中藥干預足細胞上皮間質轉分化治療糖尿病腎病的研究進展
2022-10-11 12:51:10王婉懿王悅芬
環球中醫藥 2022年10期
王婉懿 王悅芬
糖尿病腎病(diabetic nephropathy,DN)是糖尿病的嚴重微血管并發癥,中國糖尿病患者中20%~40%合并DN,是導致慢性腎臟病(chronic kidney disease,CKD)及終末期腎臟病(end stage renal disease,ESRD)的首要原因[1]。研究表明,足細胞損傷是引起蛋白尿和腎小球硬化的關鍵因素,在DN的發生和發展中起重要作用[2]。足細胞上皮間質轉分化(epithelial to mesenchymal transition,EMT)是指上皮細胞失去其標志性的表型特征,轉化為具有間充質表型細胞的過程,伴隨細胞形態的改變和上皮細胞功能的缺失。主要表現為足細胞標記蛋白如Nephrin、ZO-1、podocin、P-cadherin等表達下調;間充質細胞表型標志物如desmin、α-SMA、FSP-1、纖連蛋白(fibronectin)、I型膠原(collagen I)、N-cadherin、波形蛋白(vimentin)及轉化因子(如Snail、MMP-3、MMP-9)等表達上調[3-4]。現將與糖尿病腎病EMT相關的7條信號通路進行綜述(如圖1)。
1 中藥干預TGF-β信號通路治療DN的機制研究回顧
轉化生長因子-β(transforming growth factor-β,TGF-β)主要依靠以Smads和絲裂原活化蛋白激酶為主的信號通路發揮其生物功能。非活性狀態下,TGF-β1與TGF-β1結合蛋白非共價結合。當受到理化因素刺激時,TGF-β配體、TGF-β II型受體二聚體和TGF-β I型受體二聚體結合形成復合物,對TGF-β-I型受體的GS結構域和絲/蘇氨酸殘基磷酸化,形成TβR-II-TGF-β1-TβR-I異源三聚體,引起下游轉錄因子Smad2和Smad3的磷酸化[5]。磷酸化的Smad2/3與Smad4結合并易位到核中與靶基因結合[6]。Smads的核易位還可上調細胞核內結締組織生長因子(connective tissue growth factor,CTGF)的轉錄,可能是導致機體內所有組織器官纖維化的關鍵因子。
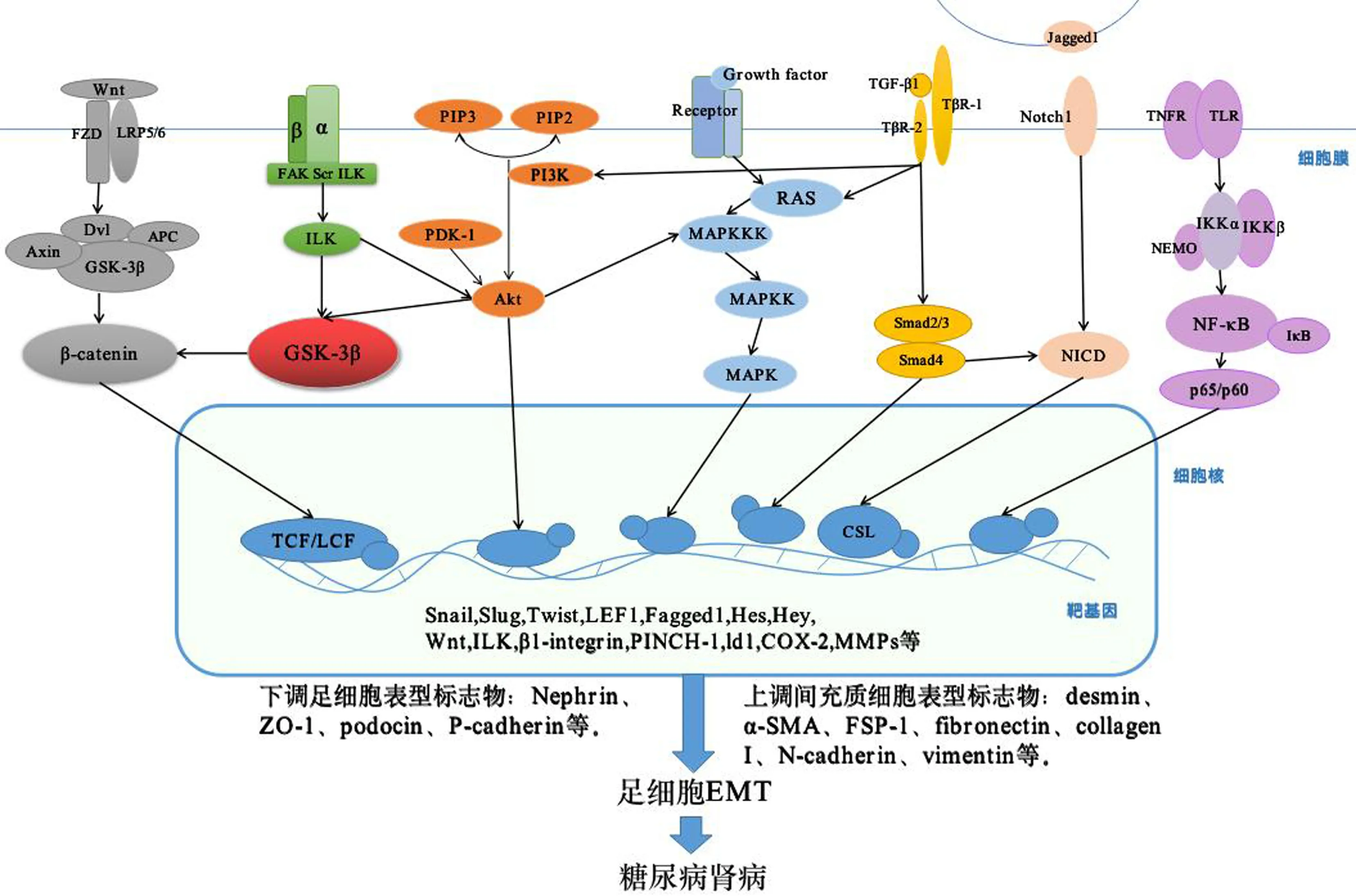
圖1 糖尿病腎病足細胞EMT相關信號通路傳遞過程
研究表明,高糖等刺激因素可引起足細胞TGF-β、Smd2/3水平上調,Smad7表達下調,引起足細胞損傷。經典益氣類中藥黃芪和人參,其主要成分多糖類、皂苷類、黃酮類等在抗氧化、清除自由基和脂質過氧化物等方面有重要作用。Mao等[7]研究發現,黃芪甲苷IV可降低miR-192和TGF-β1、Smad3和肌動蛋白α、I型膠原蛋白等纖維化相關蛋白的表達,而Smad7的表達明顯增加,并呈劑量依賴性地減輕高糖誘導的大鼠系膜細胞增殖,其機制可能是通過TGF-β1/Smad/miR-192通路抑制腎纖維化的產生及系膜細胞的過度增殖。Li等[8]發現黃芪多糖可抑制TGF-β1/Smad信號通路,調控其下游信號分子,升高MMP-2、MMP-9的表達,并降低TIMP-1、TIMP-2的表達,最終可減輕腎臟纖維化損傷。Du等[9]分別使用人參皂苷Rg1、黃芪甲苷IV和兩藥聯用干預糖尿病腎病大鼠,結果顯示,三組都能降低TGF-β1、Smad2/3和CTGF表達水平,增加Smad7的表達,人參皂甙Rg1和黃芪甲苷IV聯用組效果最好,由此發現,人參和黃芪均可通過調控TGF-β信號通路抑制足細胞EMT,保護足細胞,且兩藥協同作用效果更好。茯苓同樣具有益氣健脾,利水滲濕的作用。Wang等[10]通過觀察茯苓酸ZA干預TGF-β1和AngII誘導的HK-2細胞和足細胞,發現PZA通過阻斷Smad2/3-TGF-βRI蛋白相互作用而抑制Smad2/3磷酸化,從而抑制RAS,抑制活化的RAS和TGF-β/Smad信號通路,上調足細胞標志物的蛋白表達,減輕足細胞損傷。大黃是糖尿病腎病的經典用藥,對腎臟的保護作用有效成分主要為大黃素、大黃酸、大黃酚,能明顯改善糖尿病患者腎功能,減輕腎小球高濾過,減少蛋白尿。Guo等[11]發現大黃酚可下調Smad2/Smad3、p53的表達,通過失活TGF-β/Smad信號通路抑制DN的進展。
TGF-β是EMT最經典的信號通路,目前關于DN的研究大多選擇作用于Smad2/3信號靶點,而Smad7蛋白對TGF-β/Smads通路具有負性調控作用,進一步研究中醫藥對于Smad7和CTGF的干預作用同樣具有價值。TGF-β信號通路為新型藥物研發提供了很多思路和方向,如抑制TGF-β1的產生,抑制Smad磷酸化、P38MAPK磷酸化等,以及阻斷TGF-β信號通路對機體是否有其他不良影響尚需探討。
2 中藥干預Wnt/β-catenin信號通路治療DN的機制研究回顧
Wnt/β-catenin信號通路與多種慢性腎臟病、癌癥、骨質疏松等相關[12]。正常情況下,β-catenin與APC、Axin、CK-1、GSK-3β形成結構復合物,被CK-1和GSK-3β磷酸化失活[13]。當足細胞受到高糖或TGF-β等刺激時,胞外Wnt配體可與跨膜蛋白Frizzled受體和共同受體LRP5/6組成的復合受體結合形成二聚體。Wnt-FZD-LRP5/6復合物激活下游散亂蛋白Disheveled,并與支架蛋白Axin結合,抑制GSK-3β磷酸化β-catenin,阻斷了β-catenin降解,蓄積在細胞質中的β-catenin進入細胞核,與TCF/LEF結合,特異地啟動和激活下游靶基因Snail等的轉錄[14-15]。
丹參作為經典的活血化瘀類中藥,其主要活性成分為酚酸類和二萜醌類。Xiang等[16]利用丹參提取物干預高糖/高脂飼料喂養DN模型大鼠后發現,丹參提取物顯著降低了大鼠腎臟組織中Wnt4、β-catenin和TGF-β的高表達,并改善腎小管上皮細胞空泡變性、糖脂代謝紊亂、炎性細胞浸潤等癥狀。劉嬌嬌等[17]研究表明丹參能通過抑制Ang II誘導的系膜細胞Wnt/β-catenin信號途徑活化,使Wnt1、β-catenin、PAI-1表達與FN合成水平明顯下降,從而發揮抗腎小球硬化的作用。 故推斷,丹參可以通過調控Wnt/β-catenin、TGF-β信號通路抑制足細胞EMT過程,改善腎間質纖維化,達到治療DN的目的。同樣機制的還有黃酮類化合物白藜蘆醇,研究發現白藜蘆醇可顯著降低糖尿病腎病大鼠腎組織Wnt4、β-catenin、TGF-β1、Smad2/3 mRNA及蛋白表達[18]。雷公藤具有多種免疫抑制作用和非特異性抗炎作用,是目前中藥治療腎臟疾病最有效的藥物之一。Chang等[19]研究表明雷公藤顯著降低了大鼠腎臟中Wnt-1、β-catenin、NF-κB-p65、GSK-3β和TGF-β1 mRNA和蛋白水平,雷公藤對糖尿病大鼠的腎臟保護作用呈劑量依賴性,與厄貝沙坦治療相比有改善作用。Shi等[20]發現雷公藤甲素可明顯降低HG誘導的足細胞Wnt3α和β-catenin的高蛋白表達水平,抑制Wnt3α/β-catenin信號通路的激活。大黃具有清熱、祛瘀等功效,其主要提取物之一大黃酚已被證明對足細胞EMT有顯著抑制作用[11],其另一提取物大黃酸被證明可下調Wnt1、p-GSK-3β/tGSK-3β、p-β-catenin/tβ-catenin的表達,抑制足細胞EMT[21]。
現階段,對于Wnt/β-catenin通路的上游Wnt、GSK-3β及下游β-catenin靶點作用的研究逐漸增多,對Wnt/β-catenin信號通路的研究多集中在Wnt1和Wnt4蛋白,對其他家族成員的研究相對較少,進一步研究中藥干預Wnt/β-catenin通路的作用可以獲得更有價值的研究結果。
3 中藥干預ILK信號通路治療DN的機制研究回顧
整合素連接激酶(integrin-linked kinase,ILK)作為蛋白激酶時,ILK參與integrin介導的過程,可受TGF-β1信號通路的調節,同時也是TGF-β1信號通路必需的下游信號靶點。當足細胞受到有害刺激時可激活ILK,進一步引起下游AKT、p38 MAPK及GSK-3β磷酸化,從而抑制β-catenin磷酸化。
齊墩果酸是滋陰類中藥女貞子的主要提取物之一,臨床上作為一種廣譜抗菌藥使用,具有抗炎、細胞保護等作用。He等[22]發現齊墩果酸干預可以劑量依賴性的方式逆轉E-cadherin的下調和α-SMA和纖維連接蛋白的上調,還能恢復由TGF-β1啟動的Nrf2和klotho的表達,降低Smad2/3、ILK和Snail的磷酸化。陳皮具有理氣健脾的功效,其有效化合物之一橙皮素有抗炎、抗氧化等藥理作用。Zhang等[23]發現橙皮素可抑制STZ糖尿病腎病小鼠腎組織TGF-β1、ILK、Akt的表達,保護足細胞,延緩DN。益氣活血藥紅景天在臨床中常用于治療DN,主要提取物為紅景天甙。Lu等[24]用紅景天甙干預葡萄糖處理的足細胞,發現紅景天甙提高了p-Akt、p-ILK、p-JNK、p-ERK的表達水平和Nrf-2的定位,顯著減少了ROS的生成和Caspase9/3的表達,紅景天甙通過抑制足細胞EMT和氧化應激以減少足細胞損傷。
目前,中藥調控ILK信號通路主要通過干預ILK及其下游的intergin表達,未對其它具有可調控作用的蛋白、因子進行研究,存在一定的單極化現象,深入研究中藥干預其他信號靶點如GSK-3β的表達,可以開發出干預EMT更為有效而安全的中藥制劑。另外,整合素介導的ECM與細胞和細胞與細胞之間的黏附作用平衡也是中醫藥可進一步挖掘研究的目標。
4 中藥干預Notch信號通路治療DN的機制研究回顧
Notch信號通路與TGF-β通路存在交互協同的作用,是TGF-β1介導足細胞EMT所必需的部分。以往研究認為,Notch通路需要TGF-β通過smad3才能被激活。但近年研究顯示,Notch信號通路更趨向于是一個獨立的信號通路。Notch和Snail2的相互作用,對于Notch介導的E-cadherin或P-cadherin的抑制和β-catenin的激活是必需的。在理化因素的刺激下,Notch配體Jagged1與鄰近受體Notch1的結合,暴露蛋白水解酶的酶切位點,從而發生兩次水解,釋放出Notch受體胞內段,易位至細胞核內。與CSL蛋白結合后,誘導轉錄因子Snail、Hes或Hey與P-cadherin結合,進而使足細胞發生EMT。研究發現,Wnt1的缺失可引起Notch1及其轉錄靶點Nrarp上調,足細胞FoxC2受到抑制,Hey2表達上調[25]。
治療DN的常用清熱類中藥黃芩、黃連,其有效成分黃芩苷、黃連素在腎臟中有抗纖維化的作用。Dou等[26]發現黃芩苷可降低Noch1的活性及其下游Snail的表達,從而抑制Notch1-Snail軸介導的足細胞EMT,從而減輕阿霉素誘導的局灶性節段性腎小球硬化和蛋白尿。Yang等[27]發現黃連素抑制了DN模型KKAy小鼠Notch/Snail通路的激活,上調了α-SMA和E-cadherin水平,降低了jagged1、Notch1、hes1和Snail1的蛋白和mRNA表達水平,保護足細胞。姜黃是治療腎臟病常用的活血藥。祖寧輝等[28]的研究表明用姜黃素干預顯著下調了UUO大鼠的血清BUN、Scr水平、24小時尿蛋白水平及腎組織Notch1、Jagged-1蛋白相對表達量,且實驗組低于模型組。姜黃素可通過調節UUO大鼠腎功能指標的水平,進而改善腎間質纖維化,其作用機制可能與下調腎組織Notch1和Jagged-1蛋白表達,抑制Notch通路的信號轉導相關。
目前中藥作用于Notch信號通路主要通過抑制Notch及其配體Jagged的水解激活實現。現代醫學對Notch通路介導的關鍵蛋白和非編碼RNA的研究進展頗大,中醫藥可以此為靶點深入進行研究,進一步了解中醫藥在治療DN中的機制。
5 中藥干預MAPK信號通路治療DN的機制研究回顧
絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)有p38絲裂原活化蛋白激酶(p38MAPK)和c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)兩個亞類。TGF-β活化后可激活MAPKKK、MAPKKK磷酸化并激活MKK3/4/6,從而雙重磷酸化p38MAPK和JNK,其中MKK3/6可特異性激活p38,MKK3/6也可以在凋亡因子的刺激下被凋亡信號調節酶1直接激活。p38MAPK和TGF-β1起協同作用,一方面p38 MAPK可增強TGF-β1的活性;另一方面,TGF-β1可以使p38MAPK自磷酸化,并激活其下游轉錄因子環狀單磷酸腺苷結合蛋白[29]。
常用于治療DN的經典藥對地黃和山茱萸中,地黃滋陰以補為主,山茱萸斂陰以收為主,合用可益腎強精,大補元氣。Chen等[30]通過使用治療DN的藥對地黃和山茱萸的主要活性成分梓醇和馬黃酮調控晚期糖基化終末產物誘導的KK-Ay小鼠自發DN和足細胞損傷模型,發現梓醇和馬黃酮單獨或聯合抑制足細胞RAGE/p38 MAPK/p65 NF-κB和RAGE/Nox4/p65 NF-κB通路的激活。聯合用藥的抑制作用比單獨用藥的抑制作用更明顯。先前研究表明,袪風濕藥雷公藤的另一提取物雷公藤紅素有強抗氧化和抗炎作用[31]。Zhang等[32]發現雷公藤紅素可降低糖尿病大鼠血肌酐和尿素氮含量,減少尿蛋白排泄,改善腎臟病理損傷,下調p38MAPK和NF-κB p65的表達。丹酚酸B作為活血化瘀藥丹參的多酚酸類主要活性成分,具有調控TGF-β1/Smad信號通路和活化p38MAPK的作用,抑制高糖誘導的腎臟細胞表型轉化,減少Col I和Col III等細胞外基質分泌,減輕炎癥反應[33]。
現階段發現在動物模型中MAPK相關蛋白激酶抑制劑對足細胞EMT有一定的抑制作用,但由于其臨床耐受性及其毒副作用等進一步研究受限。中藥治療多靶點、多環節、不良反應少,因此需深入研究中藥干預MAPK信號通路,抑制足細胞EMT。
6 中藥干預NF-κB信號通路治療DN的機制研究回顧
核因子κB(nuclear factor kappa-B,NF-κB)在細胞質中與抑制性IκB結合在一起處于穩定狀態。理化因素刺激下,IKK復合體(NEMO、IKKα、IKKβ)被激活,促進IκB磷酸化和泛素化水解后釋放出p65/p50并移位至胞核中,與靶基因結合位點即啟動子或增強子結合誘導COX-2、MMPs、VEGF,以及最重要的轉移誘導物Snail等轉錄,促進足細胞EMT[34]。
經典補氣類中藥黃芪和黃芪主要活性成分分別為黃芪甲苷IV和當歸多糖。Wang等[35]觀察到在高糖誘導的小鼠永生化足細胞中,黃芪甲苷IV可抑制TGF-β1表達,并可激活SIRT1-NF-κB通路,抑制高糖誘導的足細胞EMT。當歸多糖[36]可下調糖尿病腎病大鼠腎組織中NF-κB mRNA和蛋白表達水平。化痰止咳平喘藥羅漢果的一種新型多糖提取物(SPG-1-1)可下調G蛋白偶聯細胞膜受體TLR4及其下游蛋白激酶(NF-κB p65)的mRNA和蛋白表達。通過調節炎癥,抑制TLR4-NF-κB通路,刺激超氧化物歧化酶的產生,減少細胞因子(IL-6、TNF-α)和丙二醛的產生[37]。
目前,關于NF-κB信號通路的研究多基于炎癥反應干預,對其參與足細胞EMT的研究較為不足。研究中藥對其相關受體、信號轉導、負反饋調節等都需要更深入的研究。
7 中藥干預PI3K/Akt信號通路治療DN的機制研究回顧
磷脂酰肌醇-3-激酶/蛋白激酶B(phosphatidylinositol 3-hydroxy kinase/protein kinase B,PI3K/Akt)對機體的糖脂代謝有顯著的調節作用。理化因素刺激下,活化的PI3K可特異性地將磷脂酰肌醇2磷酸(PIP2)催化為PIP3,PIP3協同磷酸肌醇依賴的激酶-1(PDK-1)可誘導Akt發生三級結構構象改變并聚集到胞膜,Akt的Thr308及Ser473位點被磷酸化。完全活化的Akt將信號傳導至mTOR,從而激活下游相關信號因子等。PTEN可將PIP2重新催化為PIP3,從而抑制PI3K/Akt信號通路。
清熱藥黃連提取物黃連素是抑菌抗炎的主要有效成分。劉青等[38]發現黃連素能明顯升高HG誘導的足細胞nephrin、WT-1蛋白的表達,降低PI3K、p-Akt、Bim蛋白的表達,明顯升高p-FOXO1蛋白的表達。Wang等[39]研究發現黃連素可以減輕來自高糖誘導腎小球系膜細胞外泌體所導致的足細胞的損傷,其機制可能與TGF-β1以及PI3K/Akt信號通路相關。姜黃具有活血行氣的功效,有效成分姜黃素具有較好的抗炎和抗癌特性。Tu[40]等發現姜黃素可上調DN大鼠和MPC5細胞E-cadherin和LC3蛋白的表達,下調Vimentin、Twist1、p62、p-mTOR、p-Akt和P13K的表達。既往研究已證明,袪風濕藥雷公藤提取物雷公藤紅素能抑制MAPK和NF-κB信號通路主要信號因子的表達。Nie等[41]發現大劑量雷公藤紅素[1.5 mg/(kg·d)]上調了LC3II和nephrin的表達,下調PI3K、p-AKT mRNA、NF-κB和mTOR mRNA的表達。雷公藤紅素通過抑制PI3K/Akt信號通路,下調NF-κB的表達,抑制GBM增厚,并減少mTOR的表達,保護足細胞,減輕糖尿病腎病腎損害。
PI3K/Akt通路被認為是經典的信號通路之一,與多條通路交叉影響。國外關于藥物機制的研究大部分已涉及到非編碼RNA、外泌體等深入分子機制層面,目前國內中藥研究較少,后期無進一步的深入研究。因此發揮中醫藥優勢,深入探索中醫藥干預PI3K/Akt及其交叉通路的分子機制十分必要。
8 小結與展望
通過對相關中藥證型進行整理分析,發現在調控足細胞EMT的中藥單體和單味中藥中,滋陰類中藥地黃、女貞子等;益氣類中藥黃芪、人參、茯苓等;補腎類中藥山茱萸、女貞子、地黃等;活血化瘀類中藥當歸、姜黃、丹參、紅景天等;清熱類中藥黃芩、黃連、生地黃、大黃等,都分別對DN足細胞EMT不同信號通路靶點有阻斷、抑制的作用。對上述藥物分析后發現,靶向干預足細胞EMT信號通路防治DN的中藥大多具有益氣養陰、清熱祛濕、活血化瘀的功用。
目前關于中藥干預足細胞EMT的研究仍存在問題:首先,目前的足細胞EMT信號通路研究多為“有效成分—單一通路”形式,但臨床上中藥單體多數可以同時作用于多靶點調控信號通路,如雷公藤可調控Wnt/β-catenin、MAPK、NF-κB、PI3K/Akt等信號通路抑制足細胞EMT。因此未來需深入研究中藥調控多個信號通路間的聯系及其協同作用以抑制足細胞EMT的具體機制;其次,目前對于中藥調控足細胞EMT的研究大多是從藥理學角度進行研究,缺乏中醫證候學模型實驗及臨床試驗研究,在臨床中根據中醫辨證指導用藥可以使中藥及其有效成分產生更大的價值。因此在下一步的研究中,應當考慮將DN辨證分型和中藥性味歸經、功效主治等方面相結合進行研究,為中藥治療DN提供有力的客觀依據。
本課題組前期研究發現,AGEs-RAGE軸為DN伏邪論提供了證據,內生伏邪與肺脾腎和三焦有關,導致濕痰瘀互結之邪伏于腎之膜原[42]。此過程與足細胞EMT活化,ECM沉積,腎小球基底膜增厚,最終腎纖維化的西醫病理過程相似,本課題組試提出“伏邪—EMT—腎纖維化”相關性假說。基于此假說自擬的保腎方被證明能顯著抑制高糖培養小鼠PI3K/Akt信號通路激活[43],因此伏邪及足細胞EMT相關信號因子可作為未來治療靶點,以期為DN的中西醫治療提供可靠的理論依據。
注:本文出現的英文簡寫之中文全稱對應如下:
P-鈣黏蛋白(P-cadherin)
N-鈣黏蛋白(N-cadherin)
纖維連接蛋白(fibronectin,FN)
I型膠原(collagen I,Col I)
α-平滑肌肌動蛋白(α-smooth muscle actin,α-SMA)
基質金屬蛋白酶3(matrix metalloproteinase3,MMP3)
基質金屬蛋白酶9(matrix metalloproteinase 9,MMP9)
轉化生長因子-β(transforming growth factor-β,TGF-β)
絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)
結締組織生長因子(connective tissue growth factor,CTGF
酪蛋白激酶1(casein kinase 1,CK1)
糖原合成酶激酶3β(glycogen synthase kinase 3β,GSK-3β)
卷曲蛋白受體(frizzled,FZD)
脂蛋白受體相關蛋白5/6(lipoprotein receptor-related protein5/6,LRP5/6)
T細胞因子/淋巴樣增強因子(T-cell factor/lymphoid enhancing factor,TCF/LEF)
纖溶酶原激活物抑制劑-1(plasminogen activator inhibitor 1,PAI-1)
整合素連接激酶(integrin-linked kinase,ILK)
核因子-E2相關因子2(nuclearfactor erythroidderived 2-like 2,Nrf2)
半胱氨酸蛋白酶9/3(caspase9/3)
Notch受體胞內段(notch intracellular domain,NICD)
叉頭框C2(forkhead box c2,FOXC2)
p38絲裂原活化蛋白激酶(p38 mitogen-activated protein kinase,p38 MAPK)
c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)
MAPK激酶激酶(MAP kinase kinase kinase,MKKK)
MAPK激酶(MAP kinase kinase,MKK)
晚期糖基化終末產物(advanced glycation endproducts,AGEs)
晚期糖基化終末產物受體(receptor of advanced glycation endproducts,RAGE)
核因子κB(nuclear factor kappa-B,NF-κB)
抑制性IκB(inhibitor of NF-κB)
kappaB抑制因子激酶(inhibitor of kappa B kinase,IKK)
環氧化酶-2(cyclooxygenase-2,COX-2)
基質金屬蛋白酶(matrix metalloproteinases,MMPs)
血管內皮生長因子(vascular endothelial growth factor,VEGF)
沉默信號調節因子1(silent information regulator 1,SIRT1)
Toll樣受體4(toll like receptor 4,TLR4)
超氧化物歧化酶(superoxide dismutase,SOD)
丙二醛(malondialdehyde,MDA)
磷脂酰肌醇-3-激酶/蛋白激酶B(phosphatidylinositol 3-hydroxy kinase/protein kinase B,PI3K/Akt)
磷酸肌醇依賴性蛋白激酶-1(phosphoinositide dependent protein kinase 1,PDK-1)
哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)
第10號染色體同源丟失性磷酸酶張力蛋白(phosphatase and tensin homology deleted on chromosome 10,PTEN)
Bcl-2蛋白家族促凋亡(Bcl-2 interacting mediator of cell death,Bim)
磷酸化叉頭框蛋白O1(recombinant forkhead box protein O1,FOXO1)
微管相關蛋白1輕鏈3(microtubule-associated protein 1 light chain 3,LC3)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年4期)2021-12-01 11:19:40
中老年保健(2021年4期)2021-08-22 07:08:32
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
金橋(2020年7期)2020-08-13 03:07:00
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
基層中醫藥(2018年6期)2018-08-29 01:20:20
