本氏煙NbMBF1c的克隆、表達及在TMV侵染過程中的功能
2022-10-28 10:25:06溫玉霞張堅王琴王靖裴悅宏田紹銳樊光進馬小舟孫現超
中國農業科學 2022年18期
關鍵詞:植物
溫玉霞,張堅,王琴,王靖,裴悅宏,田紹銳,樊光進,馬小舟,孫現超
西南大學植物保護學院植物病害生物學重慶市重點實驗室,重慶 400715
0 引言
【研究意義】煙草花葉病毒(tobacco mosaic virus,TMV)是煙草花葉病毒屬的典型代表種,可侵染十字花科、葫蘆科、茄科等植物并且危害嚴重。目前對于TMV的控制多以預防為主,而挖掘抗TMV的植物基因對于遺傳育種至關重要。多蛋白橋梁因子 1(multiprotein bridging factor 1,MBF1)是一個高度保守的轉錄輔激活蛋白,在細胞分化、脂類代謝、激素類物質調節和組氨酸類物質新陳代謝等過程中發揮重要作用。了解的抗病毒功能,對于抗病毒植株的遺傳育種具有重要意義。【前人研究進展】MBF1是植物體內一種非常保守的轉錄因子,在植物的生物脅迫與非生物脅迫中發揮著重要的功能。MBF1蛋白包括兩個結構域,C-末端的螺旋-折疊-螺旋結構域是TBP結合結構域,并且該結構域被認為在整個物種中都是保守的,N-末端是一個保守的鋅帶域,能夠與轉錄因子相結合。在植物中,MBF1蛋白與植物的生長發育和響應脅迫反應相關。擬南芥中,過表達導致擬南芥的早期開花和種子數量增加;在大豆中過表達,提高了大豆的產量。對熱脅迫下擬南芥的轉錄組進行分析發現,過表達植株中兩種海藻糖-6-磷酸合成酶的相對表達量高于野生型植株,并且在熱脅迫后過表達植株中響應脅迫反應的信號分子海藻糖水平更高,提高了植物對非生物脅迫的抗性。同時MBF1蛋白在植物響應生物脅迫也具有作用。擬南芥過表達植株中丁香假單胞菌()的增殖低于野生型植物。此外,在擬南芥感染灰霉病菌()后顯著上調表達,與野生型植株相比,過表達后病斑面積更小,并且抗病相關基因和顯著上調表達。【本研究切入點】能對包括細菌和真菌在內的多種病原物產生抗性,但其對于病毒侵染是否具有調控作用尚未報道。筆者實驗室前期研究發現,本氏煙()沉默轉錄組中的表達下調,但其對于 TMV侵染的響應尚不明確。【擬解決的關鍵問題】利用分子克隆得到全長,并利用生物信息學手段分析其蛋白特性。構建NbMBF1c熒光蛋白融合表達載體,分析其亞細胞定位;利用實時熒光定量PCR分析的組織表達與 TMV侵染脅迫表達,利用病毒介導的基因沉默技術與瞬時過表達分析對于TMV-GFP侵染的影響,實時熒光定量PCR分析影響TMV侵染的機制,為茄科作物抗病毒育種提供理論依據。
1 材料與方法
試驗于 2020—2021年在西南大學植物保護學院植物免疫與植物病害生態防控實驗室完成。
1.1 試驗材料
1.1.1 菌株、質粒與植物材料 大腸桿菌()DH5、農桿菌()GV3101購自上海唯地生物技術有限公司;VIGS沉默載體和煙草花葉病毒熒光標記載體 TMV-GFP(pSDK661)由清華大學劉玉樂教授課題組饋贈;pART27-N-eGFP、pART27-N-7*Myc載體由西北農林科技大學單衛星教授饋贈。本氏煙在恒溫育苗箱中播種(25℃,16 h光照/8 h黑暗,75%相對濕度),培養至4—6葉期左右備用。
1.1.2 試劑和引物 DNA純化回收試劑盒和質粒提取試劑盒購自北京擎科生物科技有限公司;TRIZOL、反轉錄試劑盒、高保真酶 PrimeSTAR? GXL DNA Polymerase DNA聚合酶、T4 DNA連接酶購自寶日醫生物工程(大連)有限公司;零背景平末端克隆載體Zero Background pTOPO-Blunt購自北京艾德萊生物科技有限公司;實時熒光定量試劑盒SYBR Prime qPCR Set購自重慶葆光生物技術有限公司。引物和測序由生工生物工程(上海)股份有限公司完成。引物詳見表1。
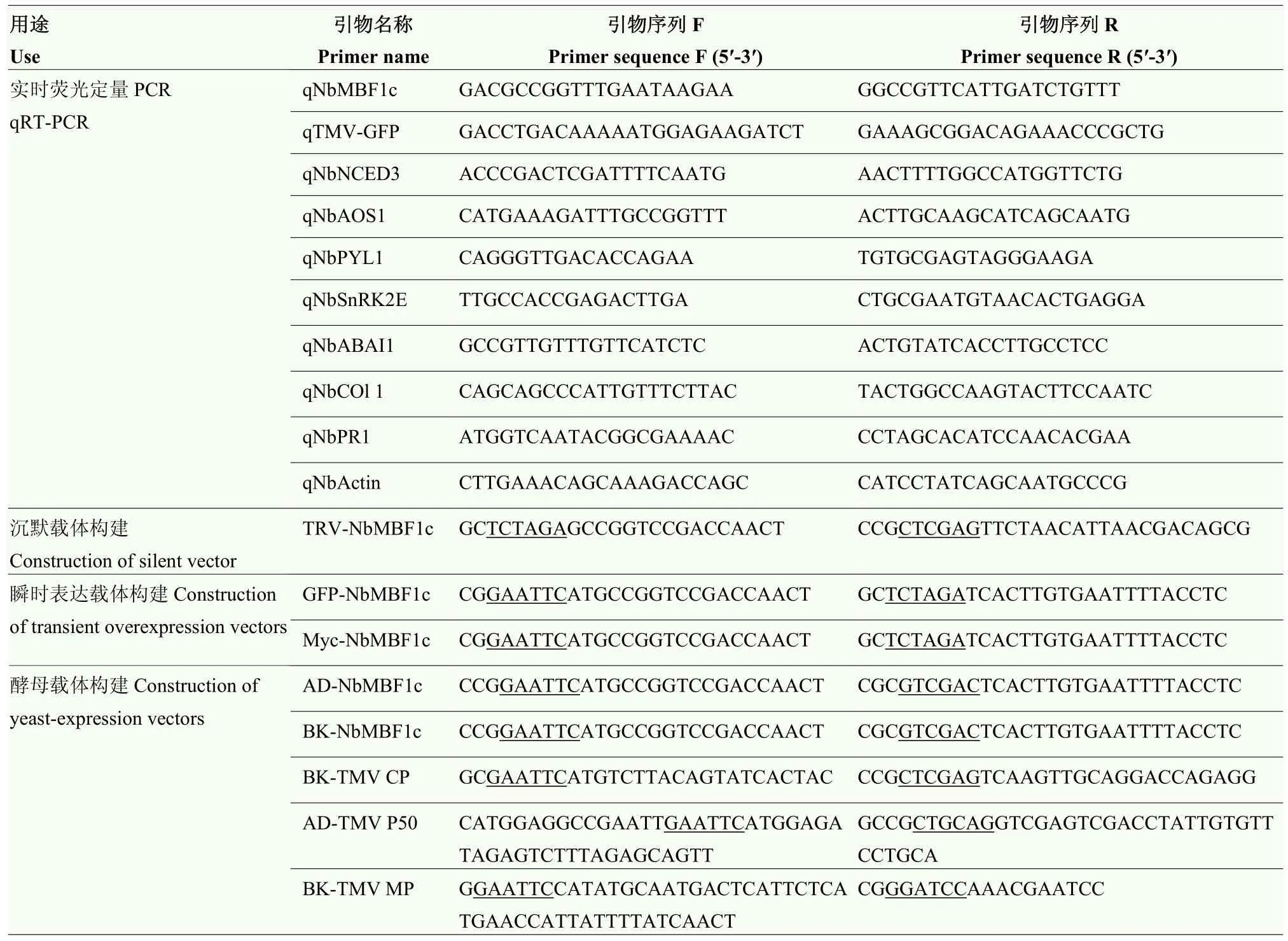
表1 本研究所用到的引物序列Table 1 Primer sequences used in this study
1.2 NbMBF1c的克隆與分析
使用TRIZOL法提取本氏煙總RNA,操作規程詳見https://www.takarabiomed.com.cn/DownLoad/9108-9109.pdf。根據Sol Genomics Network茄科作物基因組數據庫(https://solgenomics.net/)報道的核苷酸序列,設計特異引物擴增。采用PrimeSTARGXL DNA Polymerase DNA聚合酶擴增,操作詳見https://www.takarabiomed.com.cn/DownLoad/R050A.pdf。將獲得的序列連至Zero Background pTOPO-Blunt并轉化至大腸桿菌感受態細胞 DH5,操作方法詳見http://www.aidlab.cn/up_product/big/2017-4-1-10414853596.pdf。將陽性克隆菌液送至上海生工生物測序獲得序列。
運用 ExPASy ProtParam(https://web.expasy.org/protparam/)分析NbMBF1c的蛋白理化性質,TMHMM 2.0(http://www.cbs.dtu.dk/services/)分析 NbMBF1c的跨膜區域;SMART(http://smart.embl-heidelberg.de/)分析NbMBF1c的保守結構域;NCBI Blastp比對出與NbMBF1c序列相似性較高的其他物種MBF1c蛋白,利用 MEGA X軟件采用最大似然法(maximum likelihood,ML)構建系統發育進化樹,根據模型推薦,BIC最小為3 741.994時模型參數設置為JTT+G,進化樹的檢驗使用自展檢驗(Bootstrap method)法,檢驗次數1 000次。
1.3 TMV-GFP接種
TMV接種步驟參考文獻[11]。取 0.1 g被TMV-GFP侵染的本氏煙葉片于研缽中,加入石英砂和pH 7.2—7.4的磷酸緩沖液研磨均勻。將獲得的勻漿在5 000×離心機中離心3 min,取上清進行摩擦接種。每片葉片接種100 μL,每株植株接種第5、6葉位葉片。
1.4 NbMBF1c沉默載體的構建及浸潤
沉默載體的構建基于煙草脆裂病毒(tobacco rattle virus,TRV)改造的沉默體系。首先根據的全長,利用在線工具VIGS Tool(https://vigs.solgenomics.net/)獲得的最佳沉默片段,以此設計含有酶切位點H I和I的引物,將其連接在同樣酶切位點的pTRV2載體上,構建pTRV2:NbMBF1c載體。以空載體pTRV2為對照,將 pTRV1和 pTRV2:轉化至農桿菌GV3101中,28℃ 200 r/min過夜培養,利用重懸液(10 mmol·LMES,10 mmol·LMgCl,200 μmol·L乙酰丁香酮)重懸至OD=0.4,等比混合后靜止2 h并注射4葉期本氏煙。
1.5 NbMBF1c酵母載體和NbMBF1c融合表達載體構建及浸潤
根據獲得的核苷酸序列全長和 NCBI(https://www.ncbi.nlm.nih.gov/)TMV基因組的CP、MP和P50核苷酸序列設計攜帶酶切位點R I和I的引物,構建基于pGADT7和pGBKT7的酵母載體,將構建好的載體轉化大腸桿菌擴繁獲取質粒,以p53(p53)和SV40 large T-antigen(T)為陽性對照,將待驗證的質粒轉入釀酒酵母 AH109菌株,在SD-WL上生長5 d后轉染SD-WLHA和SD-WHLA+X--Gal,驗證二者的相互作用。設計攜帶酶切位點R I和I的引物,構建基于pART27的eGFP:NbMBF1c和Myc:NbMBF1c融合表達載體。將構建成功的載體轉化至農桿菌感受態細胞 GV3101中,在攜帶壯觀霉素與利福平的LB液體培養基中過夜培養,利用重懸液(10 mmol·LMES,10 mmol·LMgCl,200 μmol·L乙酰丁香酮)重懸至 OD=0.8,靜止2 h后注射6葉期本氏煙。
1.6 實時熒光定量PCR
采用qTOWER2.0 real-time PCR(Analytikjena,Germany)和 SYBR Prime qPCR Set(重慶葆光生物,中國)分析靶基因的相對表達量。使用在線工具Primer3web(https://bioinfo.ut.ee/primer3/)設計基因特異性定量引物。選擇本氏煙作為內參,使用2法定量計算基因轉錄水平的相對變化。
1.7 數據分析
所有試驗和數據至少涉及3個重復,數據表示為均值±標準誤,統計分析使用SPSS軟件student’s檢驗(*0.01<<0.05,**0.001<<0.01,***<0.001)和ANOVA單因素分析(LSD檢驗,<0.05)。
2 結果
2.1 NbMBF1c的克隆與分析
全長441 bp,編碼146個氨基酸。上傳至NCBI,獲得基因序列號ON009340。ProtParam預測顯示NbMBF1c理論分子量為16.15398 kD,理論等電點10.07,分子式為CHNOS。TMHMM 2.0分析顯示NbMBF1c蛋白不含跨膜結構。SMART分析顯示 NbMBF1c蛋白有一個保守結構域 HTH(helixturn-helix)。
根據 NbMBF1c氨基酸序列,在 NCBI中通過Blastp比對出NbMBF1c的同源蛋白,下載17條不同物種 MBF1c的氨基酸序列利用ClustalX2進行多序列比對,結合GeneDoc繪制多序列比對圖(圖1-A),利用MEGA X軟件采用ML法構建進化樹。結果如圖1-B所示,NbMBF1c與絨毛狀煙草()MBF1c(XP_009614458.1)的親緣關系最近。
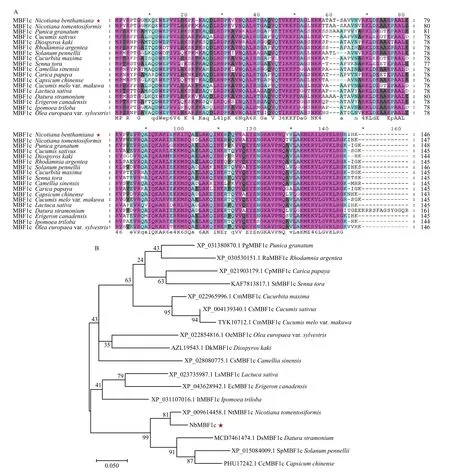
圖1 NbMBF1c和其他植物MBF1c氨基酸序列比對及其系統發育分析Fig. 1 Amino acid sequence alignment and phylogenetic analysis of NbMBF1c and MBF1c from other plants
2.2 NbMBF1c的表達分析與亞細胞定位
利用實時熒光定量PCR檢測在本氏煙各組織中的表達,發現其在根中的表達量顯著高于其他組織,莖、葉和花中的表達量遞減,表明的表達具有組織特異性(圖2-A)。為了探究響應TMV侵染的方式,對6葉期的本氏煙接種TMV-GFP并以接種PBS作為對照。選取接毒后3、5 d的接種葉和7 d的系統葉,提取其RNA并反轉錄后檢測的表達量。由圖2-B可以看出,TMV侵染后3、5和7 d,表達均極顯著高于 PBS對照,分別為對照的 47.59、499.54和 24.44倍,表明 TMV-GFP侵染后會誘導的表達,提示可能參與本氏煙抗TMV的響應。
為明確NbMBF1c的亞細胞定位,構建基于pART27的eGFP:NbMBF1c瞬時表達載體,并利用農桿菌介導的瞬時轉染技術,在本氏煙葉片表皮細胞中共表達eGFP:NbMBF1c+mCherry:00和eGFP:NbMBF1c+細胞核定位標記mCherry:H2B,48 h后在激光共聚焦顯微鏡下觀察到eGFP:NbMBF1c與mCherry:00和細胞核定位標記mCherry:H2B共定位,且利用ImageJ分析波爾遜共定位參數發現 eGFP:NbMBF1c與 mCherry:00和 mCherry:H2B都有很好的重疊性,表明 eGFP:NbMBF1c融合蛋白定位于細胞質和細胞核(圖3-A)。但是對煙草接種TMV并不能引起eGFP:NbMBF1c的定位改變(圖3-B)。
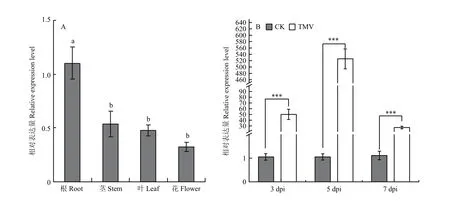
圖2 NbMBF1c的表達分析Expression analysis of NbMBF1c

圖3 NbMBF1c的亞細胞定位和TMV侵染下NbMBF1c的定位變化Fig. 3 Subcellular localization of NbMBF1c and localization change of NbMBF1c after TMV infection
2.3 沉默NbMBF1c對TMV-GFP侵染的影響
為明確對TMV-GFP侵染的影響,構建了 TRV介導的 VIGS載體 TRV:NbMBF1c。以TRV:00為對照,通過農桿菌浸潤的方式對6葉期的本氏煙進行沉默。沉默14 d后,定量結果表明TRV:NbMBF1c植株中表達量顯著低于對照植株,僅為對照植株的6.54%,說明被有效沉默(圖4-C)。相較于對照植株,沉默植株呈現矮化和開花延遲(圖4-A、4-B),提示在調控植物生長發育中可能具有重要作用。在沉默15 d時,對本氏煙摩擦接種TMV-GFP,并在紫外燈下觀察病毒侵染情況。由圖5-A可以看出,在2 dpi和 4 dpi時沉默植株接種葉上的綠色熒光數量明顯多于對照組,并且5 dpi時沉默植株系統葉上的綠色熒光顯著多于對照組,表明沉默促進了TMV-GFP的侵染。利用實時熒光定量PCR檢測接種葉(2 dpi)和系統葉(5 dpi)中TMV的含量,結果如圖5-B所示,沉默植株接種葉(2 dpi)和系統葉(5 dpi)中TMV-GFP表達量均顯著高于對照植株,分別為對照的37.4和19.2倍,與紫外燈下觀察結果一致。這些結果說明沉默促進了 TMV-GFP侵染,揭示可能為植物抗病毒正調控因子。
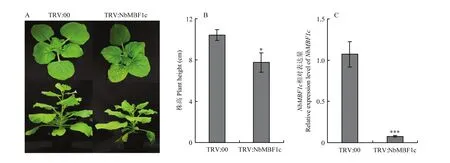
圖4 沉默NbMBF1c表型及沉默效率檢測Fig. 4 Phenotype and silencing efficiency detection in NbMBF1c silenced plant
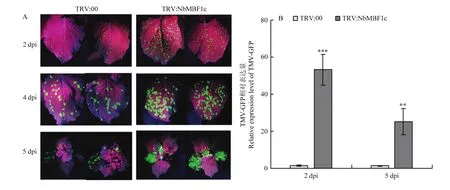
圖5 沉默NbMBF1c促進TMV-GFP侵染Fig. 5 Silencing of NbMBF1c promotes TMV-GFP infection
2.4 過表達NbMBF1c對TMV-GFP侵染的影響
TMV-GFP侵染后的表達上升,而沉默促進TMV-GFP侵染。為進一步明確在 TMV侵染時的作用,構建了重組質粒 pART27-7*Myc:NbMBF1c。以Myc:00作為對照,Myc:NbMBF1c瞬時表達48 h后,利用Western blot檢測Myc:NbMBF1c的表達情況,并接種 TMV-GFP,在紫外燈下觀察TMV-GFP的移動情況。結果顯示,約在16 kD處有一條特異條帶,與預期大小一致,而空載體無相應條帶,說明 NbMBF1c蛋白在注射葉片中成功表達(圖6-B)。接種TMV-GFP 4 d時,瞬時過表達植株葉片上出現零星的綠色熒光,而對照處理綠色熒光開始鋪滿整片葉子。接種 5 d時,瞬時過表達植株葉片上熒光數量略微擴大,而對照處理的綠色熒光已經擴散至系統葉(圖6-A)。對系統葉TMV-GFP含量進行實時熒光定量 PCR檢測,結果顯示表達Myc:NbMBF1c的植株系統葉中 TMV-GFP含量顯著低于對照,與觀察結果一致(圖6-C),表明瞬時過表達NbMBF1c抑制TMV-GFP侵染。綜合以上結果,得出 NbMBF1c作為植物正調控因子抑制 TMV-GFP侵染。
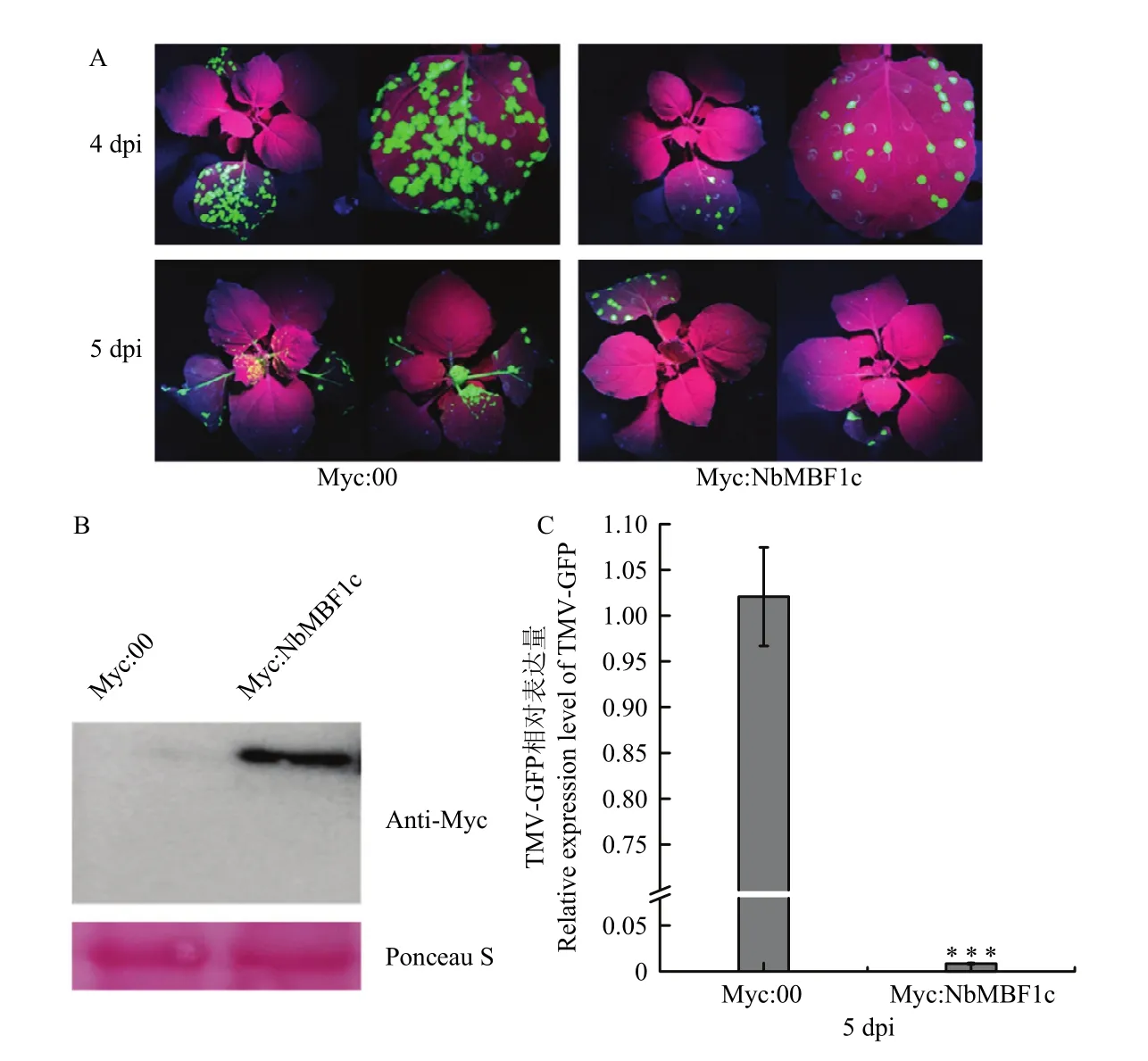
圖6 過表達NbMBF1c抑制TMV-GFP侵染Fig. 6 Overexpression of NbMBF1c inhibits TMV-GFP infection
2.5 NbMBF1c與TMV CP、MP和P50組分不互作,沉默NbMBF1c影響激素相關基因的表達
沉默顯著加快了TMV-GFP侵染(圖5),而瞬時過表達NbMBF1c則抑制了TMV-GFP侵染(圖6),表明NbMBF1c在本氏煙抵抗病毒侵染的過程中起著積極作用。筆者推測NbMBF1c可能與病毒直接作用,采用酵母雙雜交試驗來驗證此猜測,但結果發現NbMBF1c與TMV CP、MP和P50并未在酵母細胞內發生互作(圖7)。NbMBF1c是一種高度保守的轉錄因子,其能與下游多個抗性基因啟動子結合,因此筆者猜測可能會通過影響激素途徑來參加植物抗病毒。為驗證此猜測,對沉默植株中脫落酸(abscisic acid,ABA)合成關鍵酶基因、脫落酸受體ABA信號轉導因子和蛋白激酶,茉莉酸(jasmonic acid,JA)合成關鍵酶基因茉莉酸信號受體的表達情況進行檢測。結果表明,TRV:NbMBF1c中與ABA通路相關基因都顯著上調表達,、、和分別是對照表達量的1.71、1.67、4.62和2.05倍(圖8-A—8-D)。TRV:NbMBF1c中表達顯著高于對照組,較對照增加了1.29倍,但的表達較對照顯著下調,為對照的63%(圖8-E、8-F)。這些結果揭示可能通過調控ABA和JA通路來影響TMV-GFP侵染。

圖7 酵母雙雜交分析NbMBF1c與TMV病毒組分的互作Fig. 7 The interaction of NbMBF1c and TMV virus components was analyzed by Y2H
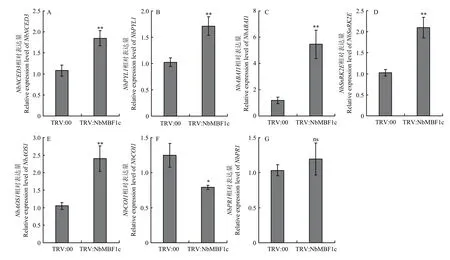
圖8 沉默NbMBF1c對ABA和JA相關基因表達的影響Fig. 8 Effect of NbMBF1c silencing on expression of ABA and JA related genes
3 討論
3.1 MBF1c廣泛參與植物的脅迫反應
在擬南芥、馬鈴薯、小麥、番茄等植物中,參與了植物的生物脅迫和非生物脅迫反應。根據這些報道,利用RT-PCR技術獲得本氏煙全長序列441 bp,編碼146個氨基酸,命名為。MBF1家族蛋白是在植物中廣泛存在參與植物調節的一類共轉錄因子,小麥上和在根、葉片和籽粒中均有表達,主要在葉片中表達,而僅在根部中特異性表達;百合中的在葉中表達量最高,根中次之,鱗莖中最低;而本氏煙在根中表達量最高,莖、葉、花依次減少。構建的進化樹表明,NbMBF1c與絨毛狀煙草MBF1c(XP_009614458.1)的親緣關系最近。這些來自其他物種共有HTH結構域的MBF1c蛋白能受到多種非生物脅迫誘導表達并且在提高植物抗逆能力上有積極作用。在多種非生物脅迫下,如熱脅迫、過氧化氫處理、干旱和鹽脅迫,的表達量會顯著上調;AtMBF1c作為轉錄因子還能通過與TPS5相互作用并與啟動子區域結合來調節擬南芥海藻糖合成并正調控對灰霉病菌的防御反應;在小麥上,受到小麥條銹菌(f. sp.)誘導高表達;過表達轉基因番茄具有抗病原細菌DC3000和尖鐮孢()的能力。本研究表明,TMW侵染下極顯著上調表達(圖2-B),并且沉默促進TMV-GFP侵染(圖5),而瞬時過表達 NbMBF1c抑制TMV-GFP侵染(圖6),這些結果提示本氏煙的 NbMBF1c蛋白在植物病毒侵染過程中具有積極正調控作用。
3.2 TMV不能改變 NbMBF1c的亞細胞定位,并且NbMBF1c不通過與 TMV組分的互作來影響病毒侵染
在激光共聚焦顯微鏡下觀察本氏煙葉片表達的eGFP:NbMBF1c融合蛋白定位在細胞質和細胞核中(圖3),這與其他物種MBF1c亞細胞定位結果一致。洋蔥表皮細胞定位試驗顯示小麥的 TaMBF1c主要定位在細胞核,部分定位在細胞質中;百合常溫下LIMBF1c定位在細胞膜、細胞質和細胞核,熱應激下LIMBF1c主要定位在細胞核中;而擬南芥MBF1c-GFP融合蛋白主要定位在細胞質中;值得注意的是,高溫脅迫下三者的 MBF1c都發生了定位改變,大部分聚集在細胞核,MBF1c在響應熱應激上起到了重要作用,但在本試驗中,對煙草接種TMV并不能引起eGFP:NbMBF1c的定位改變(圖3)。
擬南芥AtMBF1a和AtMBF1b均能與番茄花葉病毒(tomato mosaic virus,ToMV)的運動蛋白(movement protein,MP)互作,煙草的NtMBF1a能與 ToMV和十字花科植物煙草花葉病毒(crucifer tobamovirus,CTMV-W)的MP蛋白發生互作。然而,本研究酵母雙雜交試驗表明NbMBF1c與TMV病毒組分CP、MP和P50均不在酵母體內互作(圖7),表明本氏煙NbMBF1c可能不通過與病毒組分直接互作參與抗病毒防御。
3.3 NbMBF1c通過調控植物激素的產生和信號傳導來抑制TMV侵染
植物激素是一類能調節植物生長發育的微量有機物質,在非生物脅迫和生物脅迫中均發揮著重要作用。在沉默后,ABA合成關鍵酶基因脫落酸受體、ABA信號轉導因子和蛋白激酶均顯著上調表達,這一定程度上反映了ABA激素的升高。外施ABA可以促進水稻黑條矮縮病毒(rice black-streaked dwarf virus,RBSDV)侵染,沉默會抑制病毒的侵染,這說明ABA負向調節水稻對RBSDV的抗性。外施茉莉酸甲酯可以促進TMV的侵染和移動,的沉默降低了TMV的積累水平,在本氏煙上沉默能夠促進病毒的侵染。這與本研究結果是一致的,即沉默后,茉莉酸合成途徑上調表達,茉莉酸信號通路上的受體下調表達(圖8-E、8-F),TMV積累增加。在植物激素抵抗病毒侵染中,茉莉酸和水楊酸(salicylic acid,SA)是一對拮抗激素。有研究表明,JA可以調節 SA的產生,在本氏煙受到TMV-GFP侵染時,立即激活JA信號以建立早期的系統性抗性,隨后JA信號調控SA產生,激活SA信號,建立廣泛的系統抗性;擬南芥上,丁香假單胞菌誘導產生MeSA需要JA的生物合成,并依賴和介導的JA下游信號傳導。后續的試驗發現,沉默后,水楊酸通路的表達量沒有變化(圖8-G),筆者推測是由于表達量下調,JA信號傳遞給SA效率降低,SA不能在植株受到病毒侵染時起到可靠的防御作用。這些結果表明本氏煙通過影響ABA和JA信號途徑進而調控寄主抗TMV。
4 結論
的表達受 TMV侵染誘導。NbMBF1c主要定位于細胞質和細胞核中,能夠作為 TMV的抗性基因抑制病毒在本氏煙上的侵染。NbMBF1c不通過直接與 TMV組分互作來抑制病毒侵染,可以通過調控植物激素的產生和信號傳導來抑制病毒侵染。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
