑的中樞酶;二氫黃酮醇4-還原酶(dihydroflavonol 4-reductase,DFR)對花色苷積累起限制因素;花色素合成酶(anthocyanidin synthase,ANS)是花色苷合成和原花青素合成樞紐的關(guān)鍵酶,決定花色苷合成途徑的走向;類黃酮糖基轉(zhuǎn)移酶(UDP-glucoseflavonoid 3-O-glucosyltransferase,UFGT)是花色苷合成的最終作用酶;糖苷水解酶(glycoside hydrolases,GLH)和多酚氧化酶(polyphenol oxidase,PPO)對花色苷的分解氧化起重要作用[6-10]。基因是決定酶活性的關(guān)鍵因素,環(huán)境因素則通過影響酶活性影響花色苷的最終合成。環(huán)境因素中,光照是花色苷合成的重要前提[11,12],適量光照可有效提高花色苷的合成量,而強(qiáng)光則會(huì)降低花色苷的含量[13];溫度是影響花色苷代謝的第二因素[14,15],高溫能降解花色苷[16,17],低溫則抑制花色苷的合成[18];空氣濕度對花色苷的積累同樣重要,較高的空氣相對濕度可降低蒸騰速率、提高光合產(chǎn)物積累,從而為花色苷代謝提供充足的合成底物[19,20]。
膠東半島氣候溫暖濕潤,是我國重要的藍(lán)莓生產(chǎn)區(qū)[21],傳統(tǒng)以栽植北高叢藍(lán)莓為主。近年來,南高叢藍(lán)莓由于品質(zhì)優(yōu)、適應(yīng)性強(qiáng)、豐產(chǎn)性高等特點(diǎn),被大規(guī)模引種,但因花期較早,易受霜凍危害,在膠東地區(qū)較為適合設(shè)施栽培。奧尼爾和米斯蒂均為引入的優(yōu)良南高叢藍(lán)莓品種,奧尼爾果個(gè)大、香氣濃郁,米斯蒂有特殊香氣,均深受消費(fèi)者喜愛,表現(xiàn)出較高的商品價(jià)值。但引種后生長環(huán)境的改變?nèi)菀讓?dǎo)致藍(lán)莓果實(shí)中營養(yǎng)物質(zhì)成分及含量變化,從而對其品質(zhì)造成一定影響。花色苷作為藍(lán)莓的主要活性物質(zhì),其含量改變明顯影響藍(lán)莓果實(shí)品質(zhì),因此,了解生長環(huán)境條件對花色苷積累和代謝的影響對于改善藍(lán)莓品質(zhì)具有重要意義。
本試驗(yàn)以膠東地區(qū)設(shè)施栽培的奧尼爾和米斯蒂兩個(gè)品種藍(lán)莓為研究對象,分析轉(zhuǎn)色期各階段花色苷積累及代謝途徑相關(guān)酶基因表達(dá)的情況,并探究設(shè)施環(huán)境因子對花色苷積累的影響,以期為藍(lán)莓花色苷代謝研究及生產(chǎn)管理提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)地概況
試驗(yàn)于2021年在山東省煙臺(tái)市福山區(qū)藍(lán)莓生產(chǎn)基地進(jìn)行。福山區(qū)位于膠東半島北部,屬暖溫帶東亞季風(fēng)區(qū)大陸性氣候,夏季溫暖濕潤,冬季寒冷干燥,年平均降水729.2 mm,平均氣溫11.8℃,平均日照2672.2 h,空氣相對濕度65%,適合藍(lán)莓生長發(fā)育。
1.2 試驗(yàn)材料
供試藍(lán)莓品種為奧尼爾和米斯蒂,均于2017年定植于東西向暖棚內(nèi),供試品種南北向種植,株距0.8 m,行距1.8 m,栽培管理措施一致。每個(gè)小區(qū)由4個(gè)種植行組成,按種植行從東到西、植株從南到北的次序,每一種植行順次選擇5株藍(lán)莓(即第一行的5株在種植行的南邊、第二行5株接著第一行的次序向北數(shù)、第三行和第四行順次進(jìn)行),共計(jì)20株設(shè)置為一小區(qū),重復(fù)3次,總計(jì)60株。
1.3 試驗(yàn)方法
1.3.1 環(huán)境因子測定 于果實(shí)轉(zhuǎn)色前每小區(qū)中間部位放置兩個(gè)溫濕度及光照記錄儀,懸掛于植株中部外測5~10 cm處,待果實(shí)總體轉(zhuǎn)色完成后取下記錄儀,數(shù)據(jù)用于設(shè)施環(huán)境因子分析。
1.3.2 藍(lán)莓果實(shí)樣品采集 根據(jù)試驗(yàn)區(qū)內(nèi)藍(lán)莓果實(shí)總體轉(zhuǎn)色情況,分別于轉(zhuǎn)色初期、花盤轉(zhuǎn)色期、轉(zhuǎn)色中期、轉(zhuǎn)色后期、轉(zhuǎn)色完成期采摘果實(shí)樣品,每個(gè)品種每小區(qū)采集50粒,帶回實(shí)驗(yàn)室后液氮冷凍,存于-80℃冰箱供相關(guān)指標(biāo)測定。轉(zhuǎn)色期各階段果實(shí)采集標(biāo)準(zhǔn)見圖1。
圖1 試驗(yàn)材料采收標(biāo)準(zhǔn)
1.3.3 花色苷含量及其代謝相關(guān)基因表達(dá)量測定 藍(lán)莓果實(shí)花色苷含量參照張美嬌等[22]的方法測定。總RNA提取參照鄭嘉偉等[23]的方法,利用北京康為試劑公司的UltraSYBR Mixture進(jìn)行qRT-PCR檢測,相關(guān)基因表達(dá)量檢測引物序列如表1所示。基因相對表達(dá)量數(shù)據(jù)均以各品種轉(zhuǎn)色初期的表達(dá)量為對照。

表1 藍(lán)莓花色苷代謝相關(guān)基因表達(dá)量檢測的引物及序列
1.4 數(shù)據(jù)處理與統(tǒng)計(jì)分析
采用Microsoft Excel 2010對數(shù)據(jù)進(jìn)行處理及作圖,采用DPS 7.05進(jìn)行方差分析(P<0.05)。
2 結(jié)果與分析
2.1 兩種藍(lán)莓果實(shí)轉(zhuǎn)色期花色苷含量變化動(dòng)態(tài)
由圖2可見,奧尼爾的花色苷總量在轉(zhuǎn)色初期至花盤轉(zhuǎn)色期均較低,變化不顯著;轉(zhuǎn)色中期始花色苷快速積累,含量增加顯著,為花盤轉(zhuǎn)色期的5.30倍;轉(zhuǎn)色后期花色苷含量為轉(zhuǎn)色中期的3.38倍,至轉(zhuǎn)色完成期,花色苷含量達(dá)5.95 mg/kg,比轉(zhuǎn)色后期顯著提升30.86%。米斯蒂的花色苷快速積累期稍有延后,轉(zhuǎn)色中期前積累很少,含量變化不顯著;轉(zhuǎn)色后期花色苷快速積累至3.00 mg/kg,為轉(zhuǎn)色中期的8.82倍,至轉(zhuǎn)色完成期花色苷含量達(dá)到5.03 mg/kg。由此可見,奧尼爾的花色苷快速積累期在花盤轉(zhuǎn)色期至轉(zhuǎn)色后期,米斯蒂的快速積累期則在轉(zhuǎn)色中期至轉(zhuǎn)色后期;奧尼爾的花色苷快速積累持續(xù)時(shí)間更長,轉(zhuǎn)色完成時(shí),花色苷含量也更高。
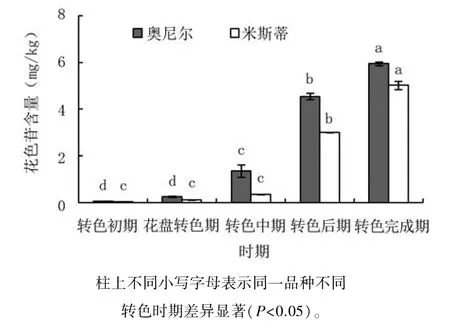
圖2 兩種藍(lán)莓果實(shí)轉(zhuǎn)色期花色苷含量變化動(dòng)態(tài)
2.2 兩種藍(lán)莓果實(shí)轉(zhuǎn)色期花色苷代謝途徑相關(guān)基因表達(dá)量變化
2.2.1 奧尼爾的相關(guān)基因表達(dá)量變化 對奧尼爾花色苷代謝途徑相關(guān)基因不同時(shí)期表達(dá)量分析可見(圖3),花色苷合成基因VcPAL、VcCHS、Vc-CHI、VcANS、VcUFGT表達(dá)量在整個(gè)測定期呈現(xiàn)上升趨勢,轉(zhuǎn)色中期表達(dá)量驟升為花盤轉(zhuǎn)色期的2.77~3.63倍,其中,VcPAL、VcCHS、VcANS和VcUFGT表達(dá)量在轉(zhuǎn)色后期至轉(zhuǎn)色完成期依舊顯著上升,即合成酶繼續(xù)發(fā)揮合成作用。VcF3H和VcDFR表達(dá)量呈現(xiàn)先上升后下降的趨勢,其中,VcF3H在轉(zhuǎn)色后期表達(dá)量最高,轉(zhuǎn)色完成期表達(dá)量顯著下降至轉(zhuǎn)色后期的45.32%;VcDFR表達(dá)量則在轉(zhuǎn)色中期達(dá)最高峰,轉(zhuǎn)色后期至轉(zhuǎn)色完成期表達(dá)量分別顯著下降4.59%、20.51%。氧化酶基因VcGLH和VcPPO表達(dá)量在整個(gè)轉(zhuǎn)色期均較低,呈現(xiàn)轉(zhuǎn)色初期較高、之后下降、轉(zhuǎn)色完成期再顯著回升的趨勢,其中,VcGLH表達(dá)量在花盤轉(zhuǎn)色期比轉(zhuǎn)色初期顯著下降79.39%,花盤轉(zhuǎn)色期至轉(zhuǎn)色后期無顯著變化,轉(zhuǎn)色完成期顯著提升至轉(zhuǎn)色后期的2.08倍;VcPPO的表達(dá)量在轉(zhuǎn)色初期至花盤轉(zhuǎn)色期有所降低,但差異不顯著,轉(zhuǎn)色中期顯著下降52.32%,轉(zhuǎn)色后期與轉(zhuǎn)色中期水平相當(dāng),轉(zhuǎn)色完成期驟升至轉(zhuǎn)色后期的6.83倍。
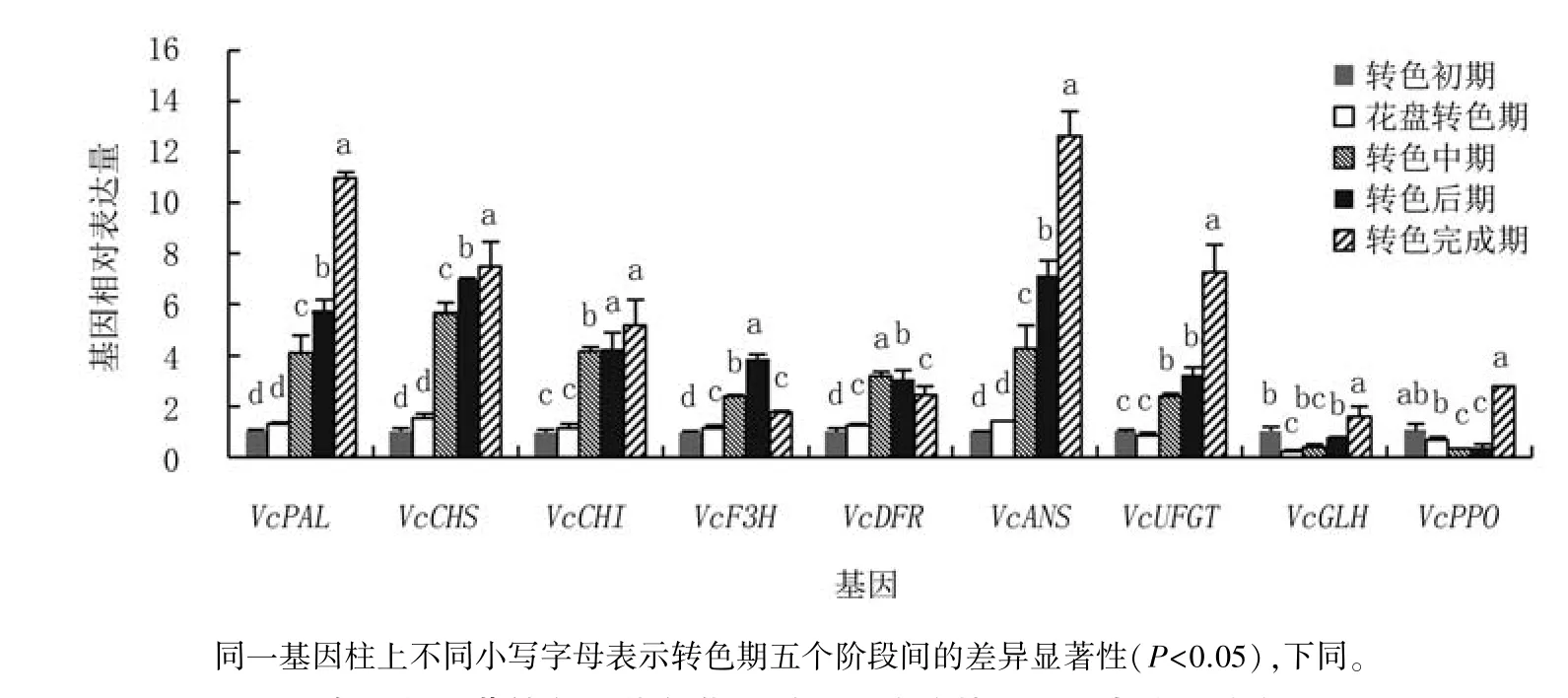
圖3 奧尼爾藍(lán)莓轉(zhuǎn)色期花色苷代謝相關(guān)酶的基因相對表達(dá)量動(dòng)態(tài)分析
可 見,奧 尼 爾 的VcPAL、VcCHS、VcCHI、VcANS、VcUFGT等合成酶基因在果實(shí)轉(zhuǎn)色期持續(xù)發(fā)揮作用,且表達(dá)量較高,而氧化酶基因VcGLH、VcPPO在整個(gè)轉(zhuǎn)色期表達(dá)量均較低,尤其合成酶基因表達(dá)高峰期極低,這更利于果實(shí)花色苷的合成與積累。
2.2.2 米斯蒂的相關(guān)基因表達(dá)量變化 對米斯蒂花色苷代謝途徑相關(guān)基因不同時(shí)期表達(dá)量分析可見(圖4),VcPAL、VcCHS、VcCHI、VcF3H、VcDFR的基因表達(dá)量均呈現(xiàn)先上升后下降的趨勢,轉(zhuǎn)色中后期表達(dá)量驟升,轉(zhuǎn)色后期表達(dá)量最高,而VcUFGT僅在轉(zhuǎn)色后期表達(dá)量驟升;至轉(zhuǎn)色完成期上述基因的表達(dá)量又顯著下降82.00%~91.30%,與花盤轉(zhuǎn)色期的表達(dá)量相當(dāng)。VcANS的表達(dá)量在花盤轉(zhuǎn)色期顯著升高至轉(zhuǎn)色初期的8.25倍,之后表達(dá)量無顯著變化。VcGLH和VcPPO相較于其他基因,表達(dá)量一直處于較低水平。
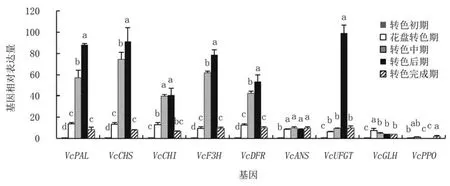
圖4 米斯蒂藍(lán)莓花色苷代謝相關(guān)基因相對表達(dá)量動(dòng)態(tài)分析
由此可見,米斯蒂各基因的表達(dá)趨勢一致性較強(qiáng),且調(diào)控花色苷合成的基因表達(dá)高峰出現(xiàn)在轉(zhuǎn)色中期和轉(zhuǎn)色后期,以轉(zhuǎn)色后期表達(dá)量更高,轉(zhuǎn)色完成時(shí),各基因的相對表達(dá)量與花盤轉(zhuǎn)色期水平相當(dāng),但VcANS的相對表達(dá)量在花盤轉(zhuǎn)色期上升后,一直保持在轉(zhuǎn)色初期的17倍左右。
2.3 果實(shí)微環(huán)境對花色苷積累的影響
2.3.1 光照 從轉(zhuǎn)色期光照強(qiáng)度(圖5)來看,4月11日至21日設(shè)施大棚內(nèi)的光照強(qiáng)度主要集中在20~40 klx,最強(qiáng)光照為40~60 klx,最高達(dá)65.25 klx,此時(shí)大部分藍(lán)莓果實(shí)處于轉(zhuǎn)色初期;4月22日至25日光照強(qiáng)度顯著下降,大多在10 klx左右,最大光照強(qiáng)度為46.40 klx;4月26日至5月1日光照強(qiáng)度波動(dòng)較大,主要分布在10~40 klx,但出現(xiàn)超過60 klx的強(qiáng)光照天氣數(shù)增多,即轉(zhuǎn)色中后期光照強(qiáng)度相對較強(qiáng)。
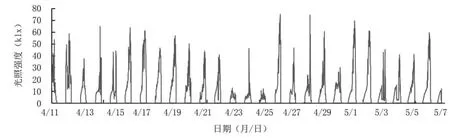
圖5 藍(lán)莓果實(shí)轉(zhuǎn)色期光照強(qiáng)度動(dòng)態(tài)分析
2.3.2 氣溫 由圖6可見,4月11日至21日,設(shè)施內(nèi)氣溫相對穩(wěn)定,大部分維持在15~35℃,溫差較小;4月26日以后,棚內(nèi)溫差較大,氣溫波動(dòng)較大,35℃以上高溫和10℃以下低溫同日出現(xiàn),甚至出現(xiàn)5℃以下低溫。即藍(lán)莓花盤轉(zhuǎn)色期棚內(nèi)溫差較小,轉(zhuǎn)色中后期溫度相對穩(wěn)定,后期晝夜溫差增大,促進(jìn)花色苷合成與積累。由日平均溫度曲線可見,設(shè)施內(nèi)日平均溫度維持在15~25℃,前期維持在20~25℃,后期降至15~20℃。
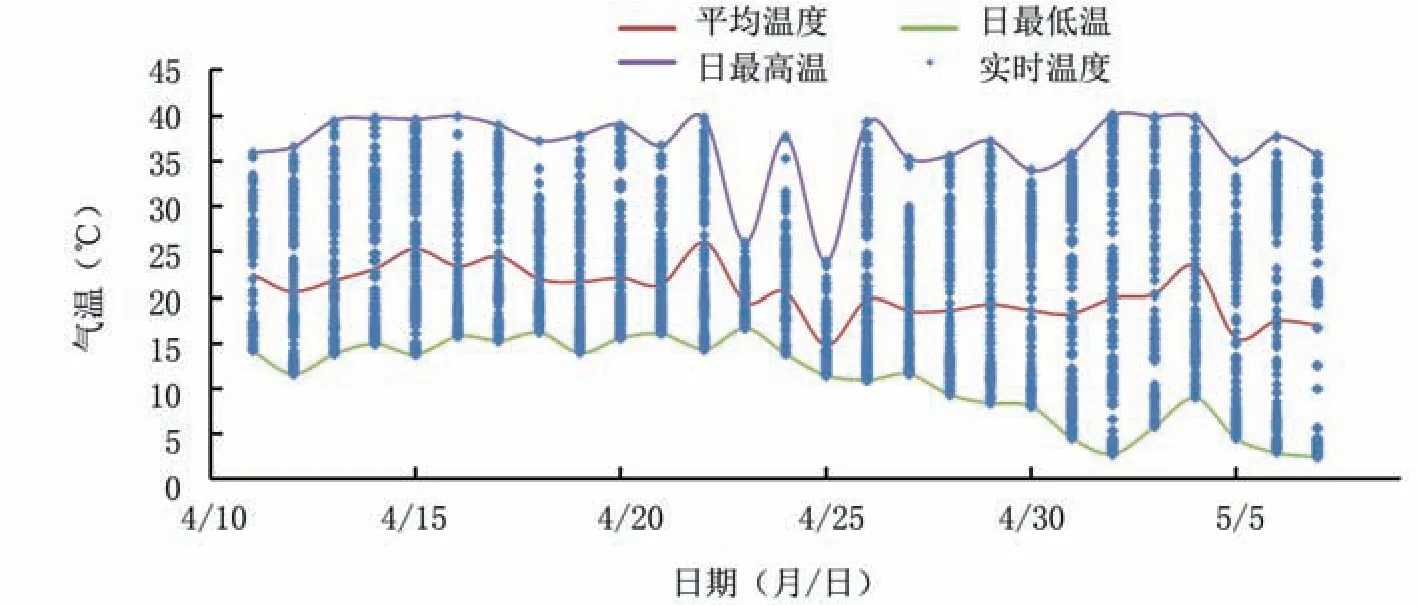
圖6 藍(lán)莓果實(shí)轉(zhuǎn)色期氣溫變化動(dòng)態(tài)
2.3.3 空氣濕度 從圖7來看,設(shè)施內(nèi)空氣濕度大部分在10%以上,4月25日左右濕度全天較高,維持在50%以上,即果實(shí)轉(zhuǎn)色中期生長環(huán)境濕度較大,這有利于減少光合產(chǎn)物消耗及為花色苷代謝提供充分的底物;4月28日以后濕度未達(dá)到100%,且10%及以下濕度出現(xiàn)幾率增加,5月2日至4日濕度波動(dòng)范圍較小,最高濕度在70%左右。
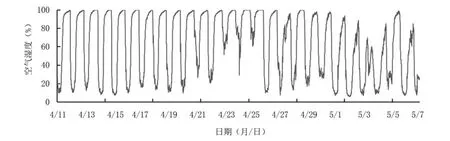
圖7 藍(lán)莓果實(shí)轉(zhuǎn)色期空氣濕度變化動(dòng)態(tài)
綜合氣象數(shù)據(jù)可見,棚內(nèi)前期光照較弱、平均氣溫較高且溫差較小、空氣濕度較大,適宜果實(shí)營養(yǎng)物質(zhì)的積累,以便為花色苷合成提供充分的底物;后期光照相對較強(qiáng),平均溫度較低、溫差較大、濕度較小、光照較強(qiáng),加之前期的底物累積,更利于果實(shí)花色苷合成。
3 討論與結(jié)論
品種是影響花色苷含量的重要因素,直接決定花色苷基因型[24]。本試驗(yàn)結(jié)果表明,引種的南高叢藍(lán)莓品種奧尼爾和米斯蒂在膠東地區(qū)設(shè)施栽培時(shí)的花色苷積累規(guī)律不同,奧尼爾的花色苷合成呈穩(wěn)定增長模式,而米斯蒂的花色苷合成屬于激增式。奧尼爾花色苷代謝相關(guān)基因中,前端合成基因VcPAL、VcCHS、VcCHI及后端合成基因VcANS、VcUFGT的相對表達(dá)量在果實(shí)轉(zhuǎn)色前期至后期逐步升高,且在轉(zhuǎn)色后期至轉(zhuǎn)色完成期的基因表達(dá)量增幅較高,合成中端基因VcF3H和VcDFR表達(dá)量先升高再降低,VcGLH、VcPPO表達(dá)量先降低再升高,這與花色苷在轉(zhuǎn)色中期開始大量累積趨勢一致。米斯蒂花色苷合成相關(guān)基因表達(dá)量在轉(zhuǎn)色前期和花盤轉(zhuǎn)色期均較為平穩(wěn),轉(zhuǎn)色中期至轉(zhuǎn)色后期表達(dá)量激增,之后又?jǐn)嘌率较碌@與花色苷含量在轉(zhuǎn)色后期大量積累的規(guī)律一致。兩個(gè)藍(lán)莓品種花色苷代謝相關(guān)基因表達(dá)量的差異可能是由品種差異造成的,也可能是由于不同品種的基因表達(dá)對外界因素如溫度、濕度及光照等的響應(yīng)不同[25]。通過對藍(lán)莓生長環(huán)境的氣溫、濕度及光照強(qiáng)度分析可見,花色苷合成的不同階段其外界條件不同,前期棚內(nèi)日平均氣溫較高、溫差較小、濕度較大、光照強(qiáng)度相對較低,可能更適合奧尼爾花色苷相關(guān)基因的表達(dá);而中后期,棚內(nèi)日平均溫度下降、溫差較大、濕度下降、光照強(qiáng)度增大,可能更適合米斯蒂花色苷相關(guān)基因的表達(dá)。
綜合上述分析,米斯蒂的花色苷基因表達(dá)高峰期稍晚,集中表達(dá)特點(diǎn)更為顯著,但奧尼爾花色苷總量在整個(gè)轉(zhuǎn)色過程中均高于米斯蒂。造成這種差異的原因首先可能是品種間差異,其次可能是米斯蒂合成途徑中的VcANS表達(dá)量相對較低,限制了花色苷的合成[26]。
