文蛤防御素基因的克隆及其免疫表達(dá)特征研究
2022-11-10 08:58:38周燁
水產(chǎn)養(yǎng)殖 2022年10期
周燁
(通州區(qū)興仁鎮(zhèn)農(nóng)業(yè)農(nóng)村和社會(huì)事業(yè)局,江蘇 南通 226300)
防御素(Defensins)是一類陽(yáng)離子抗菌活性肽,廣泛存在于各種生物體中,是生物體先天免疫防御和適應(yīng)性免疫防御機(jī)制的重要組成部分[1-3]。通常防御素都是小分子肽,由12~50個(gè)氨基酸構(gòu)成,分子內(nèi)富含精氨酸和由半胱氨酸形成的分子內(nèi)二硫鍵[4]。最初在鱟(Tachypleus tridentatus)中發(fā)現(xiàn)了一個(gè)具有79 aa的防御素基因,被命名為Big Defensin(MmBD),作為抗菌肽家族(AMPs)的一員,該基因具有顯著的抗菌活性[5]。隨后在多個(gè)軟體動(dòng)物中開展了big Defensin基因的相關(guān)研究,包括菲律賓蛤仔(Ruditapes philippinesis)[6]、海灣扇貝(Argopecten irradians)[7]、美洲簾蛤(Mercenaria mercenaria)[8]、魁蚶(Scapharca broughtonii)[9]等。以上研究表明,big defensin基因在免疫應(yīng)答中發(fā)揮著重要作用。目前在關(guān)于文蛤(Meretrix meretrix)的研究中,還未見該基因的相關(guān)報(bào)道。
文蛤,屬軟體動(dòng)物門,瓣鰓綱,簾蛤目,簾蛤科,文蛤?qū)伲俏覈?guó)沿海灘涂重要的經(jīng)濟(jì)貝類之一。但由于近年來(lái)沿海重工業(yè)的發(fā)展和臨港船舶運(yùn)輸?shù)龋斐珊Q蟓h(huán)境污染,文蛤養(yǎng)殖病害不斷發(fā)生,由此造成巨大的經(jīng)濟(jì)損失,也嚴(yán)重影響了貝類養(yǎng)殖產(chǎn)業(yè)的可持續(xù)發(fā)展。目前認(rèn)為,重金屬污染是近年漁業(yè)環(huán)境污染的公害之一,是導(dǎo)致貝類死亡的直接原因。有關(guān)重金屬對(duì)文蛤抗氧化酶影響的研究已有一些報(bào)道,任虹等[10]研究了鐵(Fe)、銅(Cu)、鋅(Zn)和鉛(Pb)等重金屬離子對(duì)文蛤肝臟過(guò)氧化氫酶(CAT)和細(xì)胞色素氧化酶(CCO)2種酶活性的影響;田鎮(zhèn)等[11]研究了在銅離子(Cu2+)脅迫下,不同群體文蛤超氧化物歧化酶(SOD)和過(guò)氧化氫酶(CAT)活力的變化。但重金屬對(duì)文蛤防御系統(tǒng)分子機(jī)制的影響尚未見報(bào)道。現(xiàn)克隆了文蛤MmBD基因,分析其特征和組織分布規(guī)律,研究了在Cu2+脅迫下,該基因應(yīng)對(duì)環(huán)境壓力的免疫應(yīng)答變化,以期為文蛤免疫防御機(jī)制研究提供基礎(chǔ)資料。
1 材料與方法
1.1 試驗(yàn)材料
本試驗(yàn)所用文蛤來(lái)自江蘇省通州灣海區(qū)。選取4個(gè)健康成熟的文蛤個(gè)體,置于冰上麻醉,分別取其外套膜、鰓、肝胰腺、足、閉殼肌以及性腺等6個(gè)組織樣品。待文蛤性腺發(fā)育成熟后,進(jìn)行催產(chǎn),獲取4細(xì)胞期、囊胚期、原腸胚、D形幼蟲、殼頂幼蟲和稚貝6個(gè)不同胚胎發(fā)育時(shí)期樣品。設(shè)置對(duì)照組與試驗(yàn)組,開展Cu2+脅迫試驗(yàn)。使用CuSO4·5H2O(AR)制備1 g/L Cu2+母液,設(shè)置Cu2+質(zhì)量濃度為0.5 mg/L的海水為試驗(yàn)組,不加Cu2+的正常海水為對(duì)照組,每組30個(gè)文蛤個(gè)體,并設(shè)置3個(gè)平行試驗(yàn)。每個(gè)組分別于12、36和72 h取3個(gè)文蛤個(gè)體的鰓組織,均用液氮速凍,并置于-80℃冰箱中保存。
1.2 MmBD基因cDNA序列全長(zhǎng)克隆及序列分析
從轉(zhuǎn)錄組文庫(kù)中獲得MmBD的EST序列,采用cDNA末端快速擴(kuò)增技術(shù)(RACE)進(jìn)行基因cDNA序列全長(zhǎng)克隆,設(shè)計(jì)3'RACE和5'RACE特異性引物進(jìn)行擴(kuò)增(表1),將獲得的PCR產(chǎn)物進(jìn)行純化、回收,隨后連接到pMD-18T(TaKaRa)載體,37℃過(guò)夜培養(yǎng),轉(zhuǎn)化到大腸桿菌DH5α(TaKaRa)中克隆,將篩選的陽(yáng)性克隆送至生工生物有限公司測(cè)序,拼接獲得結(jié)果。使用ORF Finder、SMART、ExPASY、TMHMM等在線網(wǎng)頁(yè)和工具,開展序列結(jié)構(gòu)特征分析;使用MEGA 5.0構(gòu)建系統(tǒng)進(jìn)化樹。

表1 試驗(yàn)所用引物
1.3 熒光定量分析
根據(jù)RNA提取純化試劑盒和反轉(zhuǎn)錄試劑盒(天根)的說(shuō)明書,分別獲取各個(gè)組織的總RNA,測(cè)定OD260/OD280值、1.2%瓊脂糖凝膠電泳評(píng)估RNA濃度和質(zhì)量,再將總RNA反轉(zhuǎn)錄為cDNA,于-20℃保存。根據(jù)熒光定量預(yù)混試劑盒(天根)說(shuō)明書配置體系,同時(shí)設(shè)置ABI7300系統(tǒng)擴(kuò)增程序。每個(gè)cDNA樣品做3個(gè)平行,β-actin為內(nèi)參,擴(kuò)增產(chǎn)物的解離曲線分析保證擴(kuò)增質(zhì)量。熒光定量數(shù)據(jù)按照文獻(xiàn)[12]的2-ΔΔCt方法計(jì)算,試驗(yàn)數(shù)據(jù)均以平均值±標(biāo)準(zhǔn)差(Mean±SD)表示。使用SPSS 19.0軟件的單因素方差(One-Way ANOVA)對(duì)數(shù)據(jù)進(jìn)行分析,在單因素方差分析的基礎(chǔ)上,采用Duncan多重比較法進(jìn)行分析,P<0.05為結(jié)果差異顯著。
2 結(jié)果與分析
2.1 MmBD基因序列分析
采用RACE技術(shù)克隆獲得MmBD基因(Gen Bank:ON351576)的cDNA全長(zhǎng)序列,共758 bp,其中5'非翻譯區(qū)(UTR)長(zhǎng)181 bp,3'UTR長(zhǎng)214 bp,開放閱讀框(,ORF)長(zhǎng)363 bp,共編碼120氨基酸(圖1)。預(yù)測(cè)蛋白分子質(zhì)量為13.22 kD,理論等電點(diǎn)pI為7.55。MmBD的預(yù)測(cè)氨基酸序列包含1個(gè)信號(hào)肽(1~23 aa)、2個(gè)跨膜結(jié)構(gòu)域(7~24 aa,44~66 aa)、高度保守的Cys殘基C-X6-C-X3-C-X13(14)-C-X4-C-C和kex2樣蛋白酶(KEKR-AV)的裂解位點(diǎn)。預(yù)測(cè)蛋白前端信號(hào)肽區(qū)具有較強(qiáng)的疏水性,在成熟肽區(qū)均有疏水氨基酸和親水氨基酸分布(圖2)。預(yù)測(cè)三級(jí)結(jié)構(gòu)包含3個(gè)α螺旋和6個(gè)β折疊。氨基酸序列對(duì)比、系統(tǒng)進(jìn)化樹和系統(tǒng)進(jìn)化樹所用物種序列登錄號(hào)分別見圖3、4和表2。由圖3、4可見,文蛤MmBD基因與菲律賓蛤仔的親緣關(guān)系最近,先與菲律賓蛤仔聚為一支,再與牡蠣、貽貝聚為一大支,最后再與扇貝等軟體動(dòng)物聚為一大支。

圖1 文蛤Big Defesin基因cDNA全長(zhǎng)序列及其結(jié)構(gòu)特征
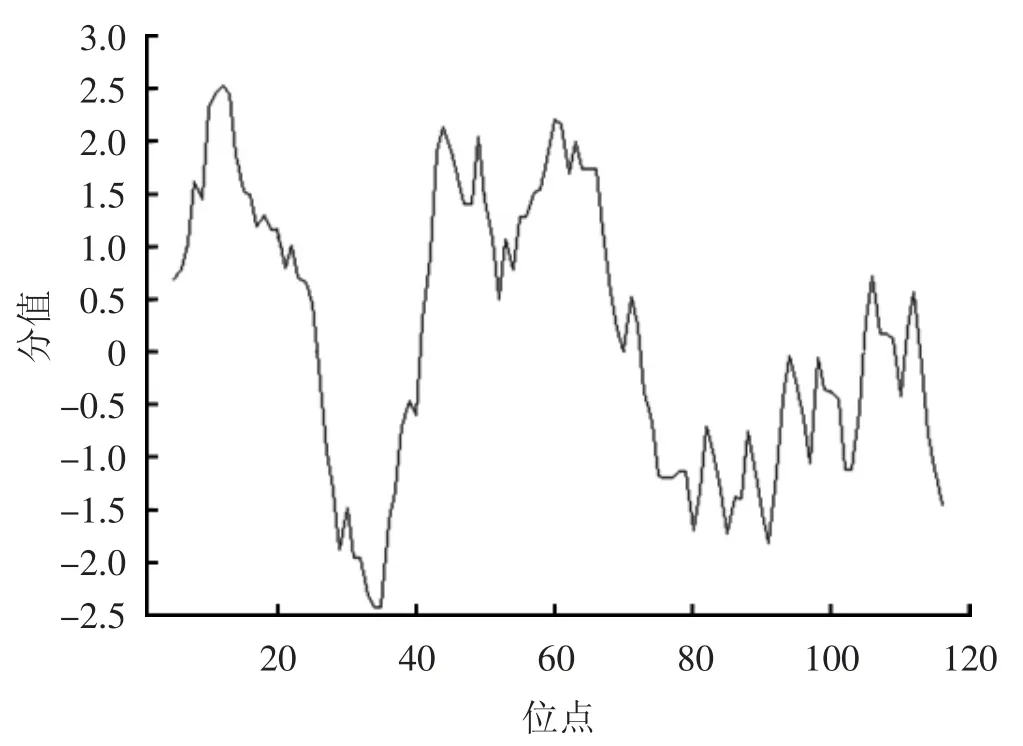
圖2 MmBD親水性與疏水性分析

圖3 文蛤與其他物種Big Defesin基因構(gòu)建的系統(tǒng)進(jìn)化樹

圖4 文蛤MmBD與其他軟體物種Big Defesin基因氨基酸序列對(duì)比
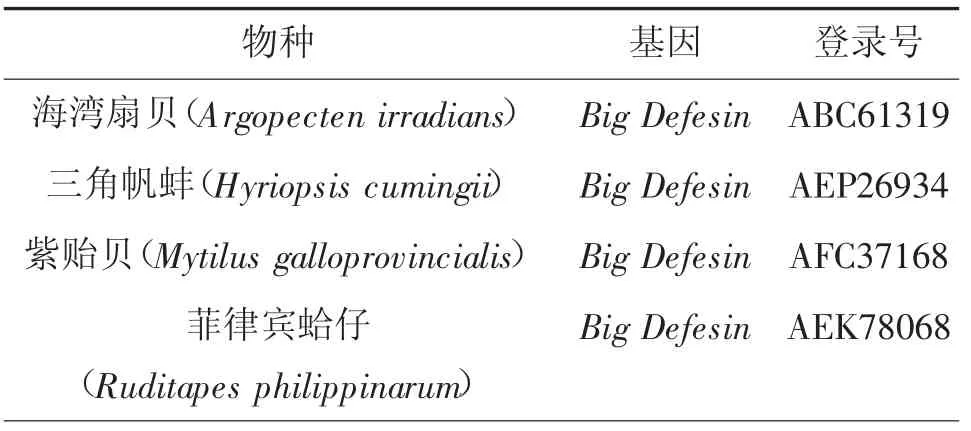
表2 系統(tǒng)進(jìn)化樹所用物種序列登錄號(hào)
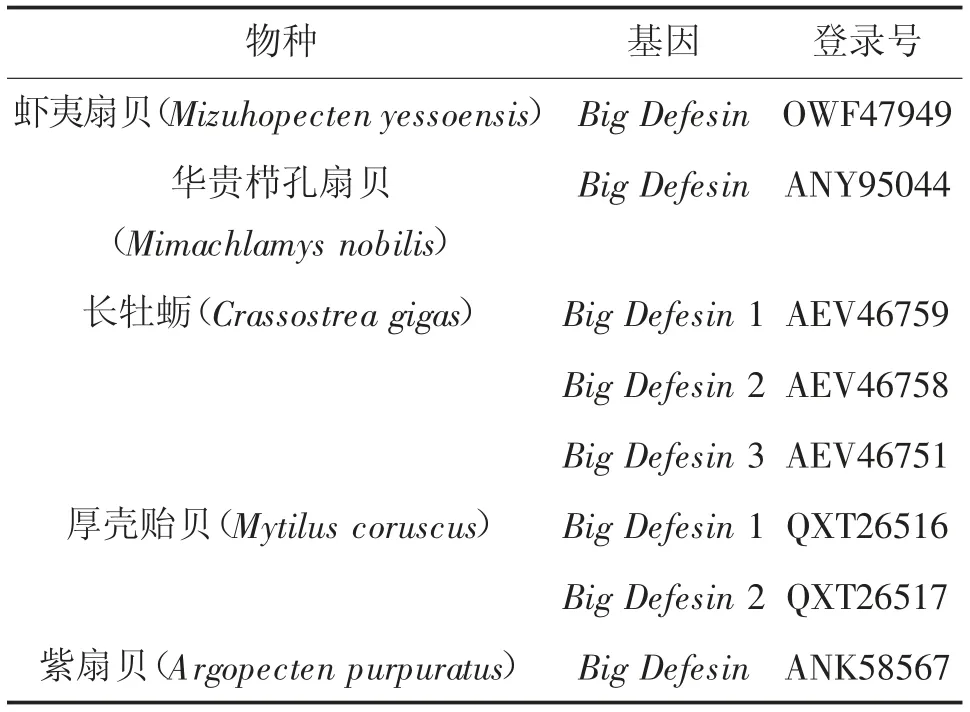
續(xù)表
2.2 MmBD基因的組織表達(dá)
使用qPCR(實(shí)時(shí)熒光定量核酸擴(kuò)增檢測(cè)系統(tǒng))分析MmBD基因在文蛤不同組織中(性腺、鰓、外套膜、肝胰腺、閉殼肌、足)的分布規(guī)律,結(jié)果見圖5。由圖5可見,MmBD在所檢測(cè)的各個(gè)組織中均有表達(dá),該基因在鰓組織中的表達(dá)量最高(P<0.05),其次是在外套膜和肝胰腺,而在閉殼肌、性腺和足中的表達(dá)量較低。
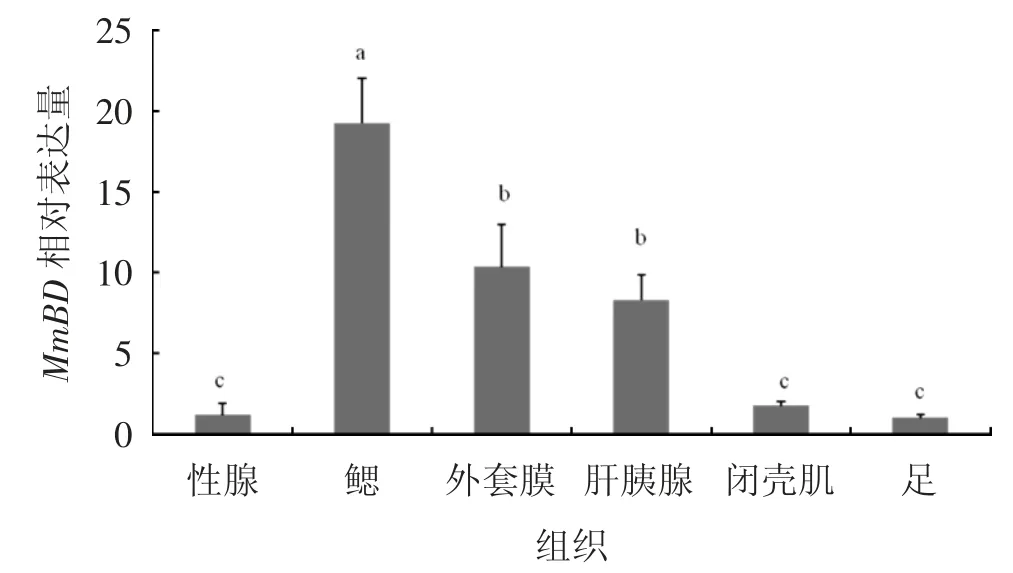
圖5 MmBD基因在文蛤各組織的分布情況
2.3 MmBD基因在胚胎發(fā)育時(shí)期的表達(dá)
MmBD基因在文蛤不同胚胎發(fā)育時(shí)期(4細(xì)胞期、囊胚期、原腸胚、D形幼蟲、殼頂幼蟲和稚貝)表達(dá)見圖6。由圖6可見,MmBD基因在D形幼蟲期表達(dá)量最高(P<0.05),其次是原腸胚期,稚貝期表達(dá)量最低(P<0.05)。
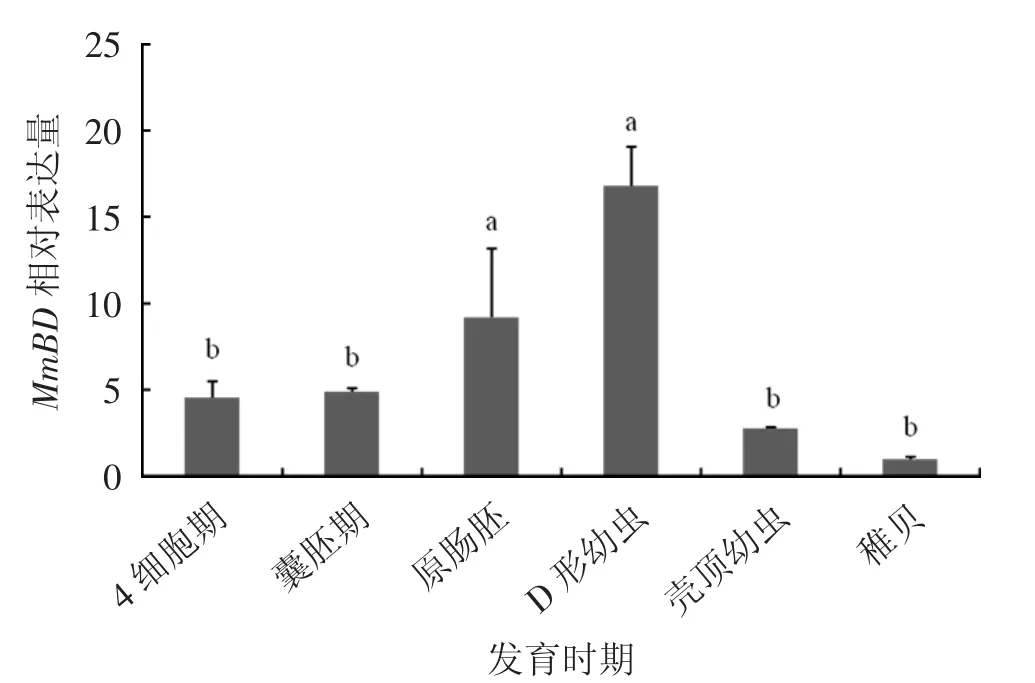
圖6 MmBD在不同胚胎發(fā)育時(shí)期的表達(dá)特征
2.4 Cu2+脅迫下MmBD基因的表達(dá)特征
高濃度Cu2+脅迫試驗(yàn)結(jié)果表明,對(duì)照組文蛤的MmBD基因表達(dá)量無(wú)顯著變化,試驗(yàn)組MmBD基因表達(dá)量明顯升高(圖7)。尤其是36和72 h,試驗(yàn)組MmBD基因表達(dá)量極顯著高于對(duì)照組(P<0.01)。

圖7 MmBD基因在Cu2+脅迫下的表達(dá)規(guī)律
3 討論
克隆獲得了文蛤Big Defensin基因的cDNA全長(zhǎng)序列,結(jié)果顯示,該基因的氨基酸序列N末端是一段具有較強(qiáng)疏水性的信號(hào)肽序列(23 aa),主要起到引導(dǎo)防御素前體蛋白通過(guò)脂膜的作用[13];C端包含1個(gè)高度保守的防御素結(jié)構(gòu),該結(jié)構(gòu)由6個(gè)特定位置的保守半胱氨酸殘基C-X6-C-X3-C-X13(14)-C-X4-C-C組成,這與魁蚶、紫貽貝、海灣扇貝等其他貝類的Big Defensin基因的結(jié)構(gòu)相似,都具有防御素基因家族的典型特征。由此推測(cè),文蛤MmBD基因可能發(fā)揮著與其他物種Big Defensin基因相似的功能。
MmBD在不同組織中的表達(dá)結(jié)果顯示,MmBD在性腺、腮等6個(gè)組織中均有表達(dá)。這與魁蚶[9]、文昌魚(Branchiostoma japonicum)[14]、雜色鮑(Halitotis diversicolor)[15]等的研究結(jié)果相似,Big Defensin基因在不同組織中廣泛表達(dá)。文蛤MmBD基因在鰓中的表達(dá)量最高(P<0.05),其次是外套膜和肝胰腺。在魁蚶中,Big Defensin基因在肝胰腺中表達(dá)量最高,隨后是鰓、性腺以及外套膜;在雜色鮑中,Big Defensin基因在肝胰腺中表達(dá)量最高,其次是外套膜、足和鰓。在雙殼貝類中,鰓組織的主要功能是過(guò)濾和呼吸作用,外套膜是暴露于外部環(huán)境最多的組織之一,是免疫防御的第一道防線;肝胰腺的主要功能是消化代謝,是一種重要的免疫組織,可合成免疫因子啟動(dòng)免疫反應(yīng);這幾個(gè)組織對(duì)調(diào)節(jié)貝類自身的免疫機(jī)制至關(guān)重要,可有效抵抗外來(lái)有害物質(zhì)的入侵[16-17]。由此可推測(cè),文蛤MmBD基因與其他物種的Big Defensin基因一樣,在鰓、外套膜和肝胰腺等組織器官中起到重要作用,在機(jī)體免疫中發(fā)揮重要功能。MmBD在不同胚胎發(fā)育時(shí)期的表達(dá)結(jié)果顯示,MmBD在原腸胚期表達(dá)量突然上升,D形幼蟲期表達(dá)量最高(P<0.05)。文蛤幼體在原腸胚后期整個(gè)胚胎形態(tài)逐漸發(fā)生變化,形成擔(dān)輪幼蟲,隨后再變態(tài)形成D形幼蟲,D形幼蟲開始從外界攝食[18],MmBD在在原腸胚期開始大量表達(dá),主要在D形幼蟲期起到重要作用,這可能是MmBD基因在這一過(guò)程中對(duì)幼體自身開展的自我保護(hù)機(jī)制。
防御素主要參與調(diào)節(jié)機(jī)體免疫系統(tǒng),具有多項(xiàng)免疫功能,是生物體防御系統(tǒng)的重要成分。多項(xiàng)研究報(bào)道,Big Defensin基因與軟體動(dòng)物免疫防御密切相關(guān)。文獻(xiàn)[7,19]研究表明,海灣扇貝和蛤仔(Venerupis philippinarum)面對(duì)細(xì)菌攻擊時(shí),兩者血細(xì)胞內(nèi)的Big Defensin基因表達(dá)量顯著上調(diào),顯示出該基因較強(qiáng)的抗菌活性;Li等[9]研究表明,魁蚶SbBDef1基因參與了自身的免疫應(yīng)答過(guò)程,細(xì)菌侵染后其血細(xì)胞和肝胰腺中的SbBDef1基因均出現(xiàn)大量表達(dá)。此外,三角帆蚌(Hyriopsis cumingii)[20]、櫛孔扇貝(Chlamys nobilis)[21]等研究中顯示,Big Defensin基因同樣在自身的機(jī)體免疫中起到重要作用。本試驗(yàn)結(jié)果與以往的研究結(jié)果相似,文蛤MmBD基因在重金屬Cu2+脅迫下出現(xiàn)明顯上調(diào),尤其是36和72 h時(shí),試驗(yàn)組MmBD基因表達(dá)量約為對(duì)照組表達(dá)量的5倍,2組差異極顯著(P<0.01)。此結(jié)果說(shuō)明,文蛤MmBD基因響應(yīng)Cu2+脅迫下的免疫應(yīng)答,該基因可能在文蛤抵御外界環(huán)境刺激時(shí)發(fā)揮一定功能,其免疫防御機(jī)制有待繼續(xù)深入研究。
