基因工程中載體概述
2022-10-13 09:54:54盛曹穎
中學生物學 2022年8期
盛曹穎
(吳江高級中學 江蘇蘇州 215200)
在基因工程中,外源基因自身很難進入宿主細胞,且在宿主細胞內不能進行正常擴增、表達,許多載體便應運而生。科學家根據其完成的不同生物學效應,將其分為以下幾種:①克隆載體(cloning vector),又稱一般載體,一般在宿主外不能存活,有能自我復制、分子量小、拷貝數多等特點,主要用于擴增外源DNA片段;② 表達載體(expression vector),一般具有高強度的啟動子和核糖體的結合序列,能高效地產生較穩定的mRNA,主要用于外源DNA在宿主細胞內的高效表達;③穿梭載體(shuttle vector),可用于不同生物間復制。
在這些載體中,克隆載體是最常見的載體,在基因工程中占有十分重要的地位。目前,科學家已構建和應用的克隆載體有數千種。根據其構建所用的DNA來源,克隆載體可分為質粒載體、噬菌體載體、病毒載體和人工染色體載體等。
1 質粒載體
質粒(Plasmid)是人們廣泛應用的克隆載體。它是細菌和真菌等細胞內獨立存在的一種環狀DNA分子。幾乎所有的質粒都帶有一個或多個基因。質粒不影響宿主細胞的生存,但其帶有特殊功能的基因可以賦予宿主細胞某些抵御外界不利環境因素的特性。
1.1 質粒的生物學特性
1.1.1 質粒的復制
幾乎所有的質粒都含有復制起點。質粒侵染宿主細胞后,不同質粒復制的方式不同。根據是否攜帶一套促進DNA轉移相關的基因(稱為基因),質粒分為整合型質粒和非整合型質粒。整合質粒可以整合到宿主細胞染色體的DNA上進行復制,而非整合質粒依賴于自身或宿主細胞體內編碼的酶進行自主復制而實現擴增,具體過程如圖1所示。
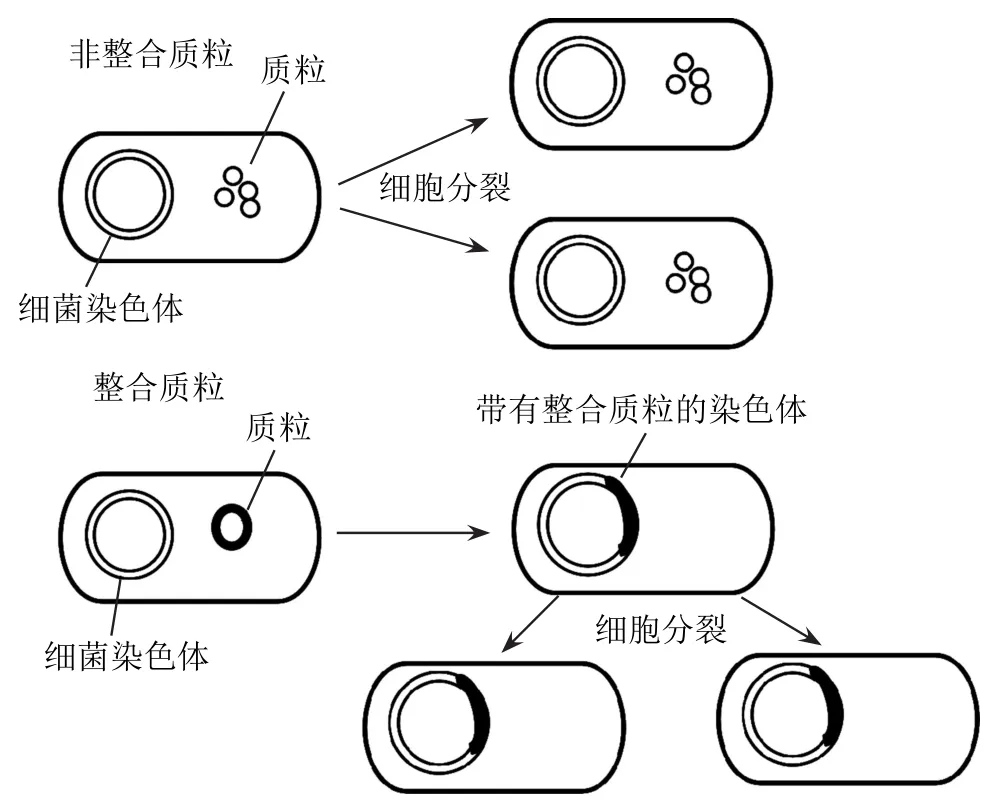
圖1 非整合質粒和整合質粒的復制過程模式圖
1.1.2 質粒的大小和拷貝數
質粒的大小一般在1~250 kb之間。但是,科學家發現一般小于10 kb的質粒才適合用于構建克隆載體,而大的質粒經過一定的改造后亦可用于基因克隆。質粒拷貝數是指在某一生物組中能夠找到質粒的分子數,目前對控制拷貝數的因素尚未了解,但發現每個質粒具有的拷貝數在1~800之間。一般來說,一個可以應用于實踐的克隆載體需要以大量的拷貝形式存在于細胞中,才能夠獲得大量的重組DNA。表1列出了幾種常用質粒的大小與拷貝數的關系。
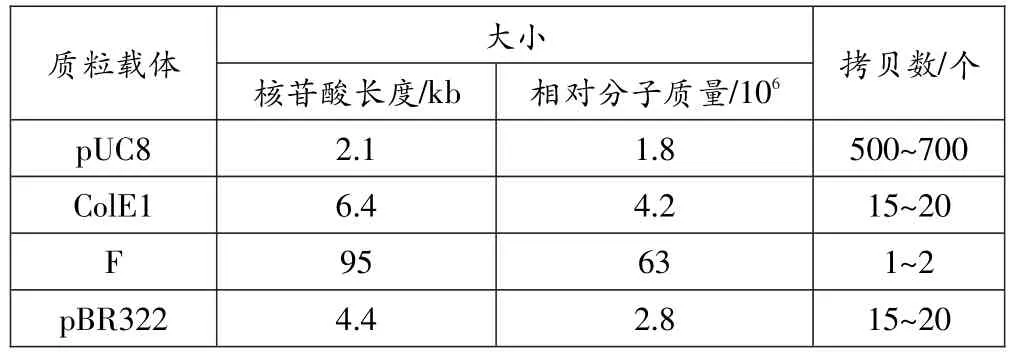
表1 常用質粒載體大小與拷貝數的關系
1.1.3 質粒的不相容性
質粒的不相容性是指利用同一復制系統的兩個質粒在同一宿主中不能共存的現象。其原因主要是在同一細胞中復制時,一般不相容的質粒具有相同的復制子,會存在競爭,從而隨機被淘汰,導致在子細胞中只存在其中一種質粒。
1.1.4 質粒的轉移性
質粒的轉移性是指在自然條件下,質粒通過細菌接合作用可以轉移到新宿主細胞內。有些質粒(如質粒pBR322)本身不能進行結合轉移,但含有轉移起始位點nic,借助于第三個質粒(如ColK)編碼的轉移蛋白的作用,通過結合質粒來進行轉移。基因操作中可以在不同的復制單位上放入轉移必需的因子,通過順反互補來控制目的質粒的接合轉移。
1.2 質粒載體的種類
按照不同的方式,質粒分類不同。質粒的基因所決定的主要特征是天然存在的質粒最主要的分類依據。根據這一分類方法,質粒可分為以下5種:①致育質粒,又稱F質粒,僅攜帶轉移基因,只具有促進質粒間有性接合轉移的特征,如大腸桿菌中的F質粒;②藥性質粒,又稱R質粒,攜帶有能夠賦予宿主細菌對某一種或多種抗菌劑的耐藥性基因,如抗氯霉素、氨芐青霉素或水銀;③Col質粒,編碼一種能夠殺死其他細菌的大腸桿菌素蛋白,如大腸桿菌的ColE1質粒;④降解質粒,使宿主菌能夠代謝一些通常情況下無法利用的甲苯、水楊酸等分子,如惡臭假單胞菌中的TOL質粒;⑤毒性質粒,賦予宿主菌致病性,如根瘤農桿菌中的Ti質粒,能夠在雙子葉植物中誘導冠癭瘤。
1.3 質粒載體的構建
由于野生質粒往往存在許多缺陷,因此目前使用的質粒載體大多是人工構建的。每種質粒的構建思路和過程都十分相似,都是根據基因工程的需要,除去一些非必需元件,添加一些必需的元件,使其成為一種帶有多個單一限制酶切位點、多種強選擇標記、低分子質量、高拷貝、易于轉化、無感染性等優點的理想載體。
1.4 質粒載體的應用
由于這類載體可以在細胞內維持較高的拷貝數,有利于擴增外源DNA,所以具有非常廣泛的應用。例如,早期的pBR系列載體以及現在實驗室常用的Takala公司的pMD18-T載體等。這些載體可以把經過PCR擴增或經聚合酶處理3′末端加A的未知序列的DNA片段直接通過T-A克隆的方法插入載體,為載體構建和未知DNA序列的鑒定提供了更加簡潔的解決方案。
2 噬菌體載體
與質粒相比,噬菌體結構要復雜些,雖然它們均可用于克隆和擴增特定的DNA片段,但是利用噬菌體感染細胞比質粒轉化細胞更為有效,且克隆容量也明顯大于質粒,已成為一種不可或缺的基因載體。
2.1 噬菌體載體的生物學特性
噬菌體是一類細菌病毒的總稱,不同種類的噬菌體顆粒在形態結構上差別很大,其感染性強,依賴于細菌宿主細胞才能進行生長、繁殖。
2.2 噬菌體載體的種類
目前使用的噬菌體載體主要分為:①λ噬菌體衍生物,是最早使用的克隆載體,分為插入型載體和置換型載體兩類。②柯斯質粒,是λ噬菌體cosDNA序列與細菌質粒復制原點DNA序列重組而成,具有兩種載體的優點。③單鏈M13噬菌體,作為單鏈DNA載體,可用于基因定位誘變以及單鏈DNA探針制備。
2.3 噬菌體載體的構建
由于野生型的噬菌體DNA中含有較多的限制位點,不適合作為基因克隆的載體。因此,構建噬菌體載體時需要切除多余的限制位點以及非必需的區段,才能將其改造成適用的克隆載體。不同的噬菌體載體構建方式不同,λ噬菌體衍生物主要集中在消除多余的限制性酶切位點和去除λDNA中一些對其生命周期無關的非必需區段;單鏈M13噬菌體先插入一個大腸桿菌的lac操縱子片段和選擇標記,再加入多克隆位點MCS。
2.4 噬菌體載體的應用
噬菌體載體主要用于建立cDNA文庫,具體過程是先將某種生物的一系列cDNA分子先通過置換或插入的方法與合適的噬菌體載體重組,或不經過包裝直接轉染受體菌細胞,或在體外包裝成噬菌體顆粒后轉導受體菌細胞。這樣所形成的噬菌斑群體就是該生物個體的cDNA基因。除此之外,噬菌體載體還可用于克隆外源目的基因,在生化和遺傳學研究中有著非常廣泛的應用。
3 病毒載體
3.1 病毒載體的生物學特性
病毒復雜而精致的結構可以高效轉染宿主細胞,并在細胞內進行復制和表達,這是作為載體的基礎。在外源基因導入真核受體細胞的各種方法中,最為常用的便是利用病毒載體感染宿主細胞的途徑,不同的病毒載體具有不同的表達特點。目前常用的介導基因轉移的病毒有腺病毒、逆轉錄病毒、慢病毒和痘病毒等。
3.2 病毒載體的種類
病毒表達載體大體可分為病毒顆粒載體(插入外源性DNA后,隨病毒的繁殖進行復制和表達)和改建的病毒DNA混合型載體(不能形成病毒顆粒,重組體在真核細胞內像質粒一樣隨宿主細胞的基因復制而復制)。其中后者是目前廣泛使用的。
3.3 病毒載體的構建
目前,科學家已研究出多種病毒載體,特別是腺病毒載體和逆轉錄病毒載體,其構建過程首先對病毒的基因組進行改造,使之能攜帶相關的病毒元件和外源基因,之后包裝成重組病毒載體,進行侵染。
3.4 病毒載體的應用
由于病毒載體可以介導外源基因在真核細胞中高效表達,可以用于實驗室基礎研究,如建立特定基因瞬時或穩定表達的細胞系,也可以表達治療性基因用于臨床基因治療,如治療Duchenne型肌營養不良癥和B型血友病。
4 人工染色體載體
4.1 人工染色體載體的生物學特性
當克隆基因組非常龐大的人類基因組等真核生物基因組時,往往需要克隆幾百甚至數十萬堿基對的DNA片段。此時,克隆載體需要具備更大的承載量,科學家便建構了人工染色體載體。人工染色體載體具備著絲粒、端粒和復制起始區域三種必需成分,才能保證其能在宿主細胞內穩定地自主復制、分離,并隨細胞分裂平均分配到兩個子細胞中,從而保證人工染色體在宿主細胞中的穩定性。
4.2 人工染色體載體的種類
人工染色體載體含有天然染色體基本功能單位,目前科學家已開發出多種類型的人工染色體載體,實驗室常用酵母人工染色體(YAC)、細菌人工染色體(BAC)、人類人工染色體(HAC)等。
4.3 人工染色體載體的構建
不同種類的人工染色體載體的構建不同,但是大體原理相同,如將細菌接合因子、酵母或人類染色體DNA上的復制區、分配區、穩定區、端粒區與質粒裝配在一起。
4.4 人工染色體載體的應用
人工染色體載體主要應用于基因分離、基因組圖譜制作以及基因組序列分析,并且在基因治療領域有很好的應用前景。例如,基因治療過程中,較大的外源DNA片段和人工染色體重組成功后,通過電穿孔技術導入宿主細胞,重組體在宿主細胞內作為一條獨立的染色體進行復制和分裂,并在宿主細胞中表達出正常功能的蛋白,以治療由于基因缺陷或基因突變導致的遺傳疾病。
