乙型肝炎病毒/S型區變異特征及其與乙肝疫苗免疫逃逸的研究
2022-10-08 01:33:06賀曉燁李雯莉高冉冉
河北醫學 2022年9期
曹 陽, 賀曉燁, 李雯莉, 高冉冉, 王 瑩
(1.新疆醫科大學第一附屬醫院, 新疆 烏魯木齊 830000 2.云南省昆明市婦幼健康服務中心基層指導科, 云南 昆 明 650000)
隨著乙型肝炎疫苗(hepatitis B vaccine,HepB)應用范圍推廣,免疫失敗問題已逐漸引起廣泛關注,其機制可能與疫苗形成的免疫壓力導致HBV基因突變,進而造成免疫逃逸存在密切聯系[1]。目前已經證實乙型肝炎表面抗原(hepatitis B surface antigen,HBsAg)由大、中和小包膜蛋白組成,位于氨基酸99-169序列區域的高度親水性結構域稱主要親水區(major hydrophilic region,MHR),其中包含的a決定簇(aa124-147)是誘導B細胞產生抗體和免疫應答的重要抗原表位,同時也是疫苗和抗病毒藥物作用靶點[2,3]。目前所用HepB為HBsAg重組蛋白,誘導產生的免疫應答主要針對S基因編碼的MHR區域,因此S基因突變,尤其MHR中的a決定簇突變容易造成疫苗免疫失敗,導致乙肝疫苗免疫逃避病毒(vaccine escaped HBV,ve-HBV)出現[4]。本研究以1973例接種HepB的人群為樣本,分析免疫逃逸發生情況及其與HBV S區基因突變的關系,以進一步明確免疫失敗的遺傳學機制,現將具體結果詳述如下。
1 資料與方法
1.1一般資料:選取2021年3月至2022年3月我院1973例住院患者為樣本進行橫斷面研究,其中男性1005例,女性968例,年齡3~28歲,平均(17.29±5.46)歲。納入標準:①均按照衛生部《預防接種工作規范》[5]中規定的0、1、6月標準程序完成HepB接種;②均采集血液樣本并完成HBV感染標志物檢測;③患者或家屬均了解本研究內容并簽署同意書。排除標準:①合并手術或輸血相關病史;②合并甲型肝炎病毒(HAV)、丙型肝炎病毒(HCV)或人類免疫缺陷病毒(HIV)等其它病毒感染;③合并免疫系統疾病;④曾接受抗病毒藥物治療;⑤臨床信息不完整。
1.2研究方法
1.2.1HBV血清標志物檢測:采集所有研究對象血液樣本3mL,以8000r/min離心后3min,取上清100μL并采用Access2全自動微粒子化學發光免疫分析儀(美國Beckman Coulter公司)及配套試劑盒檢測HBsAg、抗-HBs、HBeAg、抗-HBe和抗-HBc,所有步驟均嚴格按照說明書要求完成。
1.2.2HBV DNA提取:取HBsAg陽性患者血清200μL于離心管中,然后加入20μL蛋白酶K(德國Qiagen公司),震蕩15s以充分混勻,然后于56℃環境孵育10min,完成后加入無水乙醇200μL,搖勻后轉移至收集管離心柱中,在室溫條件下以8000r/min離心1min,更換清潔收集管并向離心柱加入500μL AW1洗液,在室溫條件下以8000r/min離心1min,去濾液并更換收集管,然后向離心柱加入500μL AW2洗液,在室溫條件下以16000r/min離心3min并去除濾液,將離心柱置于1.5mL無菌EP管中,加入30μL AE洗脫液并在室溫條件下以8000r/min離心1min,將所得DNA溶液-70℃保存備用。
1.2.3HBV DNA擴增:采用聚合酶鏈反應(polymerase chain reaction,PCR)技術進行擴增,第一輪反應體系包括10 mM dNTP 2μL,模板DNA 2μL,上、下游引物各1μL,Taq DNA聚合酶體系12.5μL、10×PCR buffer 5μL及雙蒸水26.5μL,總體積50μL。反應條件為98℃預變性10min,然后98℃變性10s,60℃退火15s,72℃延伸1min,循環35次,最后72℃延伸10min。引物由上海生工生物科技有限公司合成,上、游序列分別為5’-ATCCGCTGGATGTGTCTGCG G-3’,下游5’-GGCAACGGGGTAAAGGTTCA-3’。第二輪反應體系包括第一輪PCR產物5μL,上、下游引物各2μL,Premix Taq DNA聚合酶25μL,雙蒸水16μL,總體積為50μL。反應條件為98℃預變性10min,然后98℃變性10s,60℃退火15s,72℃延伸1min,循環35次,最后72℃延伸10min。引物由上海生工生物科技有限公司合成,上、游序列分別為5’-TTAGGG TTTAAATGTA TACCC-3’,下游5’-CATCTTCTTGTTGGTTCTTCTG-3’。將PCR產物采用1.5%瓊脂糖凝膠進行電泳,電壓100V,時間40min,采用紫外多功能凝膠成像儀(法國Vilber Lourmat公司)觀察目標條帶。
1.2.4HBV S基因型和血清型分析:取凝膠電泳產物置于EP管中,封口后送交上海生工生物科技有限公司測序,將測序結果整理后與NCBI基因庫(http://www.ncbi.nlm.nih.gov)中標準序列(見表1)進行對比和分析,采用MEGA4軟件構建系統進化樹并鑒定基因型,HBV S基因型判斷依據為與參考序列S基因同源性≥96%為同一基因型。采用MegAlign軟件分析HBV S基因序列,比對并判斷突變發生情況。然后分析S基因中特定位點氨基酸表達情況確定血清型,HBV S共包括adr、adw、ayr和ayw等4種血清型。
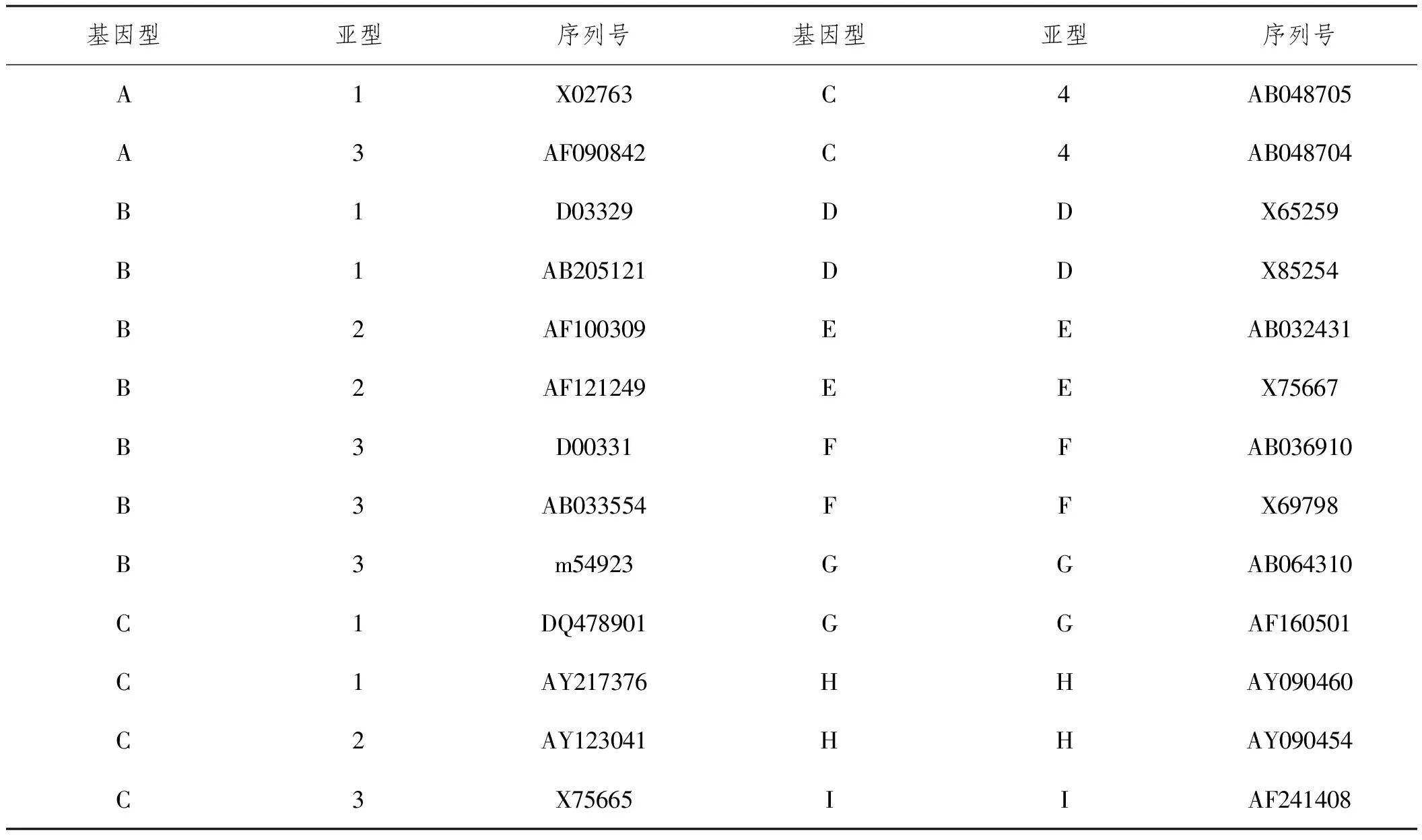
表1 HBV S基因型標準參考序列
2 結 果
2.1HepB疫苗免疫逃逸患者流行病學分布:本研究1973例患者中HBsAg陽性標本47份(2.38%),其中HBsAg/抗-HBc陽性標本26份,占55.32%,HBsAg/抗-HBs/抗-HBc陽性標本3份,占6.38%,HBsAg且抗-HBs/抗-HBc陰性標本18份,占38.30%。年齡>18歲者HBsAg陽性率高于1-18歲人群,父母HBV陰性者HBsAg陽性率低于母親HBV陽性人群,差異均有統計學意義(P<0.05),流行病學分布特征詳見表2。

表2 HepB疫苗免疫逃逸患者流行病學分布
2.2HBV基因型和血清型分析:47份HBsAg陽性標本中HBV基因型包括B型14例(29.79%),C型31例(65.96%)和D型2例(4.26%);血清型包括adr型23例(48.94%),adw型14例(29.79%)、ayr型8例(17.02%)和ayw型2例(4.26%),見表3。
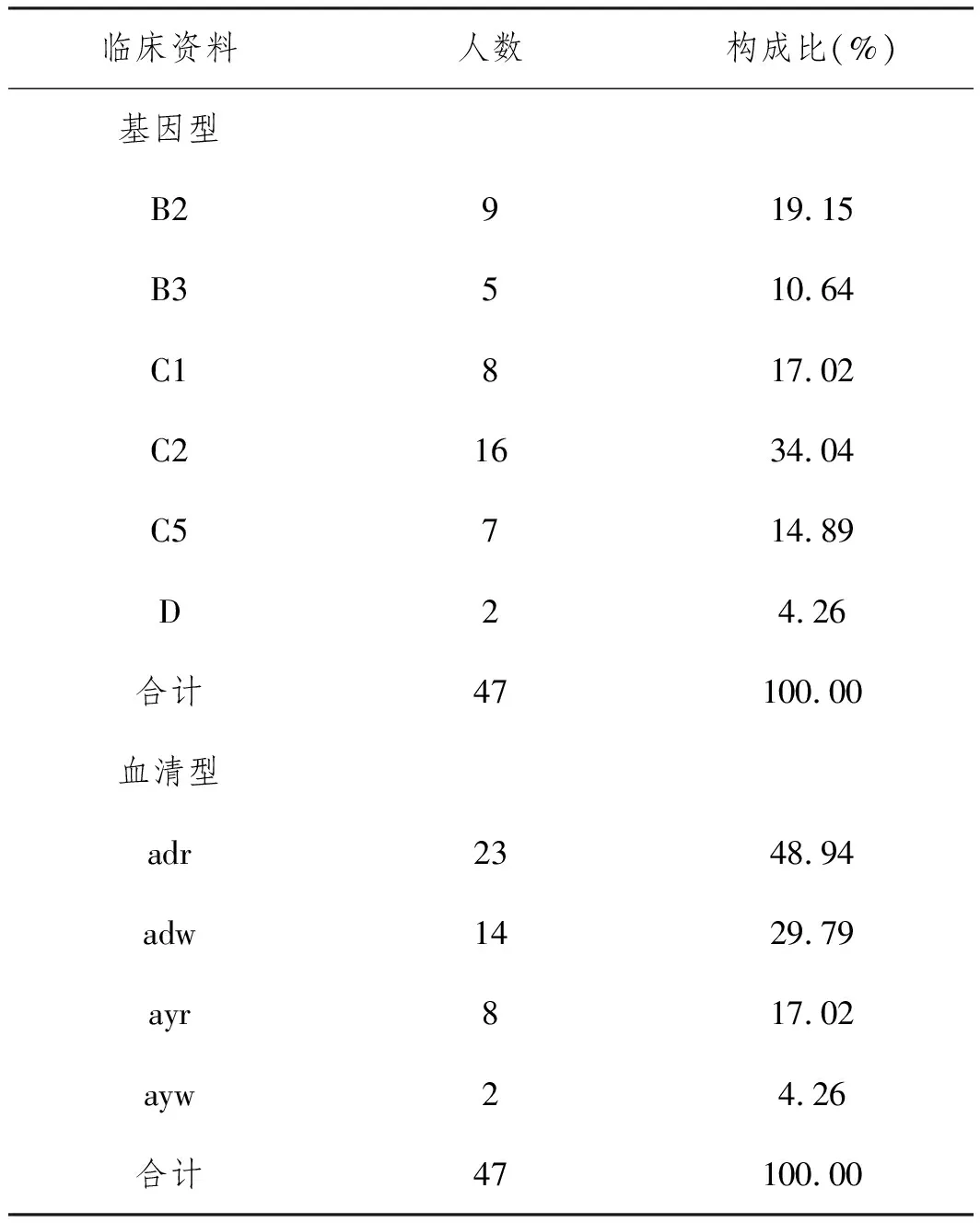
表3 HBV基因型和血清型分析
2.3HBV S基因突變位點數量分析:與標準序列相比,47份HBsAg陽性標本共存在核苷酸突變位點162個,其中突變位點數量1-13個,平均(3.45±1.06)個,突變位點眾數為3個。47份標本共存在氨基酸突變位點98個,其中突變位點數量0-6個,平均(2.09±68)個,30份HBsAg陽性標本檢出氨基酸變異,檢出率63.83%,見圖1。
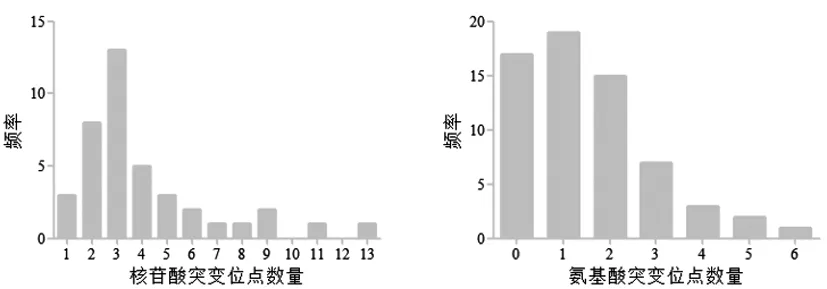
圖1 HBsAg陽性標核苷酸和氨基酸突變情況分析
2.4HBV S區域核苷酸變異情況分析:基因測序顯示HBV S基因突變主要發生于MHR,47份標本中a決定簇(aa124-147)內檢出突變位點包括I/T126A/N/I/S、Q129H/R、M133L、K141E、T143S、D144A/EH和G145R/A;a決定簇上游(aa99-123)檢出突變位點包括Q101R/H、L110I、G112K、S113T/G、S117G和P120Q/R/L;a決定簇下游(aa148-169)檢出突變位點包括Y161F/C、E164D、F161Y和R160K,詳見表4。
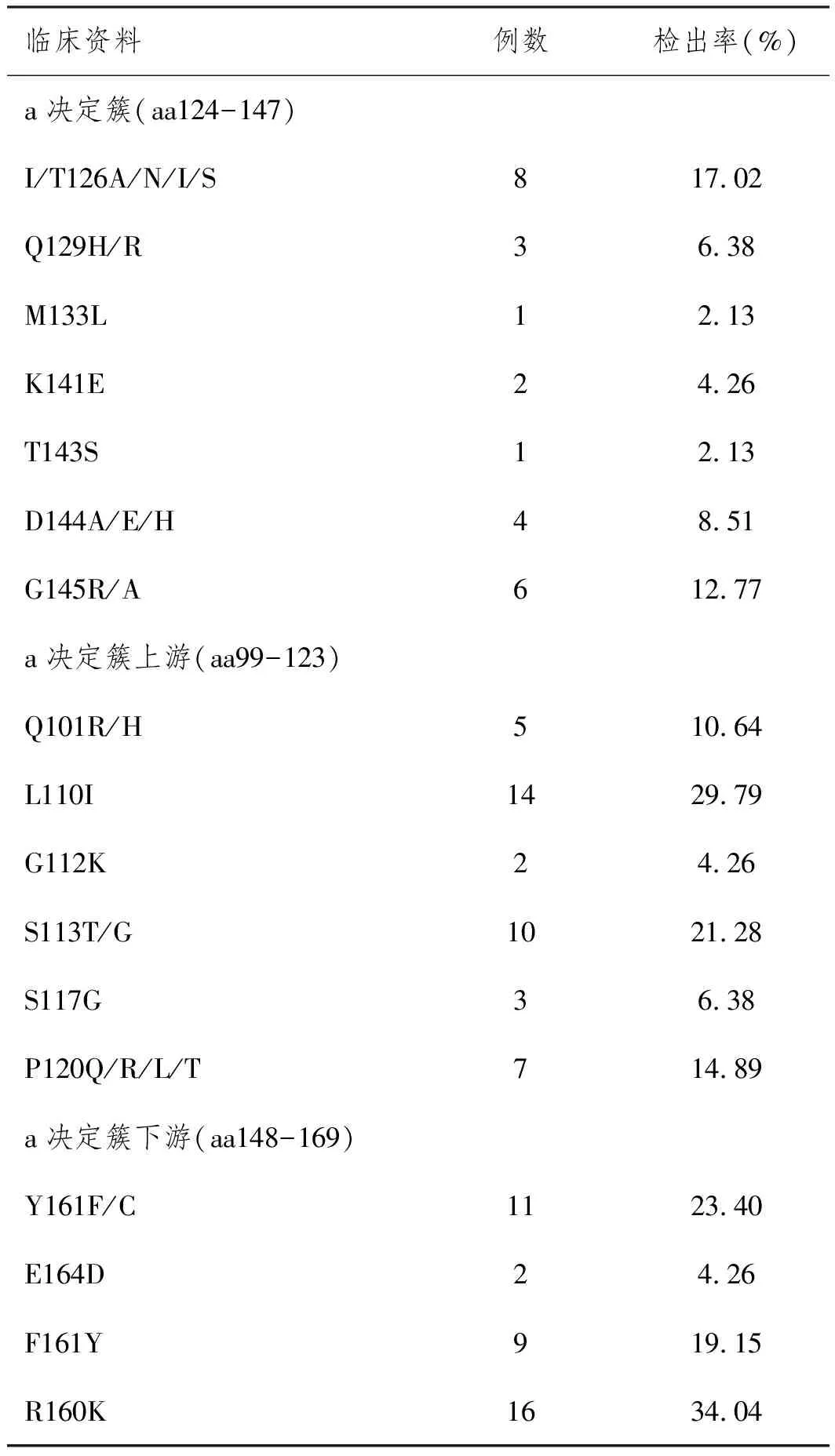
表4 HBV S區域核苷酸變異情況分析
2.5HBV S基因a決定簇氨基酸變異情況分析:30份發生氨基酸變異的HBsAg標本中,19份存在氨基酸極性改變,占63.33%,其中14份為B亞型毒株,且有11份發生于a決定簇(aa124-147)內,包括T126I、Q129R、T143S、D144A和G145A等5個變異位點,分別導致第126位氨基酸由蘇氨酸(Thr)突變為丙氨酸(Ala),第129位氨基酸由谷氨酰胺(Gln)突變為精氨酸(Arg),第143位氨基酸由賴氨酸(Lys)突變為蛋氨酸(Met),第145位氨基酸由甘氨酸(Gly)突變為Ala。
3 討 論
HBV感染是全球重要公共衛生問題,文獻報道約有20億人為HBV感染者或伴感染病史,其中持續感染者數量約2.92億,可引起肝炎、肝硬化或肝癌等嚴重病變,且每年致約60萬患者死亡[6]。我國為HBV主要流行地區之一,現階段感染者數量約8600萬,自1992年國內開始實施新生兒乙肝計劃免疫項目以來,HBV感染率呈現持續下降趨勢,但免疫失敗率約5%~10%,因此探討免疫失敗發生機制對HBV感染預防和治療具有重要意義[7]。
本研究以1973例按照0、1、6月標準程序接種HepB疫苗的患者為樣本進行橫斷面分析,結果顯示其中HBsAg陽性者47例,陽性率為2.38%,較我國60歲以下居民HBV攜帶率7.18%明顯降低[8],表明接種疫苗對預防HBV感染具有積極作用,同時也提示我國居民接種HepB疫苗后仍然存在一定免疫逃逸風險,因此HBV感染風險較高的人群需加強體檢和監測,以便及時采取預防和治療措施。同時本研究分析HBsAg陽性患者流行病學特征顯示,年齡>18歲者HBsAg陽性率明顯高于1-18歲人群,其原因可能與近年來疫苗接種技術快速進步密切相關,且1-6歲兒童HBsAg陽性率僅0.97%,達到衛生部提出的1%以下的要求。關于家族史的分析結果顯示父母HBV陰性者HBsAg陽性率明顯低于母親HBV陽性人群,可見HepB疫苗免疫逃逸可能與胎兒在功能已獲得感染存在密切聯系,從而導致母嬰傳播阻斷失敗。
基因型和血清型對判斷HBV傳播路徑具有重要參考意義,文獻報道HBV共包括A-J等10種基因型,其中分布于我國的主要為B、C和D三個亞型,優勢血清型則為adr型,其次為adw型,因不同血清對HepB疫苗反應效果存在差異,因此免疫失敗患者HBV血清型分布也可能發生變化[9]。本研究47份HBsAg陽性標本HBV基因型主要為B型(29.79%),其次則為C型(65.96%),另外發現D型2例(4.26%)。HBV血清型有S基因a決定簇產生,根據第122位和第160位氨基酸位點突變情況分為adw、adr、ayw和ayr等4種亞型,本研究47份HBsAg陽性標本血清型包括adr型23例(48.94%),adw型14例(29.79%)、ayr型8例(17.02%)和ayw型2例(4.26%),與全國流行規律相比無明顯差異[10]。由于樣本容量偏小,本研究未根據性別、年齡和家族史等口學特征進一步分析,因此HBV基因型和血清型與HepB疫苗免疫失敗是否存在聯系,以及不同亞型免疫逃逸風險是否具有差異均還有待深入探討。
近年來研究表明位于MHR(aa99-169)尤其a抗原決定簇(aa124-147)突變是影響HBsAg抗原性的重要因素,在自然條件、HepB疫苗或抗病毒藥物等作用下,a抗原決定簇突變導致HBsAg構象改變和抗原性變化,造成免疫逃逸[11]。Colagrossi等[12]對828位慢性乙型病毒性肝炎(CHB)患者進行分析,結果顯示耐藥誘導的免疫逃逸檢出率為28.6%,主要包括rtM204IsI196S、rtM204V-sI195M和rtV173L-sE164D等。本研究采用PCR技術對HBV S基因進行擴增并測序,結果顯示47份HBsAg陽性標本共存在核苷酸突變位點162個和氨基酸突變位點98個,明確存在氨基酸變異的標本為30份,檢出率63.83%,表明HepB疫苗免疫逃逸主要與HBV S基因有關。同時本研究基因測序顯示HBV S基因突變主要發生于MHR,其中a決定簇(aa124-147)內部突變發生率為53.20%,主要位點包括I/T126A/N/I/S、Q129H/R、M133L、K141E、T143S、D144A/EH和G145R/A等;a決定簇上游(aa99-123)突變發生率為87.24%,主要位點包括Q101R/H、L110I、G112K、S113T/G、S117G和P120Q/R/L等;a決定簇下游(aa148-169)突變發生率為80.85%,主要位點為Y161F/C、E164D、F161Y和R160K;可見發生HepB疫苗免疫逃逸的HBV毒株S基因突變情況較為復雜,可能同時存在a決定簇內外多個位點突變。為進一步明確HBV S基因突變與HepB疫苗免疫逃逸的關系,本研究對氨基酸突變類型進行分析,發現30份發生氨基酸變異的HBsAg陽性標本中,有19份存在氨基酸極性改變,占63.33%,且11份位于a決定簇內,是引起HepB疫苗免疫逃逸的主要原因。本研究檢測出的氨基酸突變包括T126I、Q129R、T143S、D144A和G145A等5個位點,其中T126I導致Thr突變為Ala,T143S導致LysMet,G145A導致Gly突變為Ala,均使氨基酸由極性和親水性和向非極性和疏水性轉變,導致抗原與抗體反應性下降,形成免疫逃逸。此外本研究顯示Q129R突變可將第129位氨基酸由Gln突變為Arg,親水性和抗原性均一定程度升高,但可能是由于多點位突變的關系,仍然導致免疫逃逸的結果發生。
綜上所述,HBV S基因突變與HepB疫苗免疫逃逸存在密切聯系,其中a決定簇(aa124-147)內部T126I、Q129R、T143S、D144A和G145A等位點突變可引起HBsAg極性和親水性變化,從而導致抗原性下降,造成免疫逃逸發生。本研究主要不足之處為未對不同突變情況下HBsAg結構進行微觀分析,尤其當多位點突變同時存在時,HBsAg抗原性變化情況還有待后續開展更多實驗進行深入研究,另外還有一部分無義突變和沉默突變與HepB疫苗免疫逃逸的關系也還有待探討和證實。
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中國生殖健康(2020年2期)2021-01-18 02:51:26
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
小學生導刊(2018年13期)2018-06-29 03:49:00
山東工業技術(2016年15期)2016-12-01 05:31:22
