亞砷酸鈉對PC12細胞增殖、凋亡及Hippo-YAP通路的影響
2022-11-09 01:36:56劉曉紅王宏健王東艷潘艷伶潘際剛
中國藥理學通報 2022年11期
朱 衛,李 成,劉 宇,吳 松,彭 博,劉曉紅,王宏健,王東艷,楊 莉,潘艷伶,潘際剛
(貴州醫科大學 1.基礎醫學院生理學教研室、2.臨床醫學院中醫學教研室,貴州 貴陽 550025)
砷是一種廣泛分布于地殼中的非金屬元素,人群常通過飲用地下水、進食含有砷的食物、燃燒含有砷的燃煤等途徑被暴露于高砷環境中,砷污染是一個嚴重且普遍的全球公共衛生問題[1-2]。流行病學調查發現,砷可造成皮膚、肝、腎、神經等多種臟器毒性損傷,我國是世界上砷污染最為嚴重的國家之一[3-4]。因此,開展有關砷的毒性損傷機理的研究具有重要的現實意義。砷可以誘導神經系統氧化應激、引起線粒體功能紊亂、影響神經遞質的調節從而造成神經系統的損傷[4]。但其具體的分子機制未被闡明。Hippo-YAP信號通路的核心是由哺乳動物STE20樣蛋白激酶1/2(mammalian STE20-like protein kinase 1/2,MST1/2),大腫瘤抑制因子1/2(large tumor suppressor 1/2,LATS1/2)和Yes相關蛋白(Yes-associated protein,YAP)所構成的激酶級聯,該通路在調控細胞增殖、細胞凋亡、控制組織器官大小等方面具有重要作用,Hippo-YAP通路的異常則會引起細胞增殖和凋亡間的平衡失調,從而造成組織器官的過度生長及腫瘤的發生[5-6]。目前未見文獻報道Hippo-YAP信號通路在砷誘導神經細胞凋亡方面的作用機制研究。因此,本研究以PC12細胞為研究對象,通過觀察周期、凋亡相關指標的變化以及Hippo-YAP信號通路相關蛋白的表達水平,進一步探討亞砷酸鈉(NaAsO2)誘發神經細胞毒性損傷的機制。
1 材料與方法
1.1 材料
1.1.1細胞系 PC12細胞株(中科院昆明細胞庫)。
1.1.2試劑 胎牛血清,購自以色列BioInd公司(批號:10099141C);青霉素-鏈霉素,購自美國Gibco公司(批號:15140163);胰蛋白酶,購自美國Gibco公司(批號:25200056);高糖DMEM培養基,購自美國Gibco公司(批號:8120293);NaAsO2,購自美國Sigma公司(批號:S7400);MST1多克隆抗體,購自ProteintechGroup(批號:22245-1-AP);LATS1多克隆抗體,購自ProteintechGroup(批號:17049-1-AP);YAP1多克隆抗體,購自ProteintechGroup(批號:13584-1-AP);Tubulin多克隆抗體,購自Bioworld(批號:BS1699);BAX多克隆抗體,購自ProteintechGroup(批號:50599-2-1g);BCL-2多克隆抗體,購自ProteintechGroup(批號:12789-1-AP);p53多克隆抗體,購自ProteintechGroup(批號:10442-1-AP);Phospho-MST1單克隆抗體,購自Cell Signaling Technology(批號:49332S);Phospho-LATS1單克隆抗體,購自Cell Signaling Technology(批號:9157S);Phospho-YAP1單克隆抗體,購自Abcam(批號:ab76252);抗兔IgG-HRP二抗,購自北京中衫金橋(批號:ZB-2305);細胞周期檢測試劑盒,凱基生物(批號:KGA512);Annexin V-FITC/PI細胞凋亡檢測試劑盒,凱基生物(批號:KGA1012)。
1.1.3儀器 Model 310細胞恒溫CO2培養箱(美國Thermo 公司);細胞培養超凈臺(蘇州凈化);全波段多功能酶標儀(美國BioTek公司);FACSCalibur流式細胞儀(美國BD公司);全自動凝膠成像儀(美國SynGene)。
1.2 方法
1.2.1細胞培養 PC12細胞培養在高糖DMEM培養基(含10%FBS和1%青霉素-鏈霉素)中,置于37 ℃、5%CO2、飽和濕度的培養箱中培養,隔天換液,細胞生長至80%匯合度時進行傳代或凍存。
1.1.2細胞周期檢測 取對數期的細胞鋪于6孔板中(5×108L-1),培養過夜,以含20、40、60 μmol·L-1NaAsO2的高糖DMEM培養基分別處理細胞,同時以無NaAsO2的高糖DMEM培養細胞作為空白對照,24 h后以不含EDTA的胰酶消化細胞,1 528g離心5 min,PBS洗2次,置于4 ℃冰箱用70%乙醇固定30 min,1 528g離心5 min,PBS洗2 次,1 528g離心5 min,棄PBS,加入300 μL PI/RNase染色工作液,室溫避光60 min,加入樣品管中,用流式細胞儀進行檢測。
1.2.3流式細胞儀檢測PC12細胞凋亡 本實驗分組及部分收集處理細胞步驟同“1.2.2”,最后再加入500 μL PBS緩沖液得到細胞懸液,用移液槍輕輕的吹打混勻,再依次加入5 μL的Annexin V-FITC工作液,并輕輕振蕩混勻后再加入10 μL的PI工作液,混勻后避光靜置于冰上,反應20 min后,置于流式細胞儀中檢測。
1.2.4蛋白印跡檢測 藥物處理細胞后,以RIPA細胞裂解液(含1 mmol·L-1PMSF)裂解細胞,冰上裂解10 min,2×104g離心15 min,取上清進行BCA蛋白定量,計算并配制上樣緩沖液,100 ℃變性5 min,10% SDS-PAGE電泳分離蛋白,濕轉至PVDF膜,5%的脫脂牛奶封閉2 h,一抗(1 ∶1 000)或內參抗體Tubulin(1 ∶50 000)于4 ℃搖床孵育過夜,TBST洗膜3次,抗兔IgG-HRP(1 ∶20 000)二抗室溫孵育1 h,TBST洗膜3次,以化學發光試劑盒進行顯色,凝膠成像儀成像,用ImageJ對蛋白印跡條帶進行處理和分析。
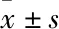
2 結果
2.1 NaAsO2對PC12細胞周期的影響通過流式細胞儀檢測不同濃度NaAsO2(20、40、60 μmol·L-1)處理24 h對PC12細胞周期的影響。如Tab 1、Fig 1所示,與對照組相比,20 μmol·L-1NaAsO2處理24 h后,PC12細胞中各期比例無明顯改變,差異無統計學意義,40 μmol·L-1NaAsO2處理24 h后,PC12細胞中G0/G1期比例明顯降低(P<0.01),而S期比例明顯增加(P<0.05),60 μmol·L-1NaAsO2明顯降低G0/G1(P<0.01)和G2/M期比例(P<0.05),而升高S期比例(P<0.01),而且,與20 μmol·L-1NaAsO2處理組相比,NaAsO2在40、60 μmol·L-1呈劑量依賴性的誘導G0/G1比例降低(P<0.05)、S期比例增加(P<0.01)。

Tab 1 Effect of NaAsO2 on cell cycle of PC12 cells n=3)
2.2 NaAsO2促進PC12細胞凋亡通過流式細胞儀檢測不同濃度NaAsO2(20、40、60 μmol·L-1)分別處理PC12細胞24 h后對細胞凋亡率的影響,結果如Fig 2所示。與對照組相比,20 μmol·L-1NaAsO2處理24 h后,PC12細胞凋亡率無明顯改變,差異無統計學意義,40、60 μmol·L-1的NaAsO2誘導PC12細胞凋亡率由3.58%分別上升至11.06%、18.20%(P<0.05),與20 μmol·L-1NaAsO2處理組相比,40、60 μmol·L-1的NaAsO2以劑量依賴性誘導PC12細胞凋亡率增加(P<0.05)。
為了進一步證實NaAsO2的促PC12細胞凋亡作用,我們通過蛋白印跡法檢測不同濃度NaAsO2(20、30、40、50、60 μmol·L-1)處理24 h后,對PC12細胞凋亡相關蛋白表達的影響。如Fig 3所示,與對照組相比,NaAsO2處理后,20、30 μmol·L-1NaAsO2處理后BCL-2蛋白的表達水平無明顯變化,當濃度增加到40、50、60 μmol·L-1時BCL-2蛋白的表達水平明顯下降(P<0.01),20、30、40 μmol·L-1NaAsO2處理后BAX的表達水平明顯升高(P<0.05),不同濃度(20、30、40、50 μmol·L-1)的NaAsO2處理后均可使p53蛋白的表達水平明顯上調(P<0.01)。
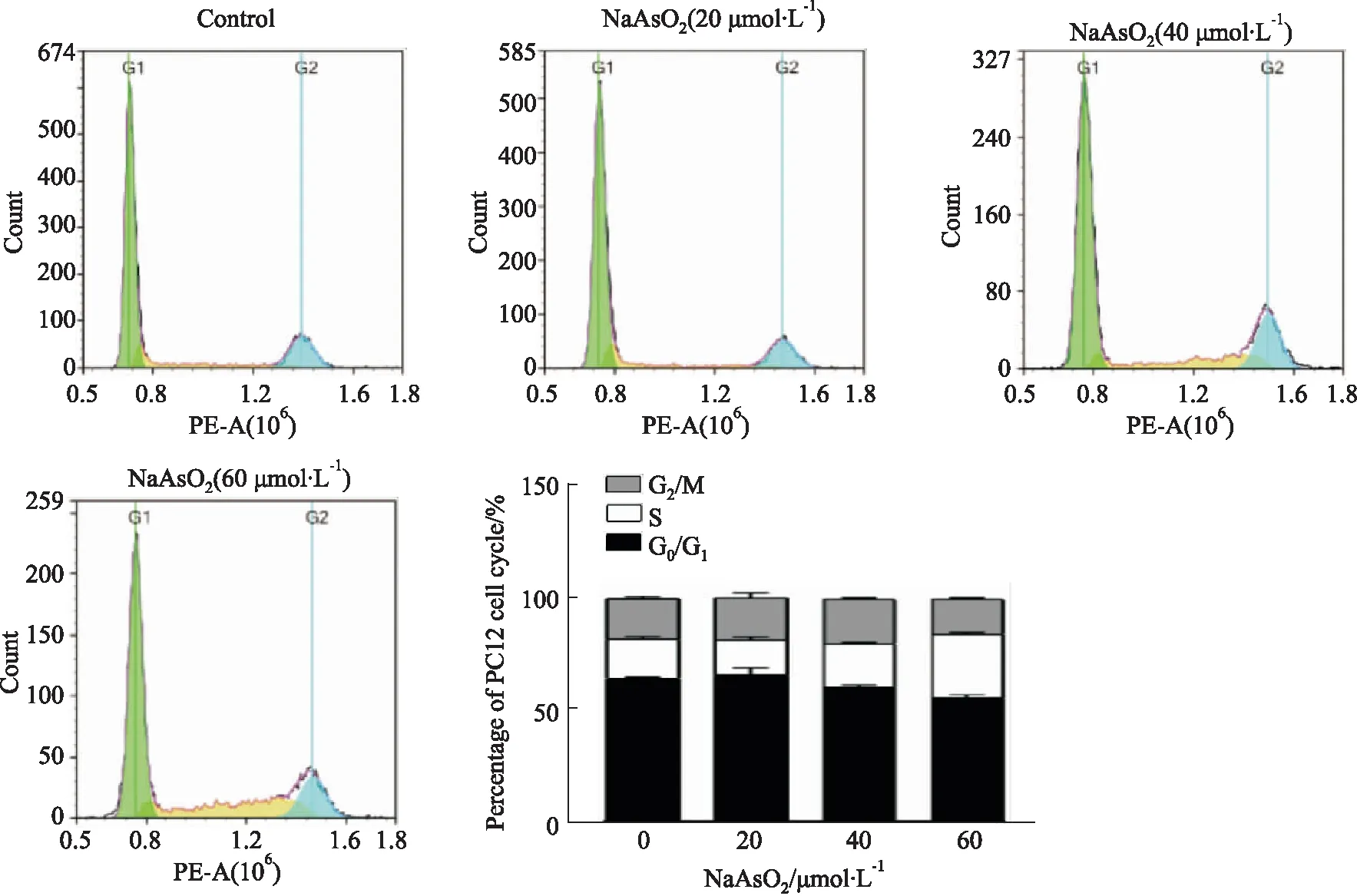
Fig 1 Effect of NaAsO2 on cell cycle of PC12 cells n=3)

Fig 2 Effect of NaAsO2 on apoptotic rates of PC12 cells n=3)

Fig 3 Effect of NaAsO2 on expression of apoptosis-related proteins in PC12 n=3)
2.3 NaAsO2對PC12細胞Hippo-YAP通路的影響為了探究NaAsO2對PC12細胞Hippo-YAP通路的影響,我們使用蛋白印跡檢測經過不同濃度NaAsO2(20、30、40、50、60 μmol·L-1)處理24 h后Hippo-YAP通路相關蛋白表達水平的變化。如Fig 4所示,與對照組比較,60 μmol·L-1NaAsO2處理后,MST1蛋白表達水平明顯下調(P<0.05),40、50、60 μmol·L-1NaAsO2處理后LATS1蛋白表達水平明顯下調(P<0.01)、50、60 μmol·L-1NaAsO2處理后YAP蛋白表達水平明顯下調(P<0.01),而p-MST1、p-LATS1、p-YAP蛋白表達水平在經不同濃度(20、30、40、50、60 μmol·L-1)的NaAsO2處理后均發生明顯上調(P<0.05)。
3 討論
越來越多的流行病學調查和臨床病例報告顯示,砷暴露會導致嚴重的神經系統功能障礙,包括神經系統的發育異常、學習記憶以及認知能力的損傷等[1-2]。本課題組的前期研究結果顯示,NaAsO2暴露后PC12細胞生長受到明顯抑制,可能與3MST蛋白表達下降有關[7]。在本研究中,以PC12細胞染砷,通過流式細胞儀檢測細胞周期以及細胞凋亡,結果顯示,NaAsO2可使PC12細胞發生S期阻滯,明顯誘導PC12細胞凋亡。細胞周期是細胞的基本生命活動,細胞的生長和復制依賴于完整的細胞周期進程,當細胞受到不良刺激時,會引起周期檢查點的異常進而使細胞周期滯留在周期檢查點上稱之為細胞周期阻滯。Zhao等[8]用二硫化二砷處理MDA-MB-231細胞發現S期細胞明顯增加。柳香香等[9]用NaAsO2處理SH-SY5Y細胞后也發現S期細胞比例增加。本實驗也發現NaAsO2處理后可使PC12細胞發生明顯的S期阻滯。
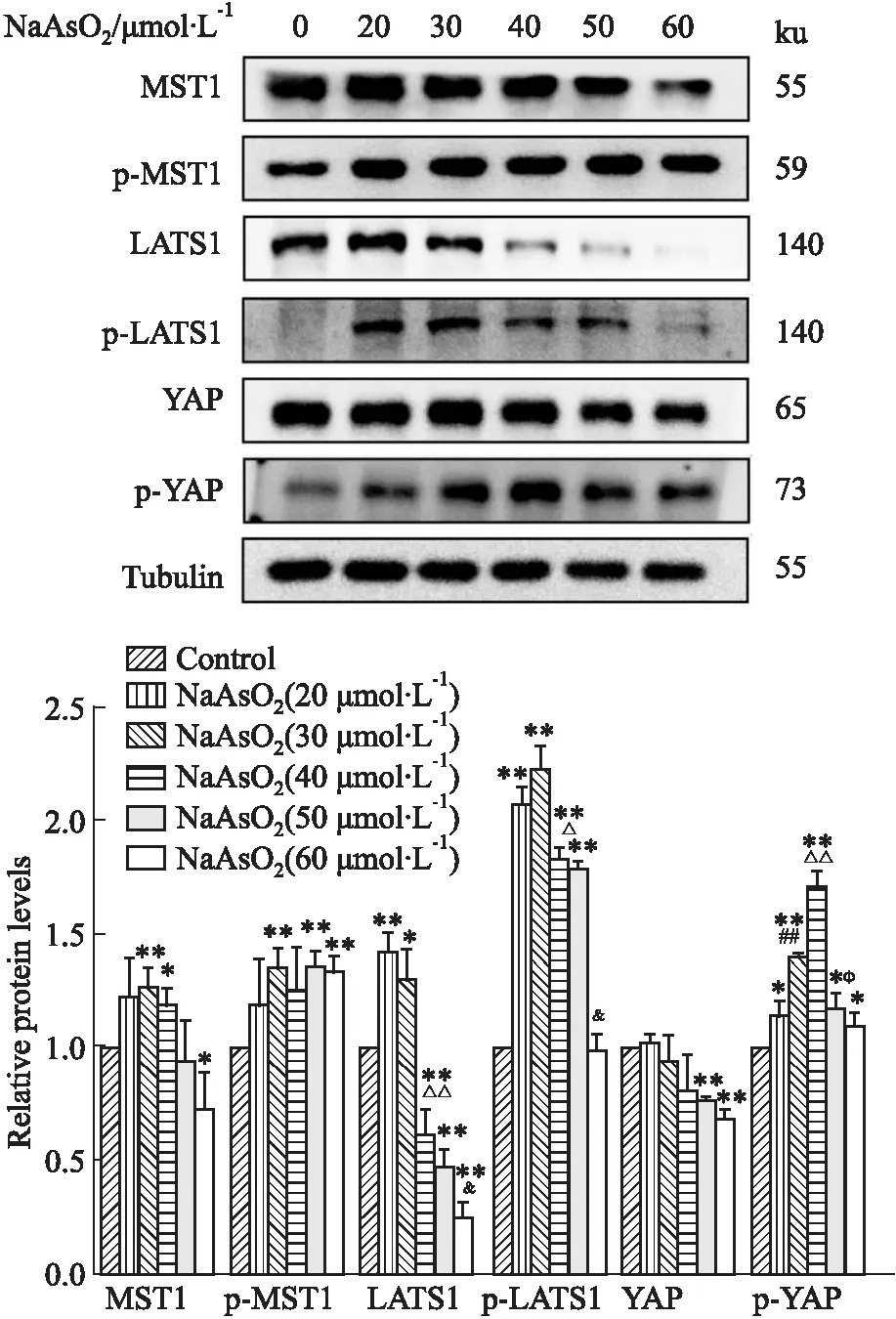
Fig 4 Effect of NaAsO2 on expression of proteins associated with Hippo-YAP signaling pathway in PC12 cells n=3)
BCL-2家族是凋亡通路中研究最為廣泛的,其中BCL-2是目前已知抗凋亡作用最強的因子之一,而以BAX為代表的一類分子則促進細胞凋亡的發生,BAX/BCL-2比例增加時促進細胞凋亡的發生,當二者比列下降時則抑制細胞凋亡的發生[10]。p53基因是一種腫瘤抑制基因,其編碼的蛋白稱為p53蛋白,Kircelli等[11]發現,As2O3以p53依賴方式誘導骨髓瘤細胞發生凋亡。Yang[12]在NaAsO2暴露處理的大鼠模型中研究發現,大鼠肝組織內p53蛋白的表達、BAX/BCL-2比例均升高。Sun等[13]研究發現,As2O3可明顯抑制神經膠質瘤細胞抗凋亡基因BCL-2的表達,并以時間依賴性方式上調凋亡基因BAX的表達,從而誘導細胞凋亡。本實驗研究中對上述BCL-2家族以及p53蛋白的檢測結果表明,經NaAsO2處理24 h后,出現了明顯的BCL-2下調,BAX和p53的表達上調,與文獻報道一致,提示NaAsO2誘導PC12凋亡可能是通過BCL-2家族蛋白實現的。
Hippo-YAP信號通路在各物種之間高度保守,其通過調控細胞增殖-凋亡平衡在維持組織穩態和器官形態上具有重要作用[5]。Wei等[14]的研究表明,維替泊芬通過抑制YAP蛋白的表達水平,引起BCL-2表達下調,從而誘導胰腺導管腺癌細胞PANC-1和SW1990凋亡。Song等[15]研究表明,子宮內膜異位癥的子宮內膜基質細胞(endometrial stromal cell,ESC)中的YAP敲低降低了細胞增殖并增強了細胞凋亡,同時降低了BCL-2的表達;而YAP的過表達導致正常ESC的增殖增加和凋亡減少,誘導BCL-2的表達增加,并證實BCL-2為YAP的直接下游靶基因,用維替泊芬(YAP抑制劑)處理子宮內膜異位癥的ESC顯示增殖減少和凋亡增強,并且在維替泊芬處理的裸鼠子宮內膜異位癥動物模型中,子宮內膜異位病變的大小明顯減小,提示Hippo-YAP信號通路參與子宮內膜異位癥的發病機制。Li等[16]研究表明,丙泊酚可以抑制YAP磷酸化和促進其核易位來激活YAP,激活的YAP增加了Bcl-2基因的轉錄并增加BCL-2蛋白表達,促進HT-22細胞存活。Li等[17]研究表明,阿霉素處理明顯下調H9c2細胞YAP的表達,引起BCL-2下調,以及BAX的表達上調,并誘導細胞凋亡,褪黑激素可以使YAP水平在阿霉素的處理下保持不變,并逆轉阿霉素誘導的H9c2細胞凋亡,而通過siRNA抑制YAP的表達時,褪黑激素失去了對阿霉素處理的心肌細胞的保護作用,提示YAP在其中起到至關重要的作用。Niu等[18]的研究指出,化合物9i通過激活p-LATS抑制YAP的活性下調BCL-2的表達,促進胃癌MGC-803和SGC-7901細胞凋亡,提示化合物9i可能通過激活Hippo-YAP信號通路誘導胃癌細胞凋亡。這些研究提示Hippo-YAP信號通路在調控細胞凋亡中扮演了至關重要的角色。在本實驗中,我們發現NaAsO2能夠上調PC12細胞內的p-MST1、p-LATS1、p-YAP蛋白表達水平從而激活Hippo-YAP信號通路,并且證實了NaAsO2可以誘導PC12細胞S期阻滯,通過上調p53、BAX的表達,抑制BCL-2的表達最終誘導PC12細胞凋亡。以上結果提示,NaAsO2誘導PC12細胞的凋亡作用可能是與Hippo-YAP信號的激活有關,Hippo-YAP通路激活從而抑制YAP的活性,導致抗凋亡蛋白BCL-2的表達下調,促凋亡蛋白BAX的表達上調,從而引起PC12細胞凋亡。然而,這些可能機制仍然有待進一步加以證實,從而有助于闡明砷中毒的分子機制,為臨床防治砷中毒提供一定的實驗依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
