中藥內生真菌的生物活性物質及其多組學研究進展
2022-10-14 07:02:02郝艷琦潘英妮趙司南李譚譚李寧孫奕
中國抗生素雜志 2022年8期
郝艷琦 潘英妮 趙司南 李譚譚 李寧 孫奕,*
(1 沈陽藥科大學 中藥學院,沈陽 110016;2 中國中醫科學院中藥研究所,北京 100700)
近年來,由于耐藥病原微生物和新疾病的出現,加之合成藥物在治療疾病過程中時常伴有的毒副作用,導致世界范圍內對天然藥物的需求不斷增加,從而促使科學家們尋找多種藥用資源來應對各種挑戰。內生微生物是生物活性代謝物的豐富來源,其次級代謝物(包括生物堿、萜類、聚酮等)具有抗腫瘤、抗炎、抗菌、抗氧化等作用,其潛在的藥用價值可作為重要的天然先導化合物進行開發利用。盡管有許多關于內生真菌生物活性化合物的研究報道,但內生真菌的研究仍處于起步階段。1993年,Stierle首次從藥用植物短葉紅豆杉的韌皮部中獲得一株產抗癌物質紫杉醇的內生真菌Taxomyces andreanae[1]。此后,內生真菌的研究受到越來越多的研究者重視[2]。本文綜述了近10年中藥來源的內生真菌次級代謝產物多樣性及其生物活性,以及與次級代謝產物相關的代謝組學、基因組學、蛋白組學等多組學研究,以概括中藥來源內生真菌活性次級代謝產物的多維度研究現狀。
1 中藥內生真菌次級代謝產物的多樣性
1.1 聚酮類
從紅豆杉莖內生真菌ColletotrichumS8中分離得到1個多肽-聚酮雜合12元環狀三肽colletopeptides A(1),結構中含有3,5,11-三羥基-2-甲基十二酸單元。Colletopeptide A可抑制脂多糖誘導的RAW264.7巨噬細胞產生NO(IC50值為8.3 μmol/L),并能夠抑制炎癥因子IL-6和TNF-α的產生,降低NF-κB相關蛋白IκBα和p65的磷酸化[3]。Yang等[4]從丹參地上部分分離到內生真菌球毛殼菌Chaetomium globosum,從其固體發酵產物中分離到2個新聚酮類化合物aureonitols A和B(2-3)和已知化合物equisetin(4)。其中,化合物4對4種多重耐藥菌(糞腸球菌、糞腸球菌、金黃色葡萄球菌和表皮葡萄球菌)具有顯著的抗菌活性,其MIC值為3.13~6.25 μg/mL。另外,從丹參的地上部分還分離出內生真菌Colletotrichum gloeosporioidesXL1200,從該真菌中的固體發酵培養物中鑒定了2個新的乙氧基苯乙酮衍生物colletotricholides A(5)和B(6)[5],化合物1~6結構式見圖1。
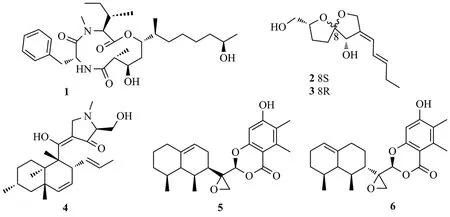
圖1 化合物1~6結構式Fig.1 The structures of compounds 1~6
從三七內生真菌裸孢殼屬EmericellaXL029分離得到4 個異戊二烯黃嘌呤型化合物14-hydroxyltajixanthone(7)、14-hydroxyltajixanthone hydrate(8)、14-hydroxyl-15-chlorotajixanthone hydrate(9)、epitajixanthone hydrate(10),上述化合物對除耐藥金黃色葡萄球菌外的所有測試革蘭陽性菌和陰性菌均有顯著的抗菌活性,MIC值為12.5~50 μg/mL[6]。從另1株三七內生真菌康氏木霉屬Trichoderma koningiopsisYIM PH30002中分離到2個新聚酮化合物koninginins O和Q(11和12),這2個化合物對尖孢鐮刀菌Fusarium oxysporum和黃瓜枯萎病菌Plectosphaerella cucumerina表現出較弱的抑菌活性(MIC值為128 μg/mL)[7]。此外,Yang等[8]從三七內生青霉菌PenicilliumYIM PH30003發酵液中分離到一個多氧芐基環己酮penginsengin A(13),該化合物對三七病原菌Fusarium.solani具有一定的抗菌活性(MIC值為64 μg/mL),化合物7~13結構式見圖2。

圖2 化合物7~13結構式Fig.2 The structures of compounds 7~13
Biasettoa等[9]從喜樹內生真菌Phomopsis stipata代謝物中分離到新化合物koninginins T(14)和U(15),其中,化合物14在10 μg時表現出抑制乙酰膽堿酯酶的作用。Wu等[10]從金錢草葉組織內生真菌Byssochlamys spectabilis中分離得到一種新骨架聚酮衍生的八酮二聚體byspectin A(16),該化合物是一種新型高選擇性的hCE2抑制劑(IC50值為2.01 μmol/L)。Feng等[11]從華布貝母的健康鱗莖中分離得到一種球孢菌Cladosporium sphaerospermumWBS017,在大米培養基上發酵,分離得到2個聚酮類化合物cladosins L和N(17和18),其中17對2種金黃色葡萄球菌(Staphylococcus39aureusATCC 29213和S.aureusATCC 700699)具有抗菌作用(MIC值為50和25 μmol/L);而在白豆培養基上培養這種真菌,得到了另外1種雜合聚酮cladosin O(19),化合物14~19結構式見圖3。

圖3 化合物14~19結構式Fig.3 The structures of compounds 14~19
Shi等[12]從艾蒿的根組織中分離得到內生真菌Trichoderma koningiopsisQA-3,從中純化到雙環聚酮類化合物15-hydroxy-1,4,5,6-tetra-epi-koninginin G(20)和14-hydroxykoninginin E(21),化合物20對水生病原菌溶藻弧菌Vibrio alginolyticus具有顯著的抗菌活性(MIC值為1 μg/mL)。從艾蒿內生真菌Trichoderma koningiopsisQA-3的培養液中分離鑒定出具有八氫萘骨架的三環聚酮化合物ent-koninginin A(22),化合物22對人類病原體大腸埃希菌(Escherichia coli)、海洋水生病原體溶藻弧菌(Vibrio.alginolyticus)和鰻鱺弧菌(V.anguillarum)等顯示出較強的抑菌活性(MIC值為8~64 μg/mL),化合物22抗創傷弧菌(V.vulnificus)的活性(MIC,4 μg/mL)強于對照藥氯霉素(MIC,8 μg/mL)[13]。Liu等[14]從巴戟天內生真菌Cytospora rhizophoraeA761中分離到4個聚酮類異二聚體cytorhizins A-D(23~26),它們具有6/6/5/6/8或6/6/5/6/7五環系統,形成籠狀骨架,由聚氧化異戊基單元和高度結構化的二苯甲酮骨架組成。化合物24和26對HepG-2、MCF-7、SF-268和A549細胞均表現出較弱的細胞毒性,IC50值在29.4~68.6 μmol/L之間,化合物20~26結構式見圖4。
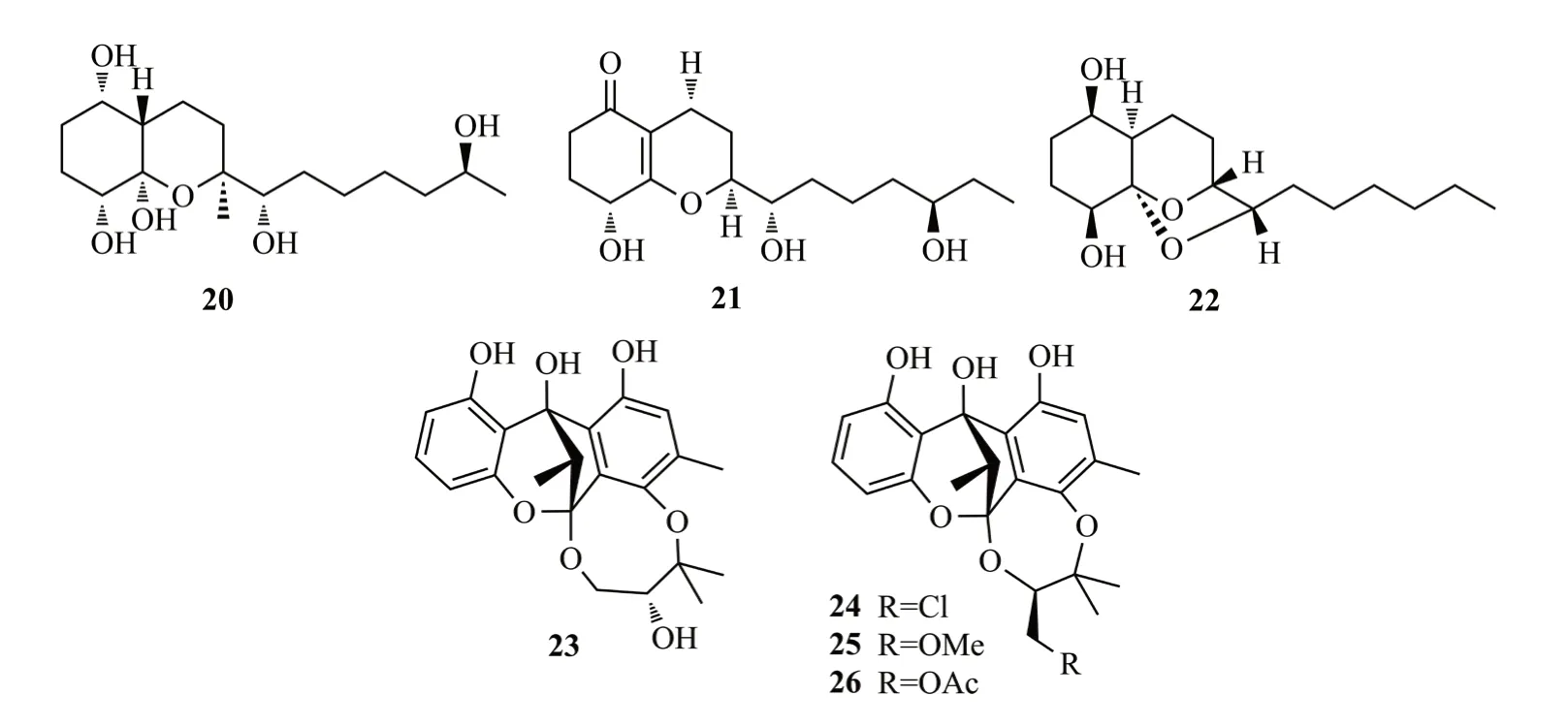
圖4 化合物20~26結構式Fig.4 The structures of compounds 20~26
Phomopsin A(27)是從大葉藤黃內生真菌PhomopsisCAM212的固體培養物中分離出的聚酮化合物,乙酰化的27a以濃度依賴的方式能夠顯著抑制酯多糖誘導的RAW264.7細胞NO的生成(IC50值為14.8 μmol/L)[15]。Bang等[16]從桑樹葉內生毛盤孢菌ColletotrichumJS-0361中分離到具有5/6/10融合環系統的聚酮化合物,命名為colletotrichalactones A和Ca(28和29)。其中,化合物28具有不穩定的醛基,在溶劑中易轉化為縮醛基。以上兩個化合物對MCF7細胞顯示較弱的細胞毒性。Antonius等[17]從蒺藜內生真菌煙曲霉Aspergillus fumigatiaffnis中分離到1個抗菌聚酮(-)palitantin(30),能抑制糞腸球菌Enterococcus faecalisUW2689和肺炎鏈球菌Streptococcus pneumonia(MIC值為64 μg/mL),化合物27~30結構式見圖5。
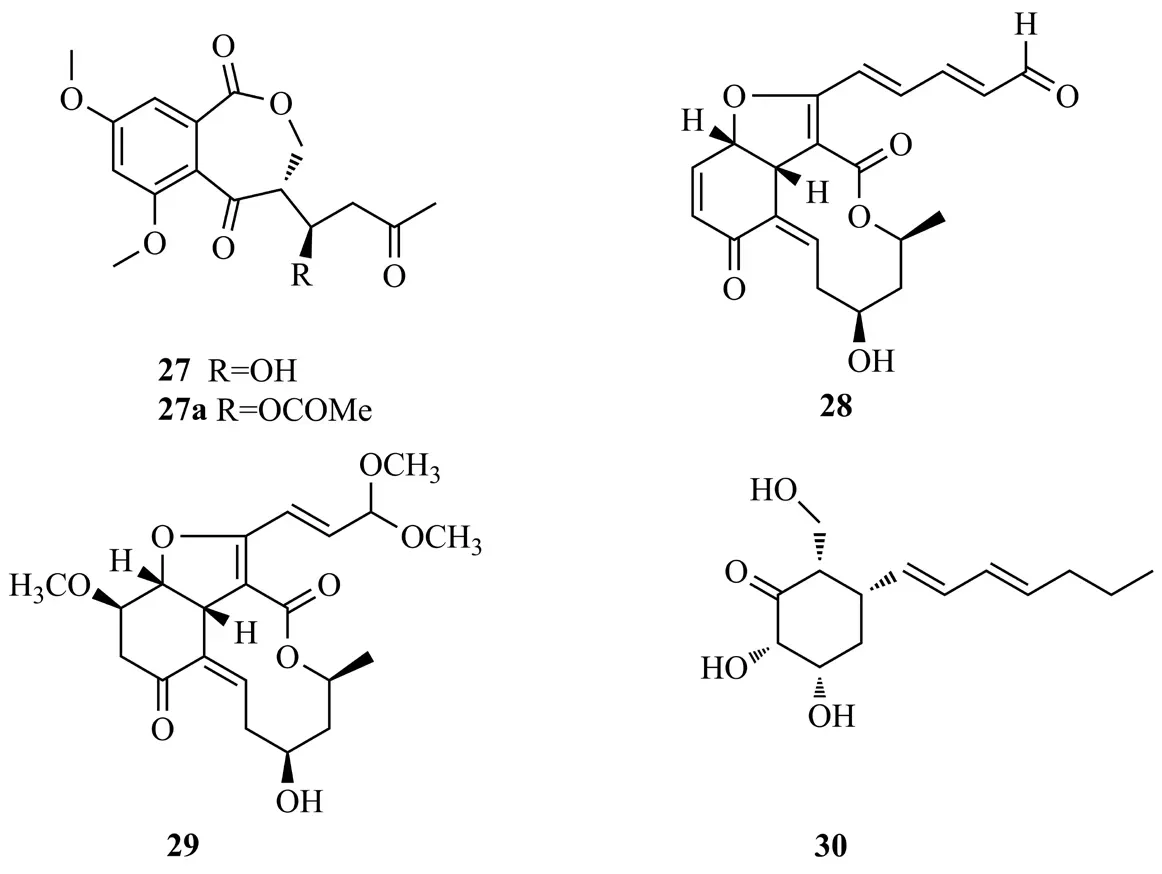
圖5 化合物27~30結構式Fig.5 The structures of compounds 27~30
Luo等[18]從廣西地不容塊根中分離得到青霉菌Penicillium citrinumDBR-9,從其發酵液中分離鑒定出2個抑菌活性的聚酮化合物citrinin(31)和emodin(32);二者對植物病原真菌菌絲生長均有明顯的抑制作用,其IC50值范圍為3.0~141.0 μg/mL;化合物32能夠影響病原真菌的菌落形態,破壞細胞膜完整性而影響所測試真菌細胞的蛋白質合成。Alternates D(33)是從蛇足石杉內生真菌Alternaria alternataMT-47的固體發酵時分離得到的聚酮類化合物,該化合物能夠抑制凝血酶激活血小板的ATP釋放,IC50值為(57.6±3.2) μmol/L[19]。Li等[20]從馬鞭草中分離的真菌Peyronellaeasp.FT431培養液中純化出一種含有苯并異喹啉-9-酮片段的聚酮化合物peyronetide A(34)及其衍生物peyronetide B(35),化合物34和35對人腎腺癌細胞TK-10、順鉑敏感的人卵巢癌細胞A2780S和順鉑耐藥的A2780CisR細胞株具有細胞毒性(IC50值為21.2~29.2 μmol/L),化合物31~35結構式見圖6。
圖6 化合物31~35結構式Fig.6 The structures of compounds 31~35
1.2 萜類
1.2.1 倍半萜
Chen等[21]從三七葉內生真菌LeptosphaeriaXL026的固體發酵培養基中分離出2個倍半萜化合物leptosphins A(36)和B(37),化合物37對蠟樣芽胞桿菌(Bacillus cereus)表現出中等的抗菌活性(MIC值為12.5 μg/mL)。Xu等[22]從三七莖中分離得到內生真菌Preussia isomera,從中分離得到一對罕見的天然存在的倍半萜對映體(+)-和(-)-preuisolactone A(38),對黃色微球菌(Micrococcus luteus)具有抗菌活性(MIC值為10.2 μmol/L)。Pang等[23]從三七葉組織得到另外一株內生真菌Emericellasp.XL 029,從其液體培養液中分離到2個具有三環[4,4,2,1]十一烷骨架的倍半萜emericellins A(39)和B(40),這2個化合物對3種真菌菌株(黃萎病菌Verticillium dahliaeKleb、玉米小斑病菌Helminthosporium maydis和葡萄座腔菌Botryosphaeria dothidea)和3種細菌菌株(枯草芽胞桿菌Bacillus subtilis、蠟樣芽胞桿菌B.cereus、大腸埃希菌E.coli)的MIC值在25~50 μg/mL之間。化合物36~40結構式見圖7。

圖7 化合物36~40結構式Fig.7 The structures of compounds 36~40
Qu等[24]從黃海棠內生真菌Phomopsis prunorum中分離得到2個倍半萜對映體(±)photerterpenes A(41)和B[(±)-1](42),photerterpene A對植物病原菌甜瓜細菌性葉斑病菌(Pseudomonas syringae Pv.lachrymans)具有中等抗菌活性(MIC值為15.6 μg/mL)。Liu等[25]從黃烏頭內生真菌Phomasp.的培養液中得到phomanolide(43),該化合物對A型流感病毒A/Puerto Rico/8/34,H1N1表現出很強的抗病毒活性IC50值(2.96±0.64)μg/mL。Wu等[26]從南方紅豆杉內生真菌Perenniporia tephropora培養液的EtOAc提取物中得到倍半萜perenniporin A(44);從另外1株南方紅豆杉樹皮內生真菌PhomopsisA240的提取物中分離得到倍半萜eremophil-9-ene-11,12,13-triol(45)[27],化合物41~45結構式見圖8。
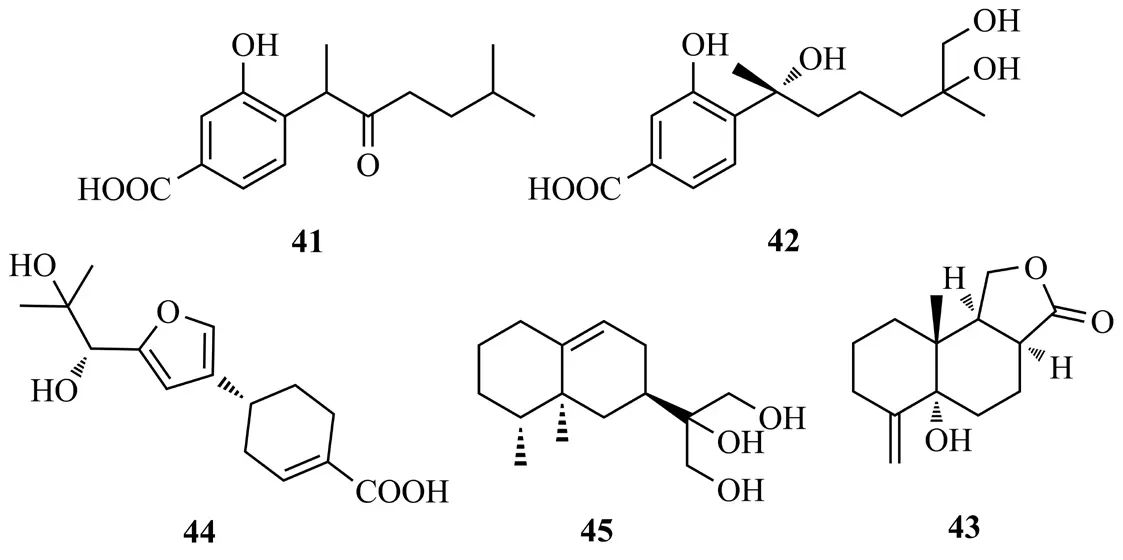
圖8 化合物41~45結構式Fig.8 The structures of compounds 41~45
1.2.2 二萜
Ariantari等[28]對生姜根部分離的內生真菌青霉Penicilliumsp.進行研究,發現吲哚二萜型化合物shearilicine(46),該化合物對L5178Y和A2780細胞系具有較強的細胞毒性(IC50值分別為3.6和8.7 μmol/L)。46是含有6/5/6/6/6/6/5雜環系統的新型吲哚二萜,其結構中的芳香環對于其細胞毒性起到重要的作用。從三七根部分離到真菌Drechmeriasp.,從中也分離得到1個吲哚二萜化合物drechmerin I(47)[29]。從虎杖內生真菌Cercosporasp.的粗提物中分離到1個guanacastane型二萜的異二聚體cercosporenes F(48),該化合物對人源腫瘤細胞系HeLa和T24具有較強的細胞毒性(IC50值分別為19.3和8.16 μmol/L)[30]。
從銀杏樹皮中分離到的一株內生真菌Trichothecium roseum,從中得到一個二萜類代謝產物rosoloctone(49),該化合物對人宮頸癌Hela細胞具有抗增殖與促凋亡作用,它能夠通過激活內質網應激和線粒體損傷來誘導Hela細胞凋亡[31]。從另外一株銀杏內生真菌Aspergillussp.的發酵液中分離到三聯苯基衍生物4′′-deoxycandidusin A(50)和terphenolide(51),它們對神經氨酸酶具有中等強度的抑制活性(IC50值:4.34~9.17 μmol/L)[32]。
從蛇足石杉內生真菌Penicillium chrysogenumMT-12固體基質培養基中分離到二萜penicichrysogenes A(52),該化合物具有抑制凝血酶活化血小板ATP釋放的作用[IC50值為(42.7± 3.5) μmol/L][33]。從火把花的健康花中內生真菌Trichoderma atrovirideB7的液體發酵物中獲得了具有4/7/5/6四環駢合的哈茨烷二萜harzianols I(53),對金黃色葡萄球菌[EC50=(7.7±0.8) μg/mL]、枯草芽胞桿菌[EC50=(7.7±1.0)μg/mL]和黃體微球菌[EC50=(9.9±1.5)μg/mL]均具有顯著的抑菌作用[34]。化合物46~53結構式見圖9。
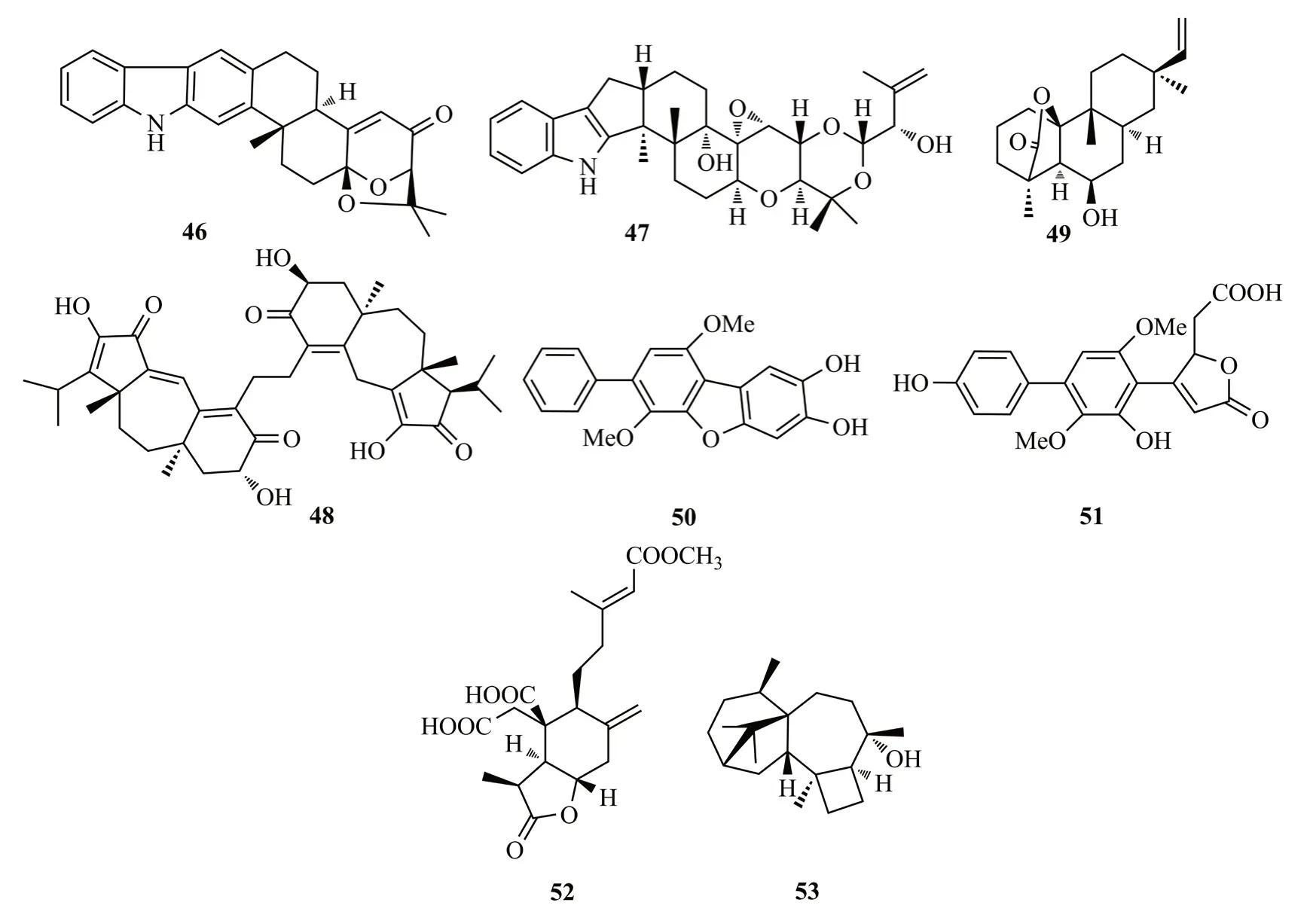
圖9 化合物46~53結構式Fig.9 The structures of compounds 46~53
1.2.3 雜萜
Yang等[35]從石菖蒲內生真菌PhyllosterpictaJ13-2-12Y中分離到雜萜化合物Phyllomeroterpenoids A(54)和C(55),這兩個化合物對金黃色葡萄球菌Staphylococcus aureus209P和白念珠菌Candida albicansFIM709具有一定的抑制活性(IC50值為32~128 μg/mL)。Qin等[36]從南五味子內生青霉菌Penicillium ochrochloronSWUKD4.1850的發酵物中分離得到一系列多氧化取代的萜類化合物,其中kadangusloctones A和C(56~57)對腫瘤細胞HepG2具有中等細胞毒性(IC50值10.2和17.3 μmol/L)。Long等[37]從枸杞內生真菌BipolarisL1-2中分離得到一系列強細胞毒性的cochlioquinones類雜萜,bipolohydroquione B(58)和C(59),cochlioquiones I(60)和K~M(61~63)。58是第一個含有苯并呋喃基團的cochlioquinones類衍生物。59~63對NCI-H226和/或MDA-MB-231具有較強的細胞毒性,IC50值為5.5~9.5 μmol/L。化合物54~63結構式見圖10。
Duan等[38]從天麻內生真菌PenicilliumT2-8中分離到1個雜萜類化合物preastinoid D(64),該化合物對白念珠菌顯示有中等的抑菌活性(MIC值128 μg/mL)。Yan等[39]從檀香樹嫩枝中得到內生真菌FusariumYD-2,并從中獲得了2個螺旋體萜類化合物,即鐮刀菌素A(65)和B(66)。化合物65對金黃色葡萄球菌Staphylococcus aureus和銅綠假單胞菌Pseudomonas aeruginosa表現出顯著的活性(MIC值為6.3 μg/mL)。Bowen 等[40]從蛇足石杉內生真菌Penicillium chrysogenumMT-12的固體底物培養物中分離出雜萜類化合物chrysogenolides C-D(67-68)和F(69)。這3個化合物具有抑制脂多糖誘導的RAW 264.7巨噬細胞釋放NO的作用(IC50值4.3~78.2 μmol/L)。Li等[41]從鉤腺大戟的內生真菌Guignardiasp.固體培養物中分離出雜萜guignardone N(70),評價該化合物在6.3 μg/mL的氟康唑溶液中,對白色念珠菌的生長有顯著的抑制作用,其抑制濃度指數為0.23;在40 μg/mL濃度下與氟康唑聯用,可抑制白色念珠菌生物膜并逆轉其對氟康唑的耐受性。化合物64~70結構式見圖11。
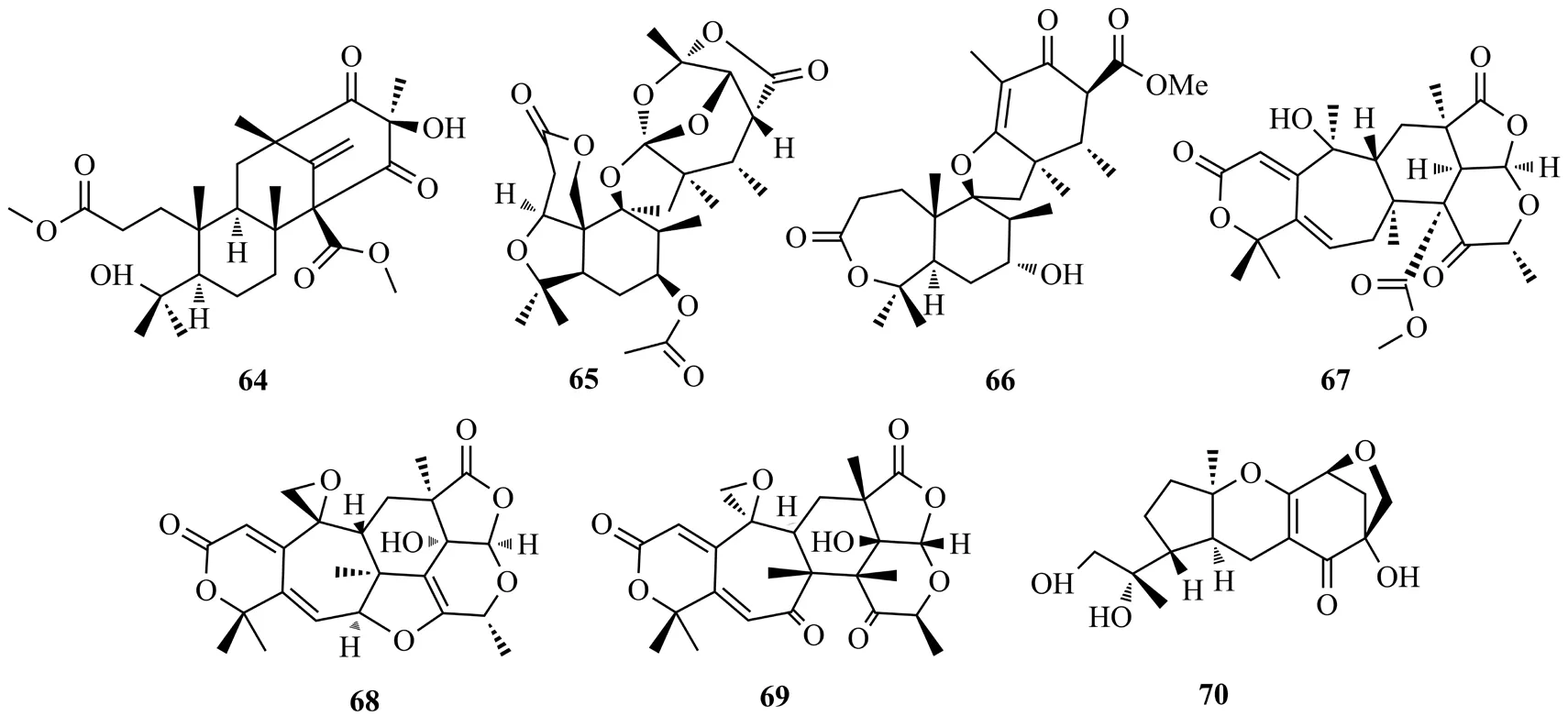
圖11 化合物64~70結構式Fig.11 The structures of compounds 64~70
1.3 生物堿類
1.3.1 哌嗪類生物堿
Zhai等[42]從糙蘇的內生真菌Phaeosphaeria fuckelii中分離得到硫二酮哌嗪生物堿phaeosphaones A~D(71~74),這4個化合物都含有β-O-硫代色氨酸基團,化合物74能夠抑制蘑菇酪氨酸酶的活性[IC50值為(33.2±0.2) μmol/L],比陽性對照藥曲酸IC50=(40.4±0.1) μmol/L活性強。
1.3.2 細胞松弛素
從三七內生真菌Trichoderma gamsii中分離得到細胞松弛素trichalasins A(75)[43]和C(76)[44]、trichoderone A(77)以及aspocharasin D(78);其中,78對HeLa細胞有抑制作用,IC50值為5.72 μmol/L[45]。
1.3.3 吲哚類生物堿
Peng等[46]從三七內生真菌Chaetomiumsp.固體發酵產物中分離出1個吲哚型生物堿chetoseminudin F(79),該化合物對人乳腺癌細胞MDA-MB-231具有強于紫杉醇的細胞毒性(IC50值為26.49 μmol/L)。化合物71~79結構式見圖12。
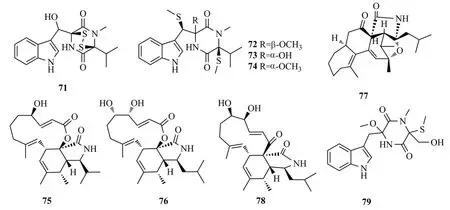
圖12 化合物71~79的結構式Fig.12 The structures of compounds 71~79
1.3.4 其他類生物堿
Sun等[47]從三七的內生鐮孢菌屬(內生鐮孢菌屬)Fusarium tricinctum分離到2個生物堿rigidiusculamide E(80)和[-(α-oxyisohexanoyl-N-methyl-leucyl)2-](81),化合物81對小鼠巨噬細胞NO產生具有顯著的抑制作用,IC50值為(18.10±0.16)μmol/L;從三七另一株內生真菌Preussia isomera中分離出生物堿(±)-Preisomide(82),結構中含有少見的天然四氫-2H-1,2-惡嗪環[48];從霍山石斛的內生真菌Stagonosporopsis oculihominis中分離得到兩個生物堿stagonoculiepine(83)和stagonoculiazin(84),二者對SMMC-7721細胞的抑制率最高,而對A549細胞的抑制率最低(IC50>40mmol)[49]。Zhu等[50]從百山祖冷杉樹皮的內生真菌Penicillium nothofagiP-6中分離鑒定了1個N-甲氧基-1-吡啶酮生物堿chromenopyridin A(85),對A549和Hela人癌細胞株具有較強的細胞毒性(IC50值分別為14.7和11.3 μmol/L);從苦楝的莖皮中得到曲霉菌Aspergillussp.,從中分離到3個吲哚二酮哌嗪生物堿,命名為asperptanthrins A-C(86-88);這些化合物結構中包含由色氨酸殘基和鄰氨基苯甲酸酯形成的色氨酸骨架,aspertypanthrin C含有稀有16元環分子骨架[51]。化合物80~88結構式見圖13。
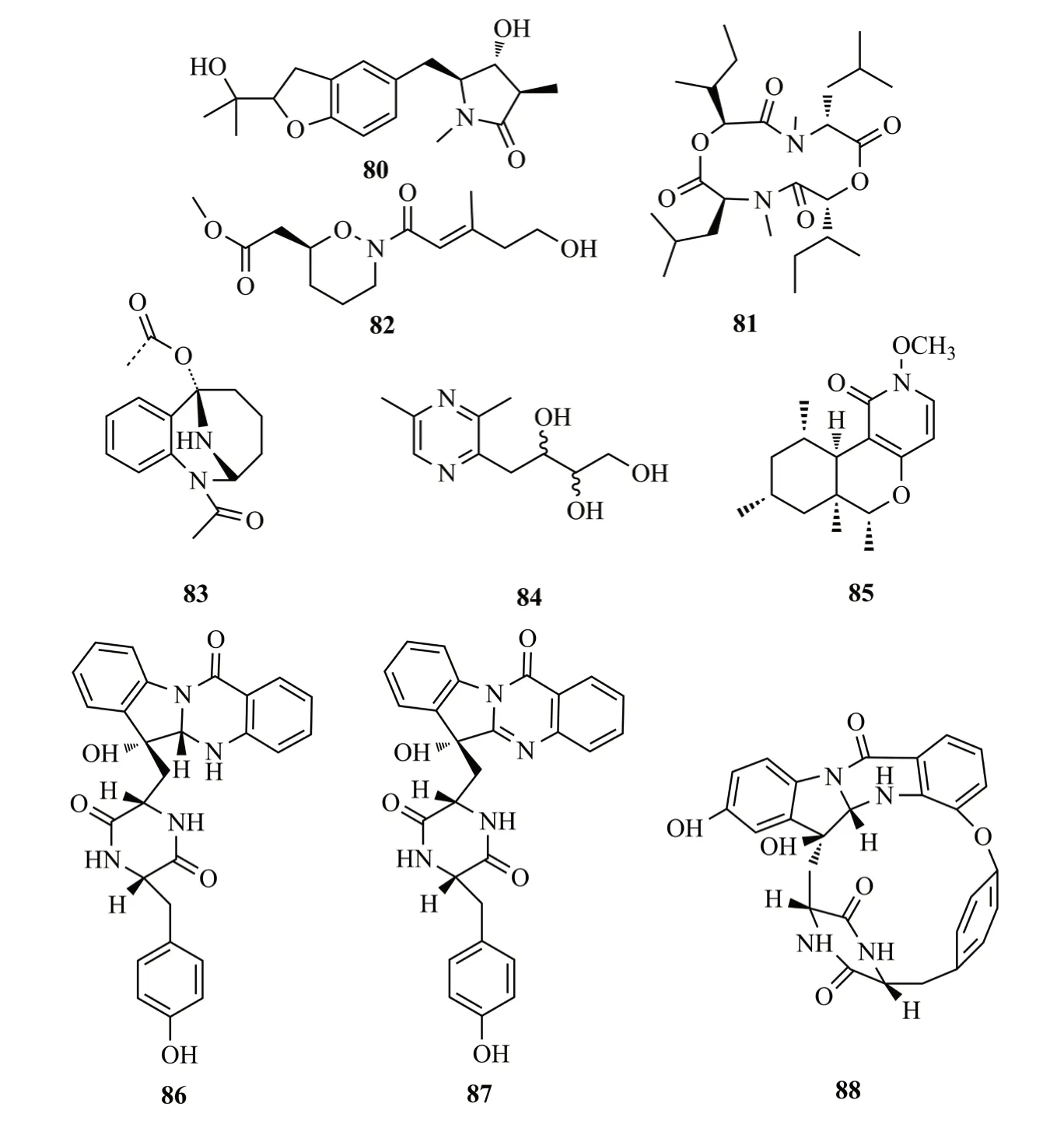
圖13 化合物80 ~88的結構式Fig.13 The structures of compounds 80~88
1.4 其他類化合物
Zheng等[52]從人參的內生青霉菌Penicillium melinii培養液中分離出苯甲醛衍生物ginsenocin(89)和brefeldin A(90);2個化合物對MKN45、LOVO、A549、MDA-MB-435、HepG2和HL-60等腫瘤細胞系表現出很強的細胞毒性(IC50值分別在0.49~7.46 μg/mL之間和IC50值<0.12 μg/mL);從人參Panax ginseng根中分離到另外一株青霉菌PenicilliumSNF123,并從中得到化合物acremonidin E(91);考察acremonidin E的抗色素沉著作用,用α-MSH刺激小鼠黑色素瘤細胞B16F10,用2.5~50 μg/mL的acremonidin E處理48 h后,顯示細胞內黑色素含量顯著降低[53]。化合物89~91結構式見圖14。
Yang等[54]從牛膝莖部的內生真菌PhomopsisCGMCC No.5416的培養物中分離到2個azaphilones類化合物 phomopsones B-C(92-93),二者對HIV-1病毒有顯著的抑制作用,其IC50值分別為7.6和0.5 μmol/L;另外,這2個化合物對A549、MDA-MB-231和PANC-1細胞具有細胞毒性(IC50值在3.2~303 μmol/L);93還能夠誘導PANC-1癌細胞早期凋亡,凋亡率為28.54%。
Xia等[55]從黃瑞香葉片中分離得到PenicilliumpurpurogenumIMM 003,從該菌株的發酵產物中分離得到3個新的含6/4/5/5四環體系的多氧佛手莫烷類化合物purpurolides D~F(94~96),這些化合物的C-14位上3-羥基癸酸部分對提高抑制胰脂肪酶(PL)的能力起到重要作用[如96的IC50值為(1.22±0.08) μmol/L]。從魚腥草葉的內生真菌TubakiaECN-111中分離得到3個異香豆素類化合物,tubakialactones A-C(97-99)[56]。Sabrin等[57]從紅花的根部內生真菌土曲霉Aspergillus terreus中分離得到2個丁內酯aspernolides F(100)和G(101)。化合物92~101結構式見圖14。
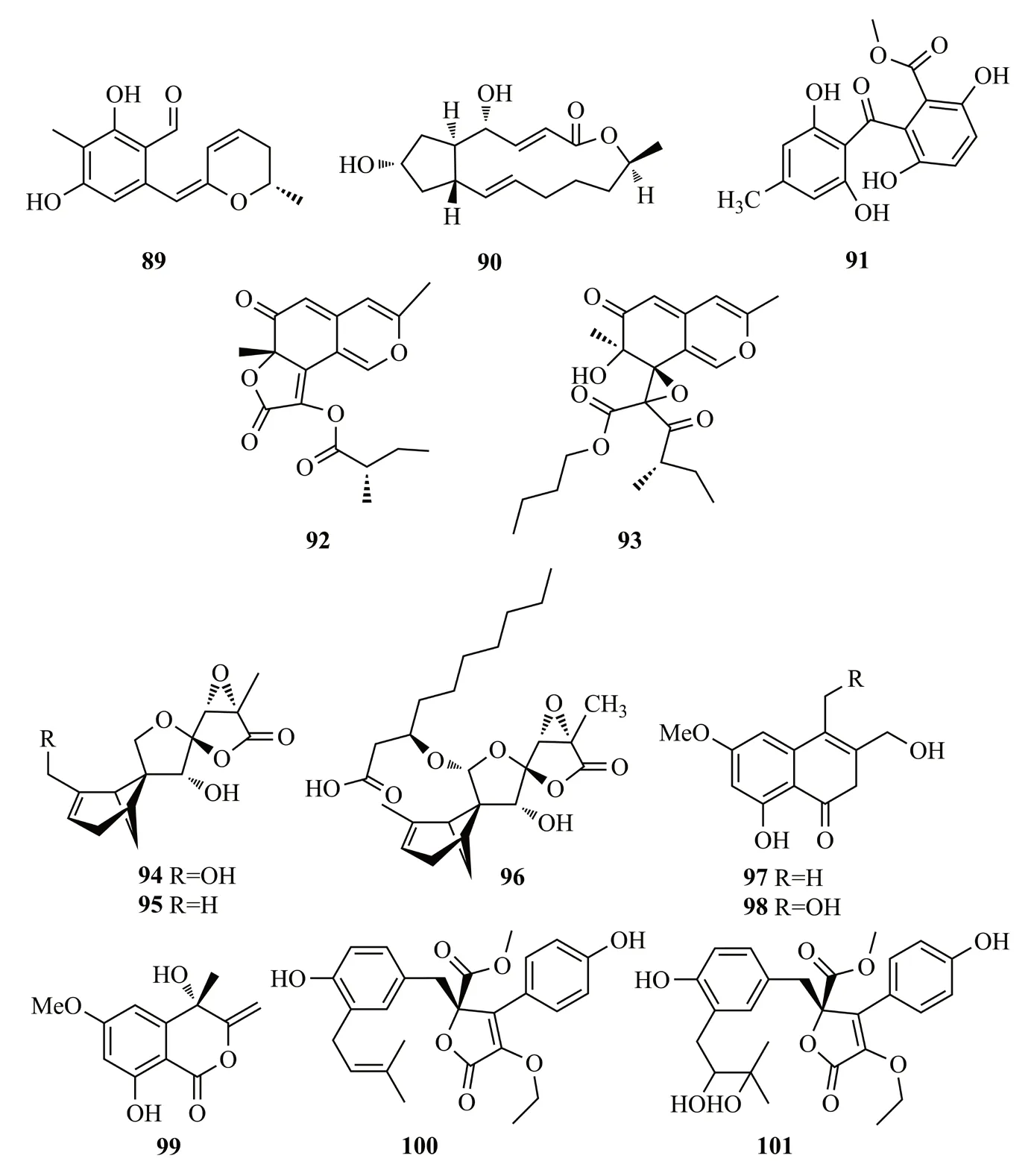
圖14 化合物89~101的結構式Fig.14 The structures of compounds 89~101
2 次級代謝組學、基因組以及蛋白質組學的應用
植物內生真菌次級代謝產物是天然藥物的重要來源之一。當以生物活性為導向的傳統篩選機制出現停滯時,采用新的篩選方法減少已知化合物的重復檢測,快速得到結構新穎且有生物活性的新化合物是藥物開發的關鍵。研究者們對復雜微生物代謝通路進行分析的技術已逐漸成熟,隨著高通量測序技術和生物信息學的發展,能夠通過基因水平和代謝調控定向尋找與實際需求相符的藥物,最終對于天然活性產物的挖掘將會得到更大的發展,次級代謝組學、基因組挖掘技術和蛋白質組學技術已成為尋找新藥源的重要手段。
2.1 次級代謝組學
以LC-HRMS和1H NMR技術作為檢測手段的微生物次級代謝組學能夠對微生物次級代謝產物進行定性和定量分析,已成為研究潛在復雜微生物次級代謝產物譜的重要技術。基于次級代謝組學分析應用于次級代謝產物之間的化學篩選和快速對比,能夠高效揭示一系列次級代謝組間的成分差異,并定位各個實驗組數據中的差異化合物[58-59]。
Tawfike等[60]從桔梗的莖中分離到內生曲霉菌Aspergillus flocculus,在次級代謝組學的輔助下從該曲霉培養物中鑒定出抗腫瘤和抗錐蟲的次級代謝產物。通過主成分分析篩選最佳的內生菌培養條件,再運用多變量數據分析輔助去重處理,有效地注釋了活性小分子化合物,最終活性跟蹤得到7個活性次級代謝產物。其中順-4-羥基甜菜堿、5-羥基甜菜堿、間苯二酚、葡萄毒香豆素A和甜菜堿能夠抑制慢性粒細胞白血病細胞株K562的生長;3-羥基水楊素和間苯二酚分別對引起寄生蟲布魯氏菌的昏睡病具有56%和97%的抑制作用。利用高分辨質譜和核磁共振譜作為次級代謝組學分析工具,Mazlan等[61]優化了兩種內生真菌鐮孢菌屬Fusariumsp.和焦腐病菌Lasiodiplodia theobromae,在不同培養基和生長階段的活性次級代謝產物的產生;通過運用定量分析軟件與數據庫的去重,最終從兩株菌中共得到8個具有萘醌結構的1,4-萘醌類化合物和雙氫異香豆素。其中雙氫異香豆素化合物對布魯氏錐蟲具有顯著的活性(MIC值為0.32~12.5 μmol/L)。使用MS/MS分子網絡技術,根據化學結構和成分相似性(CSCS)指標,通過對草菇(Crptoporus volvatus)培養物和子實體提取物的研究發現,從培養的真菌中發現的化合物也可以從野生蘑菇中分離出來。對子實體提取物進行靶向分離,得到3個未知和6個已知cryptoporic acids衍生物,它們是具有異檸檬酸部分的drimane型倍半萜,Cryptoporic acid T是首次報道的三聚體。其中,化合物5′,6′-cryptoporic acid G dimethyl ester和6′,6′Cryptoporic Acid G Dimethyl Ester對HCT-116細胞具有細胞毒性,IC50分別為4.3和3.6 μmol/L[62]。利用非靶向代謝組學,Albright等[63]利用遺傳或化學的方法檢測了降低組蛋白脫乙酰基酶活性(HDACi)后,模型真菌Aspergillus nidulans代謝的1000多個小分子的變化。通過定量的差異分析,發現幾乎相同數量的化合物被上調和下調了100倍以上。檢測到已知和未知生物合成途徑的產物,并發現構巢曲霉A.nidulans能夠產生氟果糖酰胺,這是蛋白酶體抑制劑,其表達被HDACi誘導約100倍或更多,這項工作為“組學”驅動的天然產物研究的復興提供了動力,成為新藥效團的主要篩選方法。Wei等[64]從蕪青中分離出內生真菌pr10,pr10的代謝提取物在抗腫瘤活性方面具有重要作用,通過代謝組學分析確定了pr10的主要代謝產物,結果表明,pr10的代謝產物富含氨基酸和糖衍生物(如海藻糖),具有抑制A549細胞作用。
2.2 基因組學
基因組挖掘是通過高通量測序技術對生物信息學預測的可能編碼高價值天然產物的基因進行測序,了解基因表達翻譯的控制機制并通過一系列手段將其激活表達,分離純化出相應目的產物后解析結構并對生物活性測試的技術[65]。隨著基因組測序技術的不斷進步,研究人員發現微生物中存在大量常規培養條件下未被表達的“沉默代謝途徑”,新型天然產物的生物合成基因簇往往處于沉默狀態,其編碼的次級代謝產物不表達或產量極低。因此,找到激活這些“沉默途徑”,使其產生更多新型次級代謝產物的方法已成為當前研究的熱點[66]。Cain等[67]對云南紅豆杉內生真菌子囊菌F53進行基因組挖掘,發現35個可能的代謝產物合成基因簇,且其中之一編碼聚酮合酶途徑,與氮雜磷酮生物合成途徑具有同源性。從F53中分離得到了一個新的化合物lijiquinone,并且lij基因簇上編碼的酶負責lijiquinone的合成。Wang等[68]以聚酮-二萜雜合分子為靶點的基因挖掘策略,在滇重樓的內生真菌Aspergillus versicolor0312的基因組中,鑒定出1個新化合物(chevalone E)及其氧化衍生物的生物合成基因簇,該生物合成途徑已成功重建在異源真菌Aspergillus oryzae中。
Wang等[69]對內生真菌Phomopsissp.A123的8個親本原生質體進行基因組改組,以提高化學結構新穎的抗腫瘤天然產物脫乙酰基霉環氧二烯(DAM)的產量;經過兩輪基因組改組,獲得了4株高產水壩菌株G2-119、G2-448、G2-866和G2-919;這4個菌株的產瘤率分別是原菌株A123的243、241、225和275倍,這是首次以原生質體融合中親本類型數目為重點的基因組改組優化研究。Li等[70]采用基因組數據挖掘、系統發育分析和合成生物學快速產物鑒定相結合的方法,從220株擔子菌基因組測序真菌中發現并功能鑒定了4個二萜合成酶;通過基因挖掘技術從擔子菌中發現新的二萜類化合物提供了機會。應用基因組改組技術,篩選出高產丹參酮IIA菌株F-3.4,使丹參酮IIA的產量增加了(387.56±0.02) mg/g,是原始菌株TR21的11.07倍,該菌株可替代植株丹參S.miltiorrhiza生產丹參酮IIA;通過基因組改組可以獲得高產菌株的遺傳基礎,這證明基因組改組可以縮短育種周期并提高誘變性,從而獲得具有良好性狀的菌株,這種有用的方法用于工業菌株的分子育種[71]。Soliman等[72]用含異戊烯基焦磷酸酯(IPP,異戊二烯)和GGPP的萜類前體喂養產紫杉醇的內生真菌ParaconiothyriumSSM001,與對照相比,紫杉醇的產量分別提高了3倍和5倍。Mao等[73]對蘑菇內生真菌叢枝根孢(Calcarisporium arbuscula)進行基因組測序,發現了68個與天然產物生物合成有關的核心基因,并且組蛋白H3脫乙酰酶的失活會導致超過75%的生物合成基因的多效性激活和過度表達。通過鑒定,分離得到10種化合物,其中4種含有新結構,包括環肽阿霉素和阿布霉素、二萜類阿布霉素A等。
2.3 蛋白質組學
了解內生真菌基因組編碼的全部蛋白質的表達、功能和調控,是真菌生物學的一個重要挑戰。這對理解植物-真菌的相互作用,發病機制和真菌的定植是非常寶貴的,而蛋白質組學補充了其他功能基因組學方法[74]。利用組學技術,揭示了植物干旱脅迫反應的分子基礎。Ghaffari等[75]研究了接種印度梨形孢菌(Piriformospora indica)的大麥葉片,在受到水分脅迫時蛋白質組學和代謝組學的變化。結果表明,中度干旱對接種梨狀孢菌的植物,有145種蛋白質的豐度產生了影響,而未接種梨狀孢菌的植物,有104種蛋白質的豐度也發生了變化,而更嚴重的脅迫則分別改變了144種和462種蛋白質的含量。Gao等[76]采用蛋白質雙向電泳結合MALDI-TOF/TOF質譜方法,研究了接種瘤菌根菌屬Epulorhizasp.的福建金線蓮的蛋白組學,共分析了27個差異蛋白點,在數據庫中以綠色植物為檢索范圍,確定了22個差異蛋白,功能大多是植物的信號傳導、代謝調節等,光合作用及物質代謝中的功能蛋白及酶類等也有涉及。
Sulc等[77]報道了曲霉菌Aspergillussp.的蛋白質譜,并且利用基質輔助激光解吸電離飛行時間質譜(MALDI-TOF)技術對孢子進行分析,建立了24株Aspergillus菌株的蛋白質質譜數據庫,這些質譜指紋圖譜可用于不同真菌菌株的分型和鑒定,也可用于在宿主-病原相互作用中尋找新的生物標志物。Noir等[78]采用雙向電泳(2-DE)和質譜聯用技術,對布魯美亞禾本科植物Blumeria graminisf.sp.hordei分析了Bgh分生孢子的蛋白質組;通過在NCBInr-ESTs數據庫中搜索,共鑒定出23種不同的真菌基因產物,其中大多數在碳水化合物、脂質或蛋白質代謝方面具有預測功能。
3 結論與展望
近年來,越來越多的天然產物已在臨床使用,如紫杉醇、鬼臼毒素和喜樹堿等。由于天然產物獲取途徑的局限性,產量較低,極大阻礙了中藥材及天然產物的應用和發展。中藥來源的內生真菌在活性天然產物的開發和獲取方面具有重要價值;利用內生真菌菌株生產具有藥用價值的活性物質,為新藥發現和應用開辟了新途徑。
目前,僅有小部分內生真菌資源被開發,許多資源尚處于研究空白狀態。隨著高通量篩選、組合化學、分子生物學等技術手段的發展,可將高通量測序技術與內生真菌的分離相結合,對內生真菌活性物質的發現、合成機理以及與宿主植物關系的相關生物學的研究,極大地推動了內生真菌的天然產物的發展。
