缺氧誘導Caveolin-1靶向Vimentin調控肝癌細胞遷移及侵襲*
2022-10-19 03:55:08李英王興李永寧劉鵬劉松柏潘耀振
貴州醫科大學學報 2022年8期
李英,王興,李永寧,劉鵬,劉松柏,潘耀振
(貴州醫科大學 臨床醫學院 外科學教研室,貴州 貴陽 550004)
如何有效防治肝癌的復發及轉移是改善肝癌患者預后、提高生存率的關鍵[1]。缺氧是實體腫瘤微環境的基本特征之一,在腫瘤的生長、轉移、放化療耐藥中發揮重要作用[2-5]。缺氧標志物在肝癌中可作為獨立預后因素,其表達預示著肝癌患者的不良預后[6]。缺氧參與惡性腫瘤侵襲、轉移的一個重要機制就是對腫瘤細胞上皮間質轉化(epithelial-mesenchymal transition, EMT)的調節作用[7-8]。EMT是由調節因子經過各種信號轉導通路導致上皮細胞特性消失,而表達間質細胞特性的細胞轉化過程。研究證實,EMT的發生可促進肝癌的侵襲及轉移[9-10]。小窩蛋白-1(caveolin-1, CAV1)是細胞膜小窩(caveolae)的主要支架蛋白,參與了細胞增殖、信號轉導,影響了腫瘤的發生、發展和轉移[11-12],其異常表達在多種類型的腫瘤中被研究報道,特別是在晚期癌癥轉移灶中表達明顯上調[13]。CAV1的過度表達與腫瘤侵襲性臨床行為和不良預后密切相關,在肝癌中已有少數研究報道[14-15]。然而,CAV1在缺氧條件下的作用機制尚不清楚,本研究旨在分析CAV1表達與缺氧微環境的關系,探討其對肝癌細胞遷移、侵襲能力的影響及分子機制。
1 材料與方法
1.1 材料
正常肝細胞系LO2及肝癌細胞系HepG2、HCC-9810、MHCC-97H和Hep3B購自上海生科院細胞資源中心,DMEM培養基、0.25%胰蛋白酶及胎牛血清購自Gibco公司,青霉素-鏈霉素雙抗購自Hyclone公司,CAV1、Vimentin、E-cadherin及N-cadherin抗體購自武漢三鷹公司,β-actin抗體購自CST公司,羊抗鼠及羊抗兔二抗購自博士德生物公司,siRNA及轉染試劑盒購自銳博生物科技公司,Protein A+G Agarose beads購自碧云天公司,CCK-8試劑盒購自日本同仁。
1.2 研究方法
1.2.1細胞培養及缺氧模型構建 細胞接種于含10%胎牛血清及1%青鏈霉素雙抗的DMEM培養基中,于37 ℃、5% CO2的恒溫培養箱中常規培養。取生長狀態良好的MHCC-97H與Hep3B細胞接種于96孔板中(4 000個/孔),待細胞貼壁后,更換為含50、100、150、200、250 μmol/L氯化鈷(CoCl2)的培養基繼續培養24 h,對照組不予CoCl2干預,采用CCK-8法檢測細胞活力。選取100 μmol/L CoCl2分別處理細胞0、12、24、48、72 h后, CCK-8法檢測不同時間下細胞活力。酶標儀檢測波長為450 nm,細胞活力(%)=(處理組OD450值/對照組OD450值)×100%。
1.2.2siRNA轉染 siRNA由銳博生物公司(廣州、中國)構建合成。取生長狀態良好的MHCC-97H和Hep3B細胞接種于6孔板中,細胞融合度達70%時,按照說明書采用轉染試劑盒將干擾序列(si-CAV1)及干擾對照序列(si-NC)轉染MHCC-97H和Hep3B細胞,12 h后更換培養基,48 h后收集細胞用于進一步的實驗。
1.2.3qPCR 采用TRIzol試劑提取細胞總RNA,以cDNA逆轉錄試劑盒進行逆轉錄,以 SYBR?Premix Ex TaqTM進行熒光定量PCR分析。每組設置5個復孔,反應條件:95 ℃ 30 s,95 ℃ 5 s,60 ℃ 30 s,擴增40個循環。數據采用Bio-Rad CFX Manager 軟件進行處理。結果取平均值進行計算,管家基因GAPDH作為內參,采用2-ΔΔCt計算目的基因mRNA的相對表達量。引物序列:CAV1 正向引物為5′-GCGACCCTAAACACCTCAAC-3′,反向引物為5′-ATGCCGTCAAAACTGTGTGTC-3′。
1.2.4蛋白質免疫印跡 采用RIPA裂解液提取細胞中的總蛋白,行10% SDS-PAGE 凝膠電泳分離蛋白,300 mA濕轉120 min將目的蛋白轉移至PVDF膜上,封閉液室溫封閉2 h后分別將目的條帶放入對應的一抗溶液中4 ℃孵育過夜;次日洗滌條帶后放入羊抗鼠(1∶2 000)或羊抗兔(1∶2 000)二抗中,室溫孵育1 h。采用ECL法對條帶進行顯影,以β-actin作為內參比較目的蛋白CAV1表達。
1.2.5劃痕實驗 細胞計數按5×105(個/孔)均勻的接種于6孔板內,細胞融合度達到90 %時,用已消毒的直尺置于孔徑上,固定直尺兩端勿挪動,用200 μL槍頭沿著直尺劃垂直線,用PBS清洗2遍后加入2 mL無血清DMEM培養基,立即于倒置顯微鏡下觀察并拍照,待培養48 h后再觀察拍照。
1.2.6Transwell細胞遷移及侵襲實驗 細胞消化、無血清培養基重懸計數,按2×104(個/孔)接種于未包被或基質膠包被的Transwell細胞培養小室內,補充無血清培養基至總體積200 μL。下室加入700 μL含10%胎牛血清的DMEM培養基。常規培養24 h后取出小室,用棉簽去除濾膜內側的細胞,加入4%多聚甲醛固定30 min,1%結晶紫染色30 min,清洗掉多余的結晶紫染料后烘干,顯微鏡下觀察并拍照。
1.2.7免疫共沉淀 細胞中加入500 μL NP40裂解液(蛋白酶抑制劑10 μL、磷酸酶抑制劑10 μL及PMSF 5 μL)提取總蛋白,加入20 μL Protein A+G Agarose beads于4 ℃搖床孵育2 h,對蛋白提取物進行預純化。將純化后的提取物分為3份,分別加入純化IgG(IgG組)、CAV1抗體(IP組)、留50 μL不加任何抗體作為對照組(Input組),于4 ℃搖床孵育過夜。IgG組及CAV1組孵育后的提取物中各加入40 μL Protein A+G Agarose beads,收集沉淀加入蛋白上樣緩沖液100 ℃變性5 min,采用SDS-PAGE蛋白凝膠電泳進行檢測分析。
1.3 統計學方法
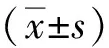
2 結果
2.1 肝癌細胞系中CAV1的表達
CAV1 mRNA及蛋白的表達在HepG2、HCC-9810、MHCC-97H、Hep3B等肝癌細胞系中較人正常肝細胞LO2中的表達量高(P<0.05),并且在MHCC-97H和Hep3B細胞中的表達最明顯。見圖1。故選取MHCC-97H及Hep3B細胞進行后續缺氧模型的構建。
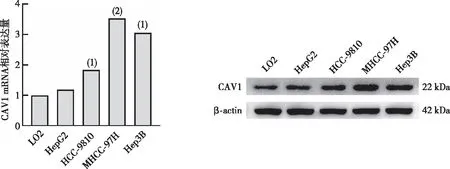
注:與LO2比較,(1)P<0.01,(2)P<0.001。
2.2 缺氧對肝癌細胞中CAV1表達的影響
以0、50、100、150、200、250 μmol/L CoCl2誘導MHCC-97H及Hep3B細胞缺氧24 h,CCK-8實驗結果顯示,0~100 μmol/L CoCl2時,肝癌細胞的生長無明顯影響;當濃度高于150 μmol/L后,MHCC-97H及Hep3B 細胞的存活率均有不同程度的下降。選取100 μmol/L CoCl2分別處理細胞0、12、24、48、72 h,結果顯示,24 h時,細胞的增殖活性最大(P<0.05,見圖2),故以100 μmol/L CoCl2作用24 h作為細胞缺氧模型的最佳濃度和時間,與對照組相比(0 h),缺氧環境明顯促進了CAV1蛋白表達。見圖3。
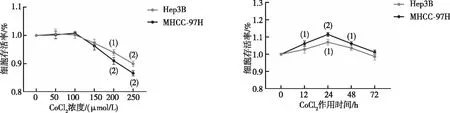
注:與0 μmol/L或0 h比較,(1)P<0.05,(2)P<0.01。
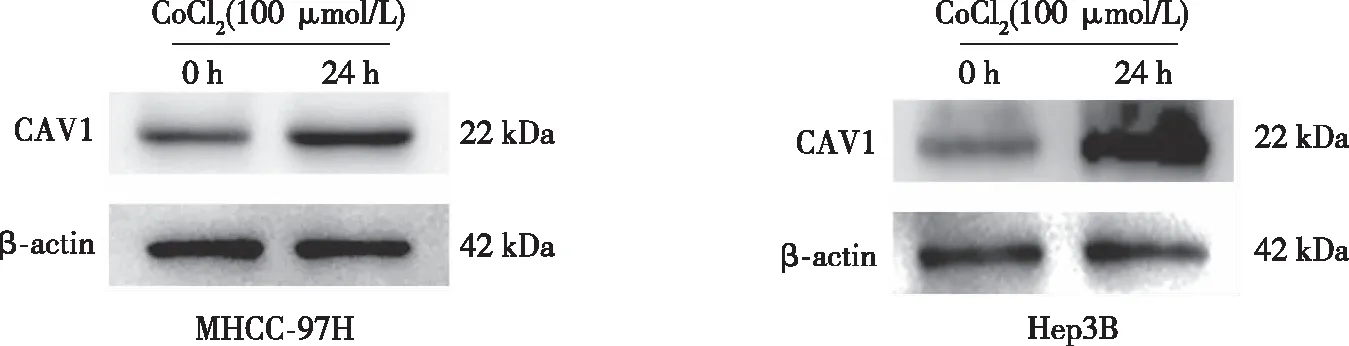
圖3 缺氧對肝癌細胞中CAV1表達的影響
2.3 缺氧對肝癌細胞遷移、侵襲能力的影響
劃痕實驗結果示,與對照組比較(0 h),MHCC-97H及Hep3B細胞在缺氧環境下細胞的相對遷移率明顯增加(P<0.05)。見圖4。Transwell實驗結果示,與對照組相比(0 h),缺氧明顯促進了細胞遷移和侵襲(P<0.05)。見圖5。

注:與0 h比較,(1)P<0.01。
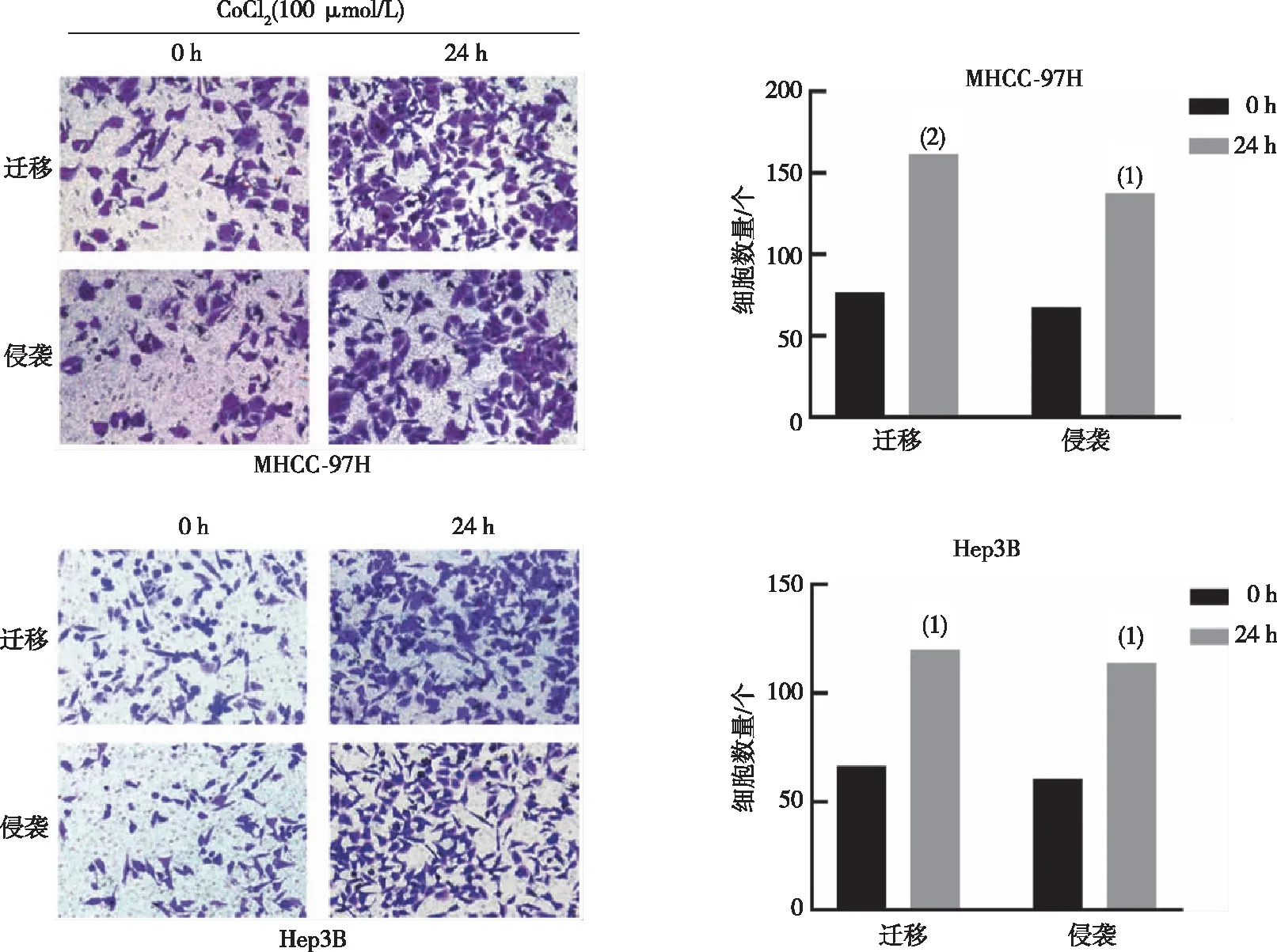
注:與0 h比較,(1)P<0.05,(2)P<0.01。
2.4 缺氧對肝癌細胞中EMT相關指標的影響
Western blot結果顯示,與對照組相比(0 h),缺氧環境下EMT相關指標Vimentin、N-cadherin蛋白明顯增加,E-cadherin的表達明顯降低(P<0.05)。見圖6。

圖6 缺氧對MHCC-97H及Hep3B細胞中EMT相關指標表達的影響
2.5 CAV1對肝癌MHCC-97H及Hep3B細胞遷移、侵襲的調控作用
采用siRNA干擾CAV1的表達后,進一步驗證在缺氧環境中CAV1對肝癌細胞株MHCC-97H及Hep3B轉移能力的影響。劃痕實驗結果示,與對照組相比(si-NC),干擾CAV1的表達后細胞的相對遷移率明顯減弱,差異有統計學意義(P<0.05)。見圖7。Transwell細胞遷移及侵襲實驗結果示,與對照組相比(si-NC),si-CAV1組細胞的遷移及侵襲能力明顯降低,差異有統計學意義(P<0.05)。見圖8。
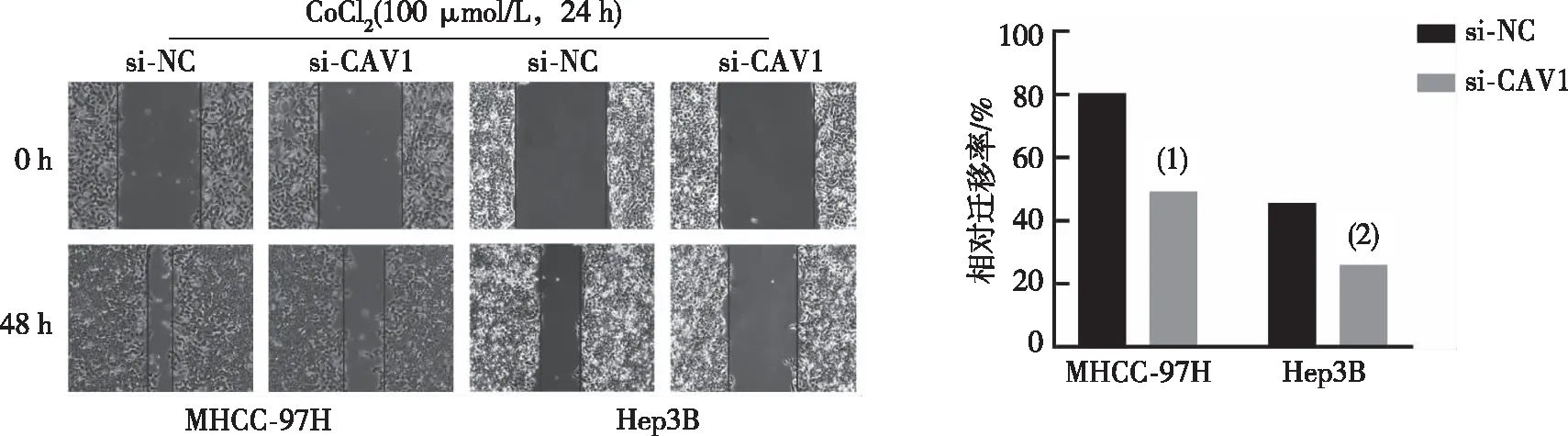
注:與si-NC相比,(1)P<0.001,(2)P<0.01。

注:與si-NC比較,(1)P<0.05,(2)P<0.01。
2.6 CAV1對肝癌細胞中EMT相關指標的影響
Western blot實驗結果示,缺氧環境下與si-NC組比較,下調CAV1表達后MHCC-97H及Hep3B細胞中Vimentin、N-cadherin蛋白表達水平明顯降低,E-cadherin表達明顯增加(P<0.05)。見圖9。

圖9 下調CAV1對肝癌細胞株MHCC-97H及Hep3B中EMT相關指標的影響
2.7 CAV1可通過招募Vimentin促進肝癌的進展
免疫共沉淀實驗顯示(圖10),在MHCC-97H及Hep3B細胞中,CAV1能與Vimentin在蛋白水平存在直接或間接結合的可能性。

圖10 CAV1與EMT互作關系的驗證
3 討論
腫瘤細胞轉移作為肝癌惡性表型的特征之一,提示了肝癌患者的不良預后。腫瘤細胞及其微環境的相互作用與腫瘤的侵襲、轉移密切相關[16]。實體腫瘤由于生長過程中腫瘤細胞的快速增殖、局部微血管形成相對滯后,使局部腫瘤組織的血供不足導致腫瘤細胞處于相對缺血缺氧的環境[17]。研究表明,在缺氧微環境的作用下,腫瘤細胞的粘附能力降低,可主動從原發灶脫離侵入間質、血管或淋巴系統,然后定植在遠處組織中形成轉移灶[18]。而該過程受多種調控因子、基因和信號通路的調控,具體機制還有待進一步研究。在肺癌、結直腸癌及胃癌等多種實體瘤中,研究發現CAV1能促進腫瘤侵襲性、藥物耐受性、抗凋亡等特性[19]。本研究結果提示CAV1在肝癌中發揮促進細胞遷移及侵襲的作用。首先,qPCR及Western blot結果顯示CAV1在多株人肝癌細胞系中高表達,提示其一定程度上發揮著促癌作用。缺氧微環境已被證明可以選擇侵襲性較強的癌細胞在缺氧條件下適應和生存。因此,了解與肝癌細胞侵襲性相關的缺氧基因是非常重要的。通過構建細胞缺氧模型發現,在缺氧應激條件下CAV1表達進一步增強,功能上表現為促進MHCC-97H及Hep3B細胞的體外遷移和侵襲能力。通過小干擾RNA技術下調CAV1的表達后,明顯抑制了缺氧環境下細胞的遷移及侵襲。上述結果表明缺氧微環境中CAV1對肝癌細胞的遷移及侵襲能力具有重要的調控作用。
近年來EMT被認為是實體腫瘤轉移的初始環節,被廣泛關注和研究[20]。EMT的典型特征是上皮細胞標志物下調,如E-cadherin、緊密連接蛋白-1(ZO-1),同時伴隨間質標志物表達的升高,如Vimentin、N-cadherin;且有研究發現,腫瘤的缺氧微環境可誘導EMT的發生,通過改變能量代謝途徑、促進血管形成、改變基因表型等方式促進腫瘤細胞適應缺氧,促進細胞發生侵襲和轉移[21]。本研究首先觀察到缺氧條件下肝癌細胞中E-cadherin表達減少,Vimentin和N-cadherin表達增加。進一步分析CAV1促進肝癌細胞遷移及侵襲的分子機制,研究發現干擾CAV1表達后明顯抑制了Vimentin、N-cadherin的表達,而促進了E-cadherin的表達。上述結果表明,CAV1在缺氧微環境中能調控肝癌細胞的EMT過程。最重要的是,本研究通過免疫共沉淀實驗發現CAV1與Vimentin在蛋白水平存在直接或間接結合的可能。
綜上所述,一定程度的缺氧能夠促進肝癌細胞的遷移及侵襲能力,其機制可能與CAV1的表達水平增加、激活EMT途徑促進了肝癌細胞的侵襲性有關。因此,CAV1可能作為分子靶點之一在肝癌缺氧微環境調控中發揮重要作用,并為下一步探索CAV1是否可作為預后標志物在臨床腫瘤診斷和治療中發揮價值提供理論基礎。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中國生殖健康(2019年11期)2019-01-07 01:28:02
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
