腐爛莖線蟲不同群體致病性差異分析
2022-11-02 07:28:26倪春輝李惠霞劉永剛石明明魏雪娟李文豪徐雪芬
西北農(nóng)業(yè)學報 2022年8期
關(guān)鍵詞:差異
倪春輝,李惠霞,劉永剛,韓 變,石明明,魏雪娟,李文豪,徐雪芬
(1.甘肅農(nóng)業(yè)大學 植物保護學院/甘肅省農(nóng)作物病蟲害生物防治工程實驗室,蘭州 730070;2.甘肅省農(nóng)業(yè)科學院 植物保護研究所,蘭州 730070)
1945年,Thorne對引起馬鈴薯塊莖腐爛的線蟲進行了形態(tài)特征描述,并將其作為一個單獨的種從鱗球莖線蟲中分離出來,命名為腐爛莖線蟲(Ditylenchus destructor)[1]。該線蟲廣泛分布于溫帶地區(qū)[2-3]。在國外,該線蟲主要危害馬鈴薯[4]。在國內(nèi)主要危害甘薯[5]和當歸[6-8],也可危害馬鈴薯和黨參[9-10]。該線蟲可侵染90~120種植物,取食40個屬70多種真菌[11]。不同來源腐爛莖線蟲ITS-r DNA序列存在差異[3,12],國內(nèi)許多研究表明,不同來源腐爛莖線蟲ITS-r DNA序列存在A(L)型與B(S)型兩類[13-15]。2010年,Subbotin等[16]發(fā)現(xiàn)GenBank中70多個群體間的主要差異出現(xiàn)在ITS1區(qū)二級結(jié)構(gòu)H9螺旋區(qū)域,并將上述群體劃分為A-G 7個基因型。甘肅農(nóng)業(yè)大學線蟲實驗室發(fā)現(xiàn)來自當歸和黨參的腐爛莖線蟲群體與A-G基因型在二級結(jié)構(gòu)上存在差異,不能歸類為A-G基因型,可能存在新基因型(待發(fā)表)。
研究表明,不同來源腐爛莖線蟲群體致病性存在差異,在不同繁殖材料上的繁殖速率也不相同[17-21]。王宏寶等[21]將A、B基因型群體接種于甘薯,發(fā)現(xiàn)在感病品種上繁殖量最大的群體偏雌化。了解不同來源腐爛莖線蟲的致病性差異,有利于作物的準確選擇及輪作制度的制定。本研究擬測定不同群體的致病性,以期明確腐爛莖線蟲不同群體間的致病性差異,為該線蟲的防治提供有效理論依據(jù)。
1 材料與方法
1.1 供試材料
供試線蟲分離自馬鈴薯、當歸和黨參病樣,詳細信息見表1。

表1 供試腐爛莖線蟲群體信息Table 1 Population information of Ditylenchus destructor
供試植物馬鈴薯購自定西市農(nóng)科院‘新大坪品種’,甘薯購自河南省‘煙薯25號’品種,黨參為岷縣農(nóng)家品種。
1.2 盆栽種植
參照徐鵬剛[22]方法,采用16 cm×17 cm花盆種植黨參,18 cm×12 cm花盆種植馬鈴薯,25 cm×30 cm花盆種植甘薯。所用基質(zhì)為土壤∶腐殖質(zhì)∶沙子=2∶2∶1(體積比),經(jīng)高壓蒸汽121℃滅菌2 h后晾涼備用。將黨參苗和馬鈴薯塊莖于75%酒精中1 min,用無菌水沖洗3遍后浸入2.5% NaClO(有效氯含量)溶液中消毒10 min,最后再沖洗3遍;剪除甘薯苗根部(排除須根內(nèi)線蟲的影響),隨后浸沒在75%酒精10 s后,在2.5% NaCl O(有效氯含量)溶液中處理3 min,最后用無菌水沖洗3遍。將消毒后的種苗及塊莖種植于基質(zhì)中,澆水保濕。黨參、甘薯和馬鈴薯各種植21盆(1株/盆)。
1.3 線蟲接種
線蟲懸浮液制備:采用改良貝曼漏斗法分離在茄鐮孢培養(yǎng)的腐爛莖線蟲,經(jīng)200目和600目套篩沖洗真菌孢子后,將線蟲移至1.5 m L離心管,以3 000 r/min離心2 min沉淀線蟲且定容至1 m L。漩渦震蕩均勻,吸取100μL線蟲懸浮液,在顯微鏡下重復3次統(tǒng)計線蟲數(shù)量,計算線蟲懸浮液濃度。
線蟲接種:種苗種植30 d后,用移液槍將線蟲懸浮液接入植物根系附近,每盆接入大約500條各齡期混合線蟲。接種60 d后,觀察線蟲危害情況,并采用改良貝曼漏斗法[23]分離線蟲,統(tǒng)計線蟲數(shù)量:線蟲數(shù)(條/g)=分離線蟲總數(shù)/組織質(zhì)量;線蟲繁殖系數(shù)=最終線蟲總數(shù)/接種量(500條)。
1.4 莖線蟲病分級標準
甘薯莖線蟲病分級標準參考王宏寶[24]的方法:0級:無癥狀;1級:發(fā)病面積占甘薯塊根橫切面積的25%以下;2級:發(fā)病面積占甘薯塊根橫切面積的25%~50%(不含);3級:發(fā)病面積占甘薯塊根橫切面積的50%~75%(不含);4級:發(fā)病面積占甘薯塊根橫切面積的75%及以上。
馬鈴薯莖線蟲病分級標準參考林茂松[25]的方法:0級:無癥狀;1級:發(fā)病面積占馬鈴薯塊莖橫切面積的25%以下;2級:發(fā)病面積占馬鈴薯塊莖橫切面積的25%~50%(不含);3級:發(fā)病面積占馬鈴薯塊莖橫切面積的50%~75%(不含);4級:發(fā)病面積占馬鈴薯塊莖橫切面積的75%及以上。
黨參莖線蟲病分級標準參考劉學周[26]當歸麻口病分級標準:0級:無癥狀;1級:發(fā)病面積占黨參表面面積的1/2以下;2級:發(fā)病面積占黨參表面面積的1/2~2/3(不含);3級:發(fā)病面積占黨參頭部表面面積的2/3;4級:發(fā)病嚴重,已無完整頭部。
1.5 數(shù)據(jù)統(tǒng)計與分析
用軟件Excel 2016進行數(shù)據(jù)統(tǒng)計,使用軟件SPSS 19.0采用Duncan’s法對所有數(shù)據(jù)進行P<0.05水平的方差分析。
2 結(jié)果與分析
2.1 腐爛莖線蟲群體對寄主的致病性
接種腐爛莖線蟲60 d后,馬鈴薯、甘薯和黨參發(fā)病情況見表2。結(jié)果表明,在接種500條/盆的情況下,6個不同來源群體均可侵染馬鈴薯,4個群體可以侵染甘薯,但6個群體均未侵染黨參。馬鈴薯的C型群體DXP2、SXP1、HLJP1和黨參B型群體TCC1可以侵染馬鈴薯和甘薯。

表2 腐爛莖線蟲對不同作物侵染情況Table 2 Infection of D.destructor to different crops
2.2 腐爛莖線蟲群體對甘薯的致病性差異和癥狀特點
4個腐爛莖線蟲群體中馬鈴薯群體SXP1致病力顯著大于其他群體(表3,圖1-B),致病級數(shù)達3.67,線蟲數(shù)量也顯著多于其他群體,繁殖系數(shù)為30.43。其次為DXP2,致病級數(shù)為3.33,繁殖系數(shù)為3.50。HLJP1致病級數(shù)為2.33,繁殖系數(shù)為3.80,其致病力弱于DXP2,但繁殖系數(shù)略大于DXP2。黨參群體TCC1致病力和繁殖系數(shù)均低于其他群體,致病級數(shù)只有1級,且繁殖系數(shù)只有0.28,分離數(shù)量少于接種數(shù)量(500條/盆)。接種后各蟲態(tài)統(tǒng)計結(jié)果顯示,所有群體的幼蟲數(shù)量均最多,除群體TCC1外,其他群體雌雄比接近1∶1。C基因型3個群體中,HLJP1與其他群體致病級數(shù)具有顯著性差異,SXP1和DXP2與B基因型群體TCC1致病級數(shù)具有顯著性差異。
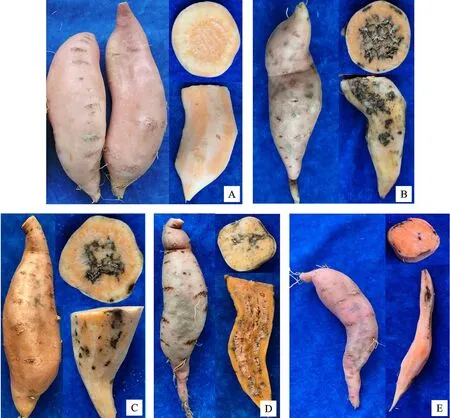
圖1 不同來源腐爛莖線蟲侵染甘薯癥狀Fig.1 Symptoms of sweet potato infected with D.destructor from different sources
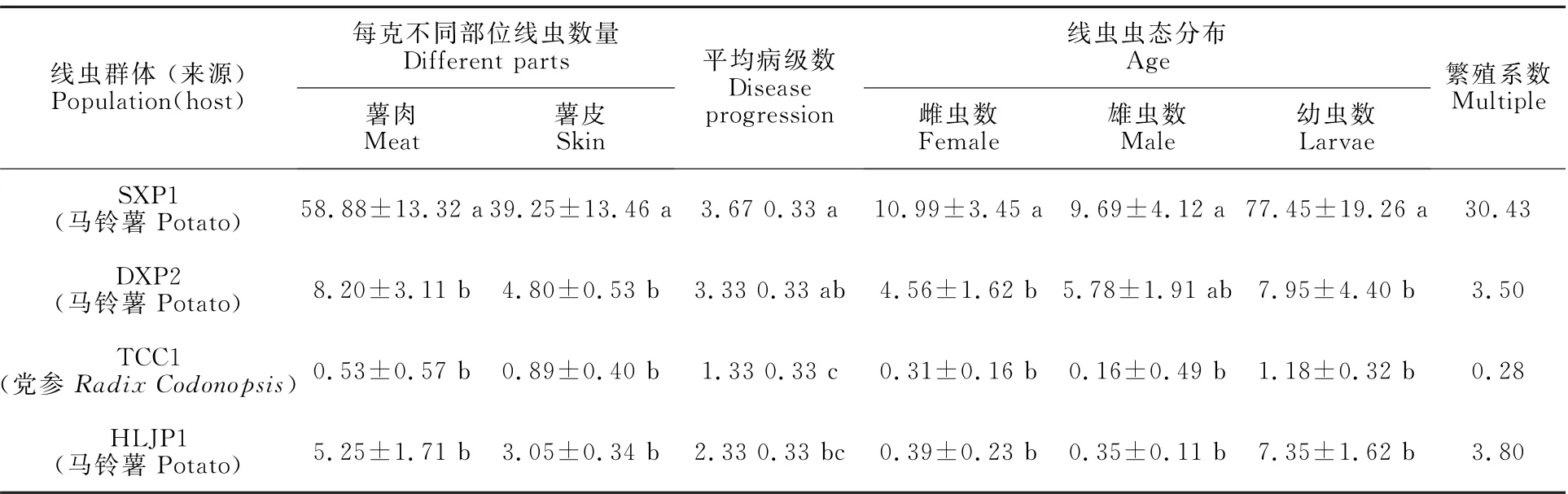
表3 腐爛莖線蟲不同群體對甘薯侵染情況Table 3 Infection of D.destructor from different sources to sweet potato
甘薯不同部位分離線蟲結(jié)果顯示,腐爛莖線蟲主要集中分布于塊根。采用削皮器將甘薯塊根削皮,分離薯皮和薯肉中的線蟲(表3),發(fā)現(xiàn)黨參群體TCC1在甘薯薯皮中的數(shù)量多于薯肉中,其他群體均為薯肉中數(shù)量較多。同時,莖、根和土壤中腐爛莖線蟲極少,大部分樣品未分離到腐爛莖線蟲,只有少數(shù)樣品分離得到1~5條線蟲。
發(fā)病癥狀觀察發(fā)現(xiàn),甘薯地上部、根系和莖基部未見明顯癥狀,塊根表皮發(fā)現(xiàn)有黑褐色斑點,發(fā)病嚴重時,薯塊表皮黑色斑點連接成片。削去表皮,可見黑色表皮下呈現(xiàn)黑褐色和白色相間的干腐癥狀,有些部位呈糠腐狀。切開薯塊,可見病斑由表皮向內(nèi)延伸,發(fā)病嚴重時,薯塊中心呈黑褐色和白色相間的干腐狀糠心。
2.3 腐爛莖線蟲群體對馬鈴薯的致病性差異分析和癥狀特點
供試6個群體對馬鈴薯致病力總體較弱。其中群體DXP2致病力最強(表4,圖2-D),致病級數(shù)為2.00,繁殖系數(shù)為2.50,但線蟲數(shù)量少于HLJP1。群體HLJP1繁殖數(shù)量最多,繁殖系數(shù)為4.11,顯著高于其他群體。群體DXP2和HLJP1繁殖數(shù)量大于接種數(shù)量,其他群體均為分離數(shù)量少于接種數(shù)量(500條/盆),且致病級數(shù)均小于2級,繁殖系數(shù)為0.002~0.59。統(tǒng)計結(jié)果顯示,所有群體均為幼蟲數(shù)量最多,雌雄比接近1∶1。C基因型群體SXP1、DXP2和HLJP1與B基因型群體TCC1致病級數(shù)無顯著性差異,與基因型L和H具有顯著差異。

圖2 不同來源腐爛莖線蟲侵染馬鈴薯癥狀Fig.2 Symptoms of potato infected with D.destructor from different sources
對馬鈴薯不同部位線蟲統(tǒng)計結(jié)果顯示,腐爛莖線蟲主要集中分布于塊莖。削皮后發(fā)現(xiàn)馬鈴薯薯皮和薯肉中線蟲數(shù)量均比較低,故未分別統(tǒng)計(表4)。其中,1個馬鈴薯樣品分離得到10條腐爛莖線蟲,大部分樣品未分離到線蟲。地上部和土壤中均未發(fā)現(xiàn)腐爛莖線蟲。

表4 腐爛莖線蟲不同群體對馬鈴薯侵染情況Table 4 Infection of D.destructor from different sources to potato
接種線蟲后,馬鈴薯地上部、根系和莖基部未見明顯癥狀。塊莖表皮有黑褐色斑點,表皮薄紙化易剝離,發(fā)病部位薯皮出現(xiàn)裂口。將薯塊橫切,可見病斑由表皮向內(nèi)延伸,呈現(xiàn)黑色點狀病斑,嚴重時為黑褐色干腐狀,薯塊中心無明顯癥狀。
2.4 腐爛莖線蟲對馬鈴薯和甘薯的致病性比較
接種60 d后,對每個處理的線蟲數(shù)量對比發(fā)現(xiàn)(圖3),SXP1、DXP2、HLJP1和TCC1等4個群體對甘薯和馬鈴薯均具有致病力,HLJP1在馬鈴薯上的繁殖數(shù)量大于甘薯,而其他群體在甘薯上的繁殖數(shù)量均高于馬鈴薯。甘薯上4個群體繁殖數(shù)量依次為SXP1>HLJP1>DXP2>TCC1;馬鈴薯上6個群體繁殖數(shù)量依次為HLJP1>DXP2>SXP1>W(wǎng)YA3>TCC1>HZA3。結(jié)合致病力結(jié)果(表2,表3)發(fā)現(xiàn),線蟲繁殖系數(shù)與其致病力大小總體呈正相關(guān),即繁殖系數(shù)越大致病力越強。HLJP1在甘薯上繁殖數(shù)略大于DXP2,但致病力DXP2大于HLJP1。
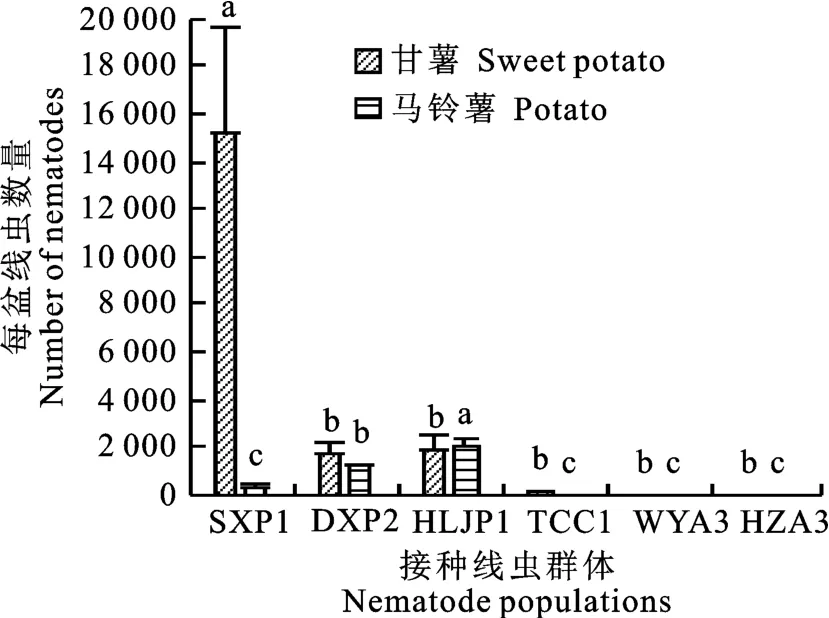
圖3 不同來源莖線蟲在甘薯和馬鈴薯繁殖數(shù)量上的對比Fig.3 Comparison of number of D.destructor isolated from different sources in sweet potato and potato
3 結(jié)論與討論
本研究對不同腐爛莖線蟲群體進行致病性測定。結(jié)果表明,3個不同地理來源的馬鈴薯C基因型群體對甘薯和馬鈴薯均具有較強致病性;黨參B基因型群體對甘薯致病力較弱,對馬鈴薯致病力極低;當歸L、H基因型群也可侵染馬鈴薯,但致病力較低,所有群體均未能侵染黨參。表明不同基因型、寄主來源和地理來源腐爛莖線蟲致病性均存在差異。
關(guān)于不同來源腐爛莖線蟲致病性差異的研究,主要集中于不同地理來源和基因型群體致病性差異分析。王宏寶[24]發(fā)現(xiàn)不同地域的10個甘薯腐爛莖線蟲群體對于甘薯塊根(‘蘇薯9號’‘寧薯9號’)的致病性,可以劃分為3個不同強度的致病類群。程云等[27]發(fā)現(xiàn)甘薯線蟲群體對于不同品種的花生致病力存在差異,證實在中國甘薯上的腐爛莖線蟲可以侵染花生。王宏寶等[21]發(fā)現(xiàn)不同基因類型(A、B型)群體在甘薯不同品種上雌雄比和繁殖數(shù)量存在差異。Macguidwin[28]發(fā)現(xiàn)腐爛莖線蟲對蠶豆致病性較強,紅三葉和玉米致病性次之,對苜蓿致病性較差,對洋蔥無侵染性。章淑玲[11]將來源于甘薯的腐爛莖線蟲接種于15種植物,發(fā)現(xiàn)腐爛莖線蟲可以侵染甘薯、白蘿卜、胡蘿卜、馬鈴薯、大蒜、花生和雍菜,且對甘薯的致病性強于馬鈴薯。本研究發(fā)現(xiàn)不同寄主、地理和基因型來源的6個腐爛莖線蟲群體,對甘薯和馬鈴薯的致病性存在差異,甘薯為腐爛莖線蟲良好寄主,馬鈴薯為中等寄主,此結(jié)果與其他相關(guān)報道相近。
Ni等[10]首次發(fā)現(xiàn)腐爛莖線蟲可以侵染黨參,田間調(diào)查發(fā)現(xiàn)該病發(fā)生較輕,盆栽接種5 000條/盆時,該線蟲可以侵染黨參。本研究中接種500條/盆時,所有群體均未侵染黨參,推測可能和黨參品種、接種數(shù)量有關(guān),或者黨參不是腐爛莖線蟲的良好寄主。
腐爛莖線蟲種內(nèi)分化嚴重,其致病性[11]、生物學特性[21]和抗藥性[29]等方面均存在差異。本研究發(fā)現(xiàn),來源于黨參、當歸和馬鈴薯的4類基因型腐爛莖線蟲群體致病性存在分化,對甘薯具有較強致病性,對馬鈴薯和黨參侵染力較低,由于條件限制,尚未測定對當歸的致病性。甘肅省是馬鈴薯和中藥材種植大省,每年有大量馬鈴薯種薯和藥材種苗銷往各地,這將有利于腐爛莖線蟲的傳播蔓延,故應(yīng)該加強腐爛莖線蟲的檢疫措施,降低該線蟲傳播和危害的風險。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
