干旱脅迫對東北紅豆杉野生種與栽培種生理與次生代謝的影響
2022-11-02 07:28:28王丹丹李曉紅張彥文劉丹梅
西北農(nóng)業(yè)學報 2022年8期
王丹丹,李曉紅,張彥文,劉丹梅
(遼東學院 農(nóng)學院,遼寧丹東 118000)
東北紅豆杉(Taxus cuspidataSieb.et Zucc.)又名紫杉、赤柏松,隸屬于紅豆杉科(Taxaceae)紅豆杉屬(Taxus)植物,屬于國家Ⅰ級珍稀保護植物,耐寒能力極強,喜蔭、冷濕肥沃排水性好的環(huán)境,林下喬木或灌木,為第三紀孑遺植物,被稱為 植物“活化石”。雌雄異株,風媒授粉;原產(chǎn)于中國東北長白山脈、朝鮮半島、日本等地。但由于東北紅豆杉是東北地區(qū)唯一的極小種群,生境狹小且分布地不連接,種群競爭力差;其樹皮、枝葉含抗癌物質紫杉醇,對治療乳腺癌、皮膚癌、卵巢癌和肺癌具有特殊的療效,木材可加工高質量家具或工藝品,實生苗木具有較高的觀賞價值,因而導致野生紅豆杉資源被過度砍伐而生境惡化,種群萎縮、遺傳多樣性降低,現(xiàn)已處于瀕危狀態(tài)[1]。因此,加大東北紅豆杉人工馴化栽培種的研究具有十分重要的意義。
干旱抑制植物生長發(fā)育,如抑制細胞有絲分裂,降低細胞膨壓,減弱細胞伸長及擴張能力,導致植株生長速度慢、植株矮細小、抽芽量少、葉面積減小、葉片枯黃皺縮以及根系發(fā)育不良等。可見,干旱脅迫造成植物不可逆的生理損傷[2]。根系是吸收水分的主要器官,干旱脅迫下,根系首先接收干旱信號,通過根系肉質化、變厚根皮、加強側根數(shù)量及向下深扎程度及提高根冠比來廣泛吸收土壤深層水分等方式響應干旱脅迫,之后信號傳遞到其他器官。此外,干旱脅迫會影響植物的滲透調(diào)節(jié)、營養(yǎng)代謝、呼吸作用與光合作用等,尤其是葉片形態(tài)特征及生理特性,如細胞壁變厚、葉片皺縮變小、氣孔下陷以及柵欄組織與海綿組織比值降低等;生殖階段干旱脅迫影響種子數(shù)量、大小和組成,而且花數(shù)、結莢率、結實率等指標均顯著下降[3]。此外,植物體內(nèi)很多基因直接參與對干旱脅迫的抵御,通過該類基因的表達或調(diào)控來減緩脅迫傷害,進而維持植物的正常生理代謝,主要包括ABRE結合蛋白/ABRE結合因子(AREB/ABFs)、脫水反應元件結合蛋白(DREB)、AP2轉錄因子、WRKY轉錄因子、N-乙酰-L-半胱氨酸(NAC)和堿性亮氨酸拉鏈(bZIP);信號轉導相關蛋白,如鈣依賴性蛋白激酶(CDPKs);抗氧化酶,如超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX))、抗壞血酸-谷胱甘肽(As A-GSH)循環(huán)系統(tǒng)中的酶,如還原型谷胱甘肽酶(GSH)、抗壞血酸酶(As A)、谷胱甘肽還原酶(GR)等;主要影響能量代謝過程中一些關鍵酶,如葡萄糖激酶(Glucokinase,GCK)、磷酸果糖激酶(Phosphofructokinase,PFK)、細胞色素c(Cytochrome c,Cyt c)、丙酮酸激酶(Pyruvate kinase,PK)、乳酸脫氫酶(Lactate dehydrogenase,LDH)與檸檬酸合酶(Citroyl synthetase,CS)等[4]均在干旱脅迫中發(fā)揮至關重要的作用。
由于東北紅豆杉生長緩慢,種子單生且發(fā)芽率極低,繁殖周期長(種子發(fā)芽需2 a,人工根插繁育需2~3個月,第一年生長較慢),而二萜類物質紫杉醇等的需求量日益增加,嚴重地制約著東北紅豆杉的工業(yè)化生產(chǎn),所以從野生東北紅豆杉培育栽培種是必然趨勢。目前,對東北紅豆杉的研究主要集中于紫杉醇生物合成途徑及其栽培技術,而對東北紅豆杉抗旱代謝物的動態(tài)變化研究較少,如氨基酸類、糖類化合物等,且對其干旱機制及抗旱代謝物與相關基因的研究相對薄弱[5]。此外,干旱脅迫對東北紅豆杉生長發(fā)育的影響遠遠超出其他脅迫,并在發(fā)育完成后繼續(xù)影響其營養(yǎng)運輸、吸收、光合作用與呼吸作用等生理代謝過程;從野生東北紅豆杉培育栽培種顯著提高了植株性狀,但對逆境脅迫下,野生種與栽培種的抗逆性及次生代謝物差異的研究尚未見報道。綜上所述,本研究則對干旱脅迫下東北紅豆杉野生種與栽培品種的植株生理指標(葉片相對含水量、可溶性糖、可溶性蛋白、葉綠素、類胡蘿卜素以及光合參數(shù))、體內(nèi)次生代謝物含量(LDH、GCK、Cyt c、精氨酸、鳥氨酸轉氨酶、脯氨酸,丙二醛、天冬氨酸)以及紫杉烷-C13-側鏈-N-苯甲酰轉移酶(Tc-DBTNBT),轉錄因子AP2,脫水反應元件結合蛋白(DREB)以及光合作用光系統(tǒng)Ⅰ亞基X蛋白(Psa K)的表達量差異進行分析,為深入開展野生東北紅豆杉種質資源的保護及栽培種的培育與開發(fā)利用提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
選取10 a生健康無病且生長勢一致的野生東北紅豆杉與栽培種‘丹紫1號’(品種權號:20210592)為試驗材料(材料取自丹東科技生態(tài)園),采用盆栽控水法,將紅豆杉裸株栽培于直徑60 cm×60 cm(直徑×高)盆缽中,土壤為沙熟土∶腐葉土∶菌根菌生物有機肥∶粗砂∶泥炭=1∶1∶1∶1∶1,一缽一株紅豆杉,生境和栽培管理措施一致,置于露地養(yǎng)護。干旱處理前將植株澆透水,后將野生種與栽培種分為兩組,分別在處理后的第0、8、16、24、32、40、48天采集紅豆杉中下部外圍針葉檢測生理指標、次生代謝物及基因表達分析[6],各組每個時間段采用2盆紅豆杉植株為1次重復,每個試驗均做3次生物學重復。
1.2 方法
1.2.1 生理指標檢測 采用烘干稱重法測定紅豆杉葉片相對含水量[7]。可溶性糖和可溶性蛋白檢測分別采用可溶性糖試劑盒與可溶性蛋白試劑盒(生工生物工程(上海)股份有限公司,中國);每天9:00采用便攜式光合熒光測量系統(tǒng)(LI-6800,LI-COR公司,美國)內(nèi)置光源[光照強度1 500 μmol/(m2·s)]對紅豆杉中下部外圍針葉(每個處理采用相同部位葉片)測定光合速率(Pn),蒸騰速率(Tr)、氣孔導度(Gs)、胞間CO2濃度(Ci)值[8],每個處理重復3次取平均值。
1.2.2 次生代謝產(chǎn)物檢測 采用ELISA試劑盒(生工生物工程(上海)股份有限公司上海,中國)說明書操作并檢測干旱脅迫下東北紅豆杉野生種與栽培種相關酶(LDH、GCK和Cyt c)的酶活性以及精氨酸(Arg)含量[9];采用GSH檢測試劑盒和As A檢測試劑盒(北京索萊寶科技有限公司,中國)測定As A-GSH循環(huán)系統(tǒng)酶活性[10]。
采用葉綠素試劑盒和胡蘿卜素試劑盒測定色素含量(合肥萊爾生物科技有限公司,中國);采用磺基水楊酸法提取、測定紅豆杉針葉脯氨酸(Pro)的含量;采用硫代巴比妥酸(TAB)法提取、測定紅豆杉針葉丙二醛(MDA)含量[11];采用鳥氨酸轉氨酶試劑盒(江蘇連云港生物科技有限責任公司)提取鳥氨酸轉氨酶的含量;采用富馬酸法提取、測定天冬氨酸(Asp)的含量[12]。
1.2.3 東北紅豆杉野生種Tc DBTNBT基因RACE克隆 以曼地亞紅豆杉(Taxus×media)DBTNBT基因序列(GenBank:AY563629.1)設計引物(表1)。按RNA提取試劑盒(EZ-10 Total RNA Mini-Preps Kit)說明書提取總RNA,以總RNA為模板,按AMV第一鏈c DNA合成試劑盒(AMV First Strand cDNA Synthesis Kit)合成紅豆杉cDNA第一鏈,-20℃保存,用于克隆DBTNBT核心序列,反應體系20μL,包含0.8 μL cDNA,1μL d NTP Mix,0.6μL 10μmol/L上下游引物,(10×)AdvantagePCR緩沖液2.0μL,(50×)Advantage聚合酶混合液0.5 μL,加dd H2O至20μL;反應程序:95℃預變性4 min;94℃變性45 s,55~58℃退火50 s,72℃延伸60 s,30個循環(huán);72℃延伸10 min;回收PCR產(chǎn)物,將篩選的陽性克隆送至生工生物工程(上海)公司測序[13]。采用RACE試劑盒(SMARTer RACE cDNA Amlification Kit試劑盒)快速擴增、拼接Tc DBTNBT基因,測序驗證[14]。共克隆6個東北紅豆杉干旱脅迫相關基因,以Tc-DBTNBT克隆為例。
1.2.4 表達分析 選擇東北紅豆杉干旱應答元件結合蛋白DREB與光合作用光系統(tǒng)Ⅰ亞基X蛋白(Psa K),以及紫杉醇生物合成相轉錄因子AP2和關鍵酶Tc DBTNBT進行實時熒光定量PCR(qRT-PCR),檢測上述蛋白于干旱脅迫下的表達情況(引物見表1)。qRT-PCR反應體系(25 μL),SYBR Premix ExTaqⅡ(2×)5.0μL,10 μmol/L上下游引物各1.5μL,ROX參比染料(50×)0.8μL,cDNA模板2.0μL,加dd H2O至25μL;反應程序:95℃預變性2 min;95℃變性50 s,56~60℃退火45 s,72℃延伸50 s,40個循環(huán);每個樣品3次生物學重復取平均值,采用2-△△Ct法進行上述蛋白表達水平的分析計算[15]。

表1 引物信息Table 1 Information of primers
1.2.5 數(shù)據(jù)統(tǒng)計與分析 采用Excel 2016和DPS 7.05軟件進行試驗數(shù)據(jù)與差異顯著性分析,采用SPASS 21.0與SAS/STAT進行方差分析,采用Duncan氏多重極差檢驗(P<0.05)。所有試驗均采用完全隨機設計,生物學重復3次,取平均值。
2 結果與分析
2.1 干旱脅迫對東北紅豆杉野生種與栽培種葉片相對含水量的影響
葉片是對干旱脅迫應答最敏感的器官之一(圖1),干旱脅迫下,紅豆杉葉片相對含水量由干旱脅迫開始逐漸呈下降趨勢,其中野生東北紅豆杉葉片相對含水量始終低于栽培種,在干旱脅迫的第24天~40天,下降趨勢顯著,至第48天時,野生種葉片相對含水量不足20%,而栽培種葉片相對含水量為30%,是野生種的1.58倍。

圖1 干旱脅迫下東北紅豆杉野生種與栽培種葉片相對含水量Fig.1 Relative water content in leaves of wild and cultivated species of Taxus cuspidata under drought stress
2.2 干旱脅迫對東北紅豆杉野生種與栽培種葉片可溶性糖和可溶性蛋白含量的影響
可溶性糖與可溶性蛋白屬于滲透調(diào)節(jié)物質,在持續(xù)干旱脅迫下,紅豆杉可通過提高二者含量來調(diào)控自身含水量,達到維持植株正常生長發(fā)育的目的。干旱脅迫促使東北紅豆杉野生種與栽培種葉片可溶性糖含量增加,曲線趨勢基本一致(圖2-A),紅豆杉栽培種可溶性糖含量顯著高于野生種,干旱脅迫初期二者差異不顯著,第24天栽培種可溶性糖含量顯著增加,至第32天栽培種葉片可溶性糖含量已為野生種的1.28倍。干旱脅迫下,東北紅豆杉野生種與栽培種葉片可溶性蛋白含量在干旱脅迫初期呈現(xiàn)明顯的上升趨勢,而后逐漸下降(圖2-B),干旱處理前及脅迫初期野生種可溶性蛋白含量明顯高于栽培種,隨著脅迫時間的延長(第16天),野生種可溶性蛋白的含量顯著低于栽培種,干旱脅迫第32天時,栽培種可溶性蛋白含量上升達峰值且為野生種的1.41倍,而后呈顯著下降趨勢,至第48天時,野生種下降至栽培種的75%,但二者始終高于干旱脅迫前。

圖2 干旱脅迫下東北紅豆杉野生種與栽培種葉片可溶性糖(A)和可溶性蛋白(B)含量Fig.2 Soluble sugar(A)and soluble protein(B)content in leaves of wild and cultivated species of Taxus cuspidata under drought stress
2.3 干旱脅迫對東北紅豆杉野生種與栽培種葉片光合參數(shù)的影響
與干旱脅迫前相比,持續(xù)干旱脅迫抑制紅豆杉葉片的凈光合速率(Pn)、蒸騰速率(Tr)和氣孔導度(Gs)3個光合參數(shù),均呈下降趨勢(圖3)。干旱脅迫下,東北紅豆杉野生種葉片Pn、Tr和Gs均顯著高于栽培種,曲線基本一致,其中野生種Pn值在葉片光和參數(shù)中顯著高于栽培種且差值最大,干旱脅迫前約為栽培種的1.41倍,至第48天時已為栽培種的1.69倍,而栽培種Gs在干旱脅迫初期顯著高于野生種,但在第24天后則低于野生種,至第48天時約為野生種的44.1%;而細胞間隙CO2濃度(Ci)變化趨勢則與上述3項指標差異較大,呈顯著上升趨勢,干旱脅迫初期野生種Ci值略低于栽培種,持續(xù)干旱脅迫至第24天后逐漸上升且顯著高于栽培種,第48天指標約為栽培種的1.32倍。
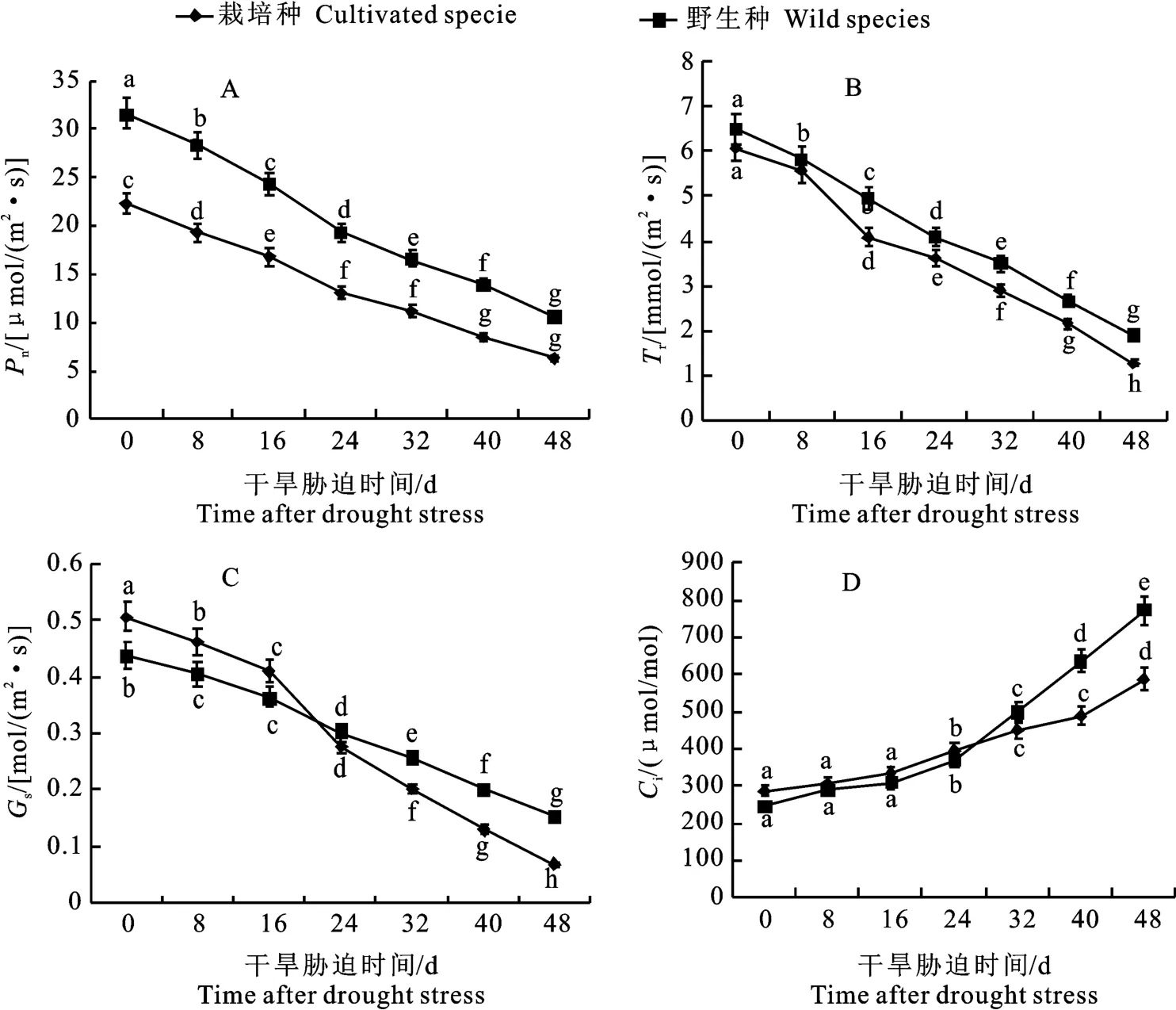
圖3 干旱脅迫下東北紅豆杉野生種與栽培種葉片光合特性Fig.3 Leaves photosynthetic characteristics of wild and cultivated species of Taxus cuspidata under drought stress
2.4 干旱脅迫對東北紅豆杉野生種與栽培種葉片色素含量的影響
干旱脅迫對植物光合作用影響較大,由于干旱脅迫促使植物進行光合作用時葉片的氣孔導度(Gs)下降,CO2進入葉片受阻,致使光合效率降低,加之葉綠體中類囊體膜及其超微結構均受損傷,最終導致紅豆杉光合色素的合成發(fā)生變化。如圖4-A所示,干旱脅迫下,紅豆杉野生種與栽培種葉片的葉綠素含量下降且顯著高于栽培種,至第48天時約為栽培種的2.23倍;而類胡蘿卜素含量上調(diào)(圖4-B),干旱脅迫初期野生種類胡蘿卜素的含量高于栽培種,第8天后栽培種含量急劇升高且顯著高于野生種,至第48天時約為野生種的1.44倍。

圖4 干旱脅迫下紅豆杉野生種與栽培種葉片色素含量Fig.4 Leaves pigment of wild and cultivated species of Taxus cuspidata under drought stress
2.5 干旱脅迫對東北紅豆杉野生種與栽培種次生代謝物的影響
脯氨酸(Pro)為滲透調(diào)節(jié)物質,在植物抵御干旱脅迫過程中發(fā)揮重要作用,與植物代謝以及水分吸收密切相關;丙二醛(MDA)是植物在逆境條件下膜脂過氧化的重要產(chǎn)物之一,能夠反映細胞膜的過氧化程度與植物抗逆能力,干旱脅迫下Pro和MDA在紅豆杉體內(nèi)會大量積累呈穩(wěn)定上升趨勢,且野生種顯著低于栽培種。干旱脅迫前,野生種Pro含量略高于栽培種(圖5-A),隨著持續(xù)干旱脅迫,野生種Pro含量顯著低于栽培種,約為栽培種的1.1倍;而干旱脅迫初期,野生種MDA含量與栽培種基本一致(圖5-B),至第24天時急劇上升,至第48天時約為栽培種的1.36倍;谷胱甘肽(GSH)在干旱脅迫下呈先上升后下降趨勢(圖5-C),但下降趨勢較為平緩,栽培種在第32天達到峰值,約為野生種的1.15倍,而后呈下降趨勢,至第48天時約為野生種的1.14倍,野生種在第40天達到峰值,約為栽培種的91.6%,后呈下降趨勢;而干旱脅迫下,類黃酮的含量均有不同程度的升高(圖5-D),且栽培種顯著高于野生種,干旱脅迫初期野生種類黃酮含量略高于栽培種,至第48天時,栽培種其含量約為野生種的1.05倍。
干旱脅迫下野生東北紅豆杉與栽培種相關酶(LDH、GCK和Cyt c)的酶活性均呈顯著上調(diào)趨勢(圖5-E,5-F,5-G),野生種的3種酶活性均始終高于栽培種,如LDH酶活性在干旱脅迫初期,野生種與栽培種無明顯差異,但在第24天后野生種LDH酶活性急劇上升為栽培種的1.24倍,至第48天時約為栽培種的1.37倍。東北紅豆杉野生種與栽培種的精氨酸和鳥氨酸轉氨酶含量隨干旱脅迫時間的延長均呈上升趨勢(圖5-H,5-I),干旱脅迫初期,野生種與栽培種二者含量基本一致,但第16天左右,栽培種的精氨酸和鳥氨酸轉氨酶含量急劇上升,分別為野生種的1.05倍和1.19倍,而栽培種的天冬氨酸含量在第16天~40天時急劇上升且顯著高于野生種(圖5-J),在第24天達到峰值,約為野生種的1.07倍,而后逐漸下降,至第48天時,約為野生種的97.28%。

圖5 干旱脅迫下紅豆杉野生種與栽培種次生代謝物的含量Fig.5 Secondary metabolites content in wild and cultivated species of Taxus cuspidata under drought stress
2.6 干旱脅迫對東北紅豆杉野生種與栽培種相關蛋白表達的影響
植物抗旱性是多信號轉導途徑以及多基因偶聯(lián)控制的復雜應答反應,在分子水平上探究紅豆杉抗旱相關基因的表達調(diào)控模式,可更加明確東北紅豆杉野生種與栽培種的抗旱機制以及野生種與栽培種的差異,分析二者干旱應答、光合響應蛋白以及紫杉醇生物合成相關基因表達水平的變化(圖6),野生種與栽培種的干旱應答元件結合蛋白DREB表達量升高(圖6-A),曲線基本一致,野生種干旱脅迫初期DREB表達量略高于栽培種,第8天后顯著低于栽培種,栽培種DREB表達量于第40天達到峰值,約為野生種的1.28倍;紫杉醇生物合成途徑中轉錄因子AP2與關鍵酶TcDBTNBT的表達量隨干旱脅迫時間而呈顯著下降趨勢,二者曲線基本一致,其中野生種AP2基因與Tc DBTNBT基因表達量始終高于栽培種;干旱脅迫后,Psa K表達水平上調(diào),干旱脅迫初期野生種與栽培種Psa K的表達量差異不大,干旱脅迫第8天后野生種光合基因的表達水平較栽培種呈顯著上調(diào)趨勢,在第24天時野生種的表達量急劇升高,約為栽培種的1.57倍,這與之前光和參數(shù)變化基本一致。
3 討論
東北紅豆杉是耐旱能力較強的植物,干旱脅迫下其形態(tài)與組織結構、干旱相關基因的表達等均有不同程度改變[16]。本研究表明,東北紅豆杉栽培種的大部分次級代謝產(chǎn)物含量高于野生種,如干旱脅迫下東北紅豆杉葉片含水量降低,其野生種葉片相對含水量始終低于栽培種。此外,干旱脅迫雖能引起東北紅豆杉形態(tài)結構、生理指標和代謝產(chǎn)物的改變,但東北紅豆杉本身為預防、降低或自我修復應答干旱帶來的損傷也會發(fā)生生理代謝的連鎖反應,以維持正常的生理狀態(tài),并且東北紅豆杉野生種與栽培種之間存在顯著差異,其中作為滲透調(diào)節(jié)物質的可溶性糖與可溶性蛋白含量受干旱脅迫的影響較大,野生種與栽培種可溶性糖含量持續(xù)增加且曲線基本一致,干旱脅迫初期二者差異不明顯,第16天后栽培種顯著高于野生種,增強了紅豆杉栽培種的滲透調(diào)節(jié)作用,可消除或修復栽培種因干旱引起的應激損傷,這與干旱脅迫下金露梅等植物可溶性糖積累趨勢一致[17]。而東北紅豆杉葉片可溶性蛋白含量呈先升高后降低的趨勢,野生種可溶性蛋白的含量顯著低于栽培種,可能由于可溶性蛋白多為生物代謝的酶類,干旱脅迫初期,由于東北紅豆杉產(chǎn)生應激反應而使體內(nèi)代謝酶的含量增加、酶活性提高,致使東北紅豆杉葉片可溶性蛋白含量增加,后期由于持續(xù)干旱脅迫抑制了紅豆杉體內(nèi)相關蛋白質的合成,加之酶快速降解等因素導致紅豆杉可溶性蛋白含量先增加后降低;細胞內(nèi)滲透物質增加是引起滲透勢下降的關鍵因素之一,體內(nèi)積累滲透調(diào)節(jié)物質以維持細胞含水量和膨壓勢,保證細胞生長、氣孔開放和光合等生理過程的進行,除可溶性糖與可溶性蛋白外,Pro亦為東北紅豆杉細胞質的主要滲透物質,Pro親水性極強,干旱脅迫下其含量顯著增加,可穩(wěn)定原生質及細胞內(nèi)的代謝過程,干旱脅迫下蛋白質合成減慢,導致Pro參與蛋白質合成量減少,Pro積累急劇升高,保持胞質內(nèi)溶膠與環(huán)境的滲透平衡,防止細胞失水,增強蛋白質的可溶性以及降低可溶性蛋白的沉淀,保護這些生物大分子結構和功能的穩(wěn)定。此外,干旱脅迫產(chǎn)生的氨亦可形成Pro,起到解毒作用[18];而MDA具有很強的毒性,是膜脂過氧化物作用的主要產(chǎn)物之一,是膜系統(tǒng)受傷害的重要標志,干旱脅迫下它能與膜上的氨基酸殘基或核酸反應生成shiff堿,增加膜透性,干旱脅迫下MDA在東北紅豆杉體內(nèi)會大量積累呈穩(wěn)定上升趨勢,且野生種MDA含量明顯低于栽培種。為防止ROS積累造成的細胞損害,植物進化出保護酶系統(tǒng)As A-GSH酶循環(huán)系統(tǒng)等抗氧化防御系統(tǒng),通過增加酶活性來清除體內(nèi)積累的ROS,干旱脅迫下,GSH呈先上升后下降趨勢,但下降趨勢較為平緩,栽培種在第32天達到峰值約為野生種1.15倍,而后呈下降趨勢,這可能是由于干旱初期GSH可以正常合成,但持續(xù)干旱破壞了GSH合成酶的結構,造成GSH生物合成量降低,這與Wada等[19]的研究結果一致。此外,糖代謝也是植物代謝的重要途徑之一,糖代謝與植物對逆環(huán)境下的抗逆性應答以及植物體生長有關,干旱脅迫下東北紅豆杉野生種與栽培種LDH、GCK和Cyt c的酶活性均呈顯著上調(diào)趨勢,且野生種始終高于栽培種,說明能量代謝方面野生東北紅豆杉顯著優(yōu)于栽培種,這可能與野生東北紅豆杉所處東北地區(qū)的地理環(huán)境以及氣溫急劇變化有關;氨基酸為植物體內(nèi)主要抗逆物質之一,干旱脅迫下大部分糖類和氨基酸顯著增加,說明東北紅豆杉通過糖類、氨基酸的顯著增加來抵御干旱脅迫,其中戊糖磷酸途徑是植物體內(nèi)干旱應答重要的葡萄糖代謝途徑之一[20]。精氨酸和天冬氨酸代謝可以減輕干旱脅迫對東北紅豆杉的損傷,具有維護東北紅豆杉穩(wěn)態(tài)的作用,說明東北紅豆杉通過精氨酸和天冬氨酸代謝通路調(diào)控干旱脅迫對紅豆杉的有害影響,增加東北紅豆杉對干旱脅迫的抵抗性和適應性。本試驗精氨酸和天冬氨酸的含量在脅迫初期時明顯高于栽培種,但隨著干旱脅迫時間的延長,第16天~40天內(nèi),栽培種應激反應產(chǎn)生的精氨酸和鳥氨酸轉氨酶含量顯著高于野生種,抗旱基因DREB表達量與之結果一致,可見,從應激代謝方面分析,東北紅豆杉栽培種對干旱脅迫的適應性和抵抗性更強。
植物光合參數(shù)會由于干旱脅迫生物量分配的變化而變化,如干旱脅迫初期,紅豆杉葉片Pn、Tr、Gs整體變化趨勢一致,而持續(xù)干旱脅迫導致Pn、Tr、和Gs均呈下降趨勢,且野生種均高于栽培種,可見紅豆杉葉片Gs降低,偶聯(lián)Pn和Tr均降低來抵御干旱脅迫;細胞間隙CO2濃度(Ci)變化趨勢與上述3項指標差異顯著,均呈顯著上升趨勢,干旱脅迫初期野生種Ci值略低于栽培種,第24天后逐漸上升且顯著高于栽培種,第48天指標約為栽培種的1.32倍,其原因可能是干旱脅迫初期,氣孔開度降低但仍可消耗CO2進行光合作用,但持續(xù)脅迫致使氣孔關閉,光合作用停止,紅豆杉葉片中CO2含量則劇烈積累增加,這與竹柳Ci變化趨勢一致[21]。干旱脅迫下紅豆杉葉片光合作用相關的蛋白Psa K表達量顯著上升且野生種明顯高于栽培種,表明干旱脅迫損傷了紅豆杉葉片葉綠體,減弱了光合作用,但野生種抵御脅迫的能力強于栽培種,這與之前光和參數(shù)的結果相一致,從光合參數(shù)分析可見,栽培種抵御干旱脅迫的能力不及野生東北紅豆杉。植株通過提高光合作用相關基因的表達水平以維持正常生長發(fā)育,光合色素含量亦可直接影響紅豆杉的光合能力[22-23],干旱脅迫下由于活性氧(ROS)代謝失衡,葉綠素分解加速導致葉綠素含量呈線性下降,野生種始終高于栽培種,而紅豆杉葉片類胡蘿卜素和類黃酮含量增加,栽培種二者含量顯著高于野生種,如干旱脅迫初期野生種類胡蘿卜素的含量高于栽培種,第8天后栽培種含量急劇升高且顯著高于野生種,至第48天時約為野生種的1.44倍,類胡蘿卜素可降低干旱脅迫葉綠素的傷害,增加紅豆杉葉片的抗性;紫杉醇生物合成途徑中相關轉錄因子AP2與關鍵酶Tc DBTNBT的表達量隨干旱脅迫時間而呈顯著下降趨勢,其中野生種表達量始終高于栽培種,說明干旱脅迫降低了紅豆杉紫杉醇的生物合成量,但野生種紫杉醇的合成量高于栽培種。
綜上所述,干旱脅迫對東北紅豆杉野生種與栽培種植株的表型特征、生理特性、光合特性、次級代謝物以及相關基因的表達水平等影響較為顯著,且干旱脅迫程度不同,東北紅豆杉野生種與栽培種植株抗旱響應度亦存在較大差異,如栽培種干旱脅迫應答的次級代謝產(chǎn)物如類黃酮等含量顯著高于野生種。東北紅豆杉栽培種在干旱應答中大部分氨基酸和有機酸的積累比例高于野生種,栽培種擁有更多抗氧化性物質和緩解逆境脅迫的調(diào)節(jié)物質,可以更快的緩解和消除逆境脅迫所帶來的傷害和影響,更好地保護植物組織和細胞,所以,栽培種對脅迫逆境下具有更強抗性和適應性;而從能量代謝、光合參數(shù)以及紫杉醇合成量方面分析,東北紅豆杉野生種顯著優(yōu)于栽培種,可通過野生種與栽培種的種間雜交將野生種的優(yōu)勢轉育到栽培種,以豐富和改善現(xiàn)有栽培種的遺傳特性或創(chuàng)造新的優(yōu)異種質材料供育種利用,尤其是培育紫杉醇含量較高的栽培種,以減少對野生東北紅豆杉的過度砍伐,此外,東北紅豆杉野生種亟需更多的保護,為進一步研究東北紅豆杉的抗逆性及保護野生東北紅豆杉奠定基礎。
