石榴低溫響應因子CBF基因家族鑒定及其表達分析
2022-10-08 11:07:30劉龍博鄭樹軒
西北農業學報 2022年9期
關鍵詞:分析
劉龍博,鄭樹軒,鄭 潔
(1.淮北師范大學 生命科學學院,安徽淮北 253001;2.河南省農業廣播電視學校 夏邑分校,河南商丘 476400)
植物生長發育過程中不可避免地遭遇多種多樣的不良環境脅迫,其中低溫被認為是限制植物生長和地理分布的重要生態因素,也是影響農作物與園藝植物生產的主要自然災害。低溫脅迫可引起植物形態變化,并導致代謝紊亂,作物減產,甚至死亡[1]。研究指出植物可通過調控一系列生理生化過程以適應不利生長環境,如冷馴化(Cold acclimation,CA)被認為是溫帶植物提高耐寒性和安全越冬的重要策略[2]。CBF(C-repeat binding factors)又稱DREB(dehydration-responsive element binding),是參與冷馴化的重要轉錄因子[3]。它能夠識別并結合至低溫應答基因COR(cold-regulated)啟動子區的CRT/DRE(G/ACCGAC)元件參與冷馴化過程[4]。模式植物擬南芥CBF1/DREB1C、CBF2/DREB1B、CBF3/DREB1A能夠迅速響應低溫誘導,進而調控173個COR 參與低溫脅迫應答[5]。過表達AtCBF1能夠顯著提高水稻、煙草、草莓、番茄和黃瓜低溫脅迫抗性[6],而CBF1和CBF3敲除后擬南芥低溫脅迫抗性降低約60%[7]。以上研究表明CBF 在植物低溫脅迫抗性調控過程中發揮重要功能。
CBF屬于AP2轉錄因子家族的一個保守分支,除含有AP2結構域外,在其上下游還分布有PKKPAGRx KFx ETRHP 和DSAWR 兩 個 特 征氨基酸序列。擬南芥CBF基因家族共6個成員,除CBF1~CBF3外,還 包 含 有CBF4/DREB1D、DREB1E/DDF2和DREB1F/DDF1[8]。 但AtCBF4、At DREB1E和At DREB1F主要參與ABA、干旱以及鹽漬脅迫響應,而并非低溫。基于全基因組數據,大量CBF基因家族成員已在多個植物物種中被廣泛研究并證實參與植物低溫脅迫響應,如:小麥(Triticum aestivum)[9]、油菜(Brassica rapa)[10-11]、蘋 果(Malus domestica)[12]、茶樹(Camellia sinensis)[13-14]、月季(Rosa chinensis)[15]等。然而有關石榴CBF基因家族的研究鮮有報道。
石榴(Punica granatumL.)起源于中亞地區,經由絲綢之路引入中國,距今已有2 000多年栽培歷史。其果實外型美觀、口感鮮甜,含大量安石榴苷等鞣花單寧類抗氧化活性物質,具備潛在保健和醫用價值[16]。石榴花期長、樹姿美觀,兼具有觀賞價值,逐步被應用在園林綠化。石榴業已成為一種價值較高的經濟樹種。但石榴喜暖畏寒,在中國北方及黃淮石榴栽植片區冬春季氣溫較低,常有降溫霜凍甚至倒春寒等災害性天氣,石榴易遭遇周期性凍害問題,特別是以‘突尼斯’軟籽石榴為代表的軟籽系列品種耐寒性更差,不利于優良品種推廣[17]。但目前石榴抗寒性研究主要集中在通過測定生理生化指標比較不同石榴品種低溫脅迫抗性方面[17-18]。‘大笨籽’‘泰山紅’‘突尼斯’基因組密碼的成功破譯,高質量基因組圖譜的成功構建,為進一步系統開展石榴分子生物學和遺傳改良奠定了研究基礎[16,19-20]。本研究從石榴全基因組中共鑒定到7個CBF 基因家族成員,基于生物信息學方法對PgCBFs基因家族成員氨基酸保守序列、染色體定位、理化性質、進化關系、外顯子/內含子排列、蛋白質二級與三級結構、共線性關系、GO 注釋、啟動子順勢作用元件和表達模式等方面進行綜合分析,為進一步探索石榴CBF家族基因功能提供參考和理論依據。
1 材料與方法
1.1 試驗材料與處理
‘突尼斯’軟籽石榴(Punica granatumL.‘Tunisia’)扦插苗種植于淮北師范大學園藝實驗基地溫室內,溫度控制在26°C 左右,濕度為60%~70%,光照時間為白天14 h,夜間10 h,栽培基質為泥炭土、珍珠巖和蛭石(體積比為3∶1∶2),低溫處理前每5 d澆水1次;采用上口徑和高分別為28 cm 與30 cm 的種植盆進行栽種,每盆栽3株,每5盆為1次重復。試驗所采用的石榴幼苗均生長穩定且長勢一致,無病蟲害。低溫處理時將石榴幼苗與種植盆共同放置于低溫冷藏柜中,處理溫度為-4℃,分別在處理后0、1、3、6、12、24 h收集幼嫩不定根和1 a生枝條韌皮部。每個樣品收集時間點均設置3次重復,樣品收集后置于液氮處理并儲存于-80 ℃低溫冰箱,待用。
1.2 石榴CBF基因家族成員鑒定
石榴基因組數據下載自NCBI(https://www.ncbi.nlm.nih.gov/genome/? term=Punica+granatum)[20],擬南芥和水稻基因組數據分別來源于TAIR(http://www.arabidopsis.org)和RGAP(http://rice.uga.edu/)。使用7個擬南芥(Arabidopsis thaliana)和10 個水稻(Oryza sativa)CBF 蛋白序列通過本地Blast初步鑒定石榴CBF 候選基因(E<1.00E-10)。在Pfam 數 據 庫(http://pfam.xfam.org/)中 下 載AP2結構域(PF00847)種子文件,通過HMMER 3.1軟件鑒定含有AP2結構域的候選基因家族成員(E<1.00E-10)。提取Blast與HMMER 相同候選基因蛋白序列,利用SMART(http://smart.emblheidelberg.de)數據庫和Clustal W2(https://www.ebi.ac.uk/Tools/msa/clustalo/)分析候選PgCBFs蛋白質保守結構域與CBF基因家族特征序列,確定石榴CBF 基因家族成員。
根據基因組注釋文件,通過TBtools軟件對PgCBF基因家族成員進行染色體定位繪圖[21],并根據染色體定位信息對PgCBF 成員命名。PgCBF基因家族成員m RNA 和CDS 數據均來自于基因組注釋信息。通過Ex Pasy 在線網站(http://www.expasy.org/tools/)分析各成員的蛋白質長度、分子量和理論等電點。利用在線工具BUSCA(http://busca.biocomp.unibo.it/)對7個PgCBF成員亞細胞定位進行預測。
1.3 系統進化樹構建
除石榴7個PgCBF基因家族成員外,收集擬南芥(A.thaliana)、水稻(O.sativa)、大豆(Glycine max)、番 茄(Solanum lycopersicum)、桃(Prunus persica)、蘋果(M.domestica)以及茶樹(C.sinensis)等7個物種的49個CBF 基因家族氨基酸序列,通過MEGA X 軟件構建進化樹。設定條件為:最優替代模型使用JTT+ G,構建方法使用最大似然法(Maximum Likelihood method,ML),Bootstrap設置為1 000重復。
1.4 石榴PgCBF基因結構、保守基序和蛋白質二級及三級結構分析
根據7個物種的GFF3文件,提取50個CBF基因家族成員的基因序列、外顯子、內含子以及上下游非翻譯區長度與排布信息。通過Motif Elicitation(http://meme-suite.org/tools/meme)分析CBF基因家族成員的保守基序類型,設置鑒定到MEME 數量為10 個。利用TBtools工具對CBF基因家族系統進化樹、基因結構和保守基序進行可視化[21]。分別利用NPS@:SOPMA(https://www.predictprotein.org/signin)和Ex-PaSy Swiss-Model(http://swissmodel.expasy.org)預測PgCBF基因家族成員蛋白質二級和三級結構。
1.5 CBF基因家族成員共線性分析
蘋果(M.domestica)和桃(P.persica)基因組數據下載自GDR(https://www.rosaceae.org/)。使用MCScan X 軟件對4 個物種的基因組序列進行比對,獲得它們的CBF基因家族共線性關系,使用TBtools繪制基因共線性結果[21]。
1.6 PgCBF基因家族成員GO 注釋分析
將石榴蛋白序列數據比對到Swiss-Prot數據庫,利用TBtools 進行蛋白序列GO 注釋分析[21]。
1.7 石榴PgCBFs基因家族啟動子區順式作用元件分析
使用TBtools提取PgCBF 基因家族各成員起始密碼子(ATP)上游2 000 bp的堿基序列作為啟動子區域,利用plantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)進行順式作用元件分析。通過TBtools構建熱圖呈現順勢作用元件分析結果[21]。
1.8 石榴PgCBFs基因家族轉錄表達分析
分別選取石榴根、葉片、花、內種皮、外種皮和果皮轉錄組數據,分析石榴PgCBF基因家族在不同組織中的表達模式。石榴轉錄組數據下載自NCBI SRA 數據庫(No.SRP100581)[19]。下載獲取轉錄組后,使用TBtools轉錄組數據分析系列功能,通過FastQC進行質控,利用Trimmomatic去除接頭,使用Kallisto對基因表達分析,通過TBtools繪制基因表達熱圖[21]。
1.9 RNA提取與石榴PgCBFs表達分析
使用RNA 提取試劑盒(TIANGEN,China)提取石榴根和枝條韌皮部總RNA,去除基因組DNA 后,合成cDNA,并使用DEPC水稀釋10倍用做qRT-PCR 模 板。QRT-PCR 使 用20μL 反應體系,包含1μL cDNA 模板、0.5μL上和下游引物、10μL SYBR 預混液(cham Q SYBR qPCR Master Mix,Vazyme,China)以及8μL DEPC水。使用ABI 7300進行qRT-PCR 反應,設定程序為:95 ℃5 min預變性;95 ℃5 s,60 ℃35 s,40個循環。基因表達量使用Pg Actin作為內參基因,通過2-ΔΔCT方法計算相對表達量[22]。本研究使用的引物信息見表1。
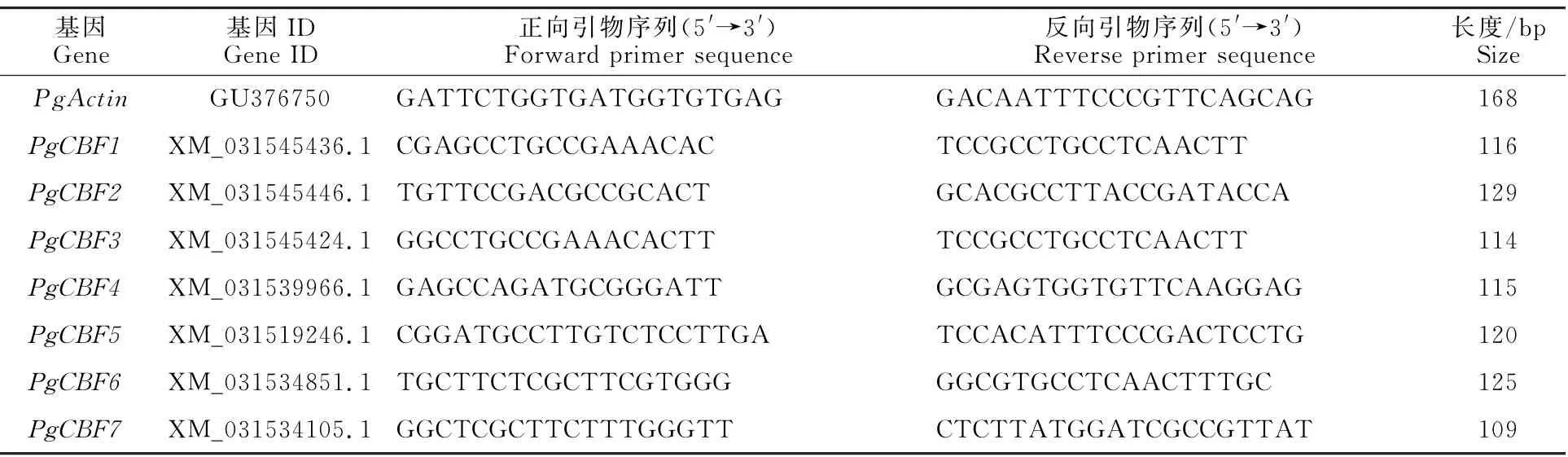
表1 石榴PgCBF基因qRT-PCR 引物Table 1 Primers used for qPCR assay of PgCBF genes in pomegranate
2 結果與分析
2.1 石榴CBF基因家族成員鑒定與染色體定位分析
利用17個擬南芥和水稻CBF蛋白序列通過Blast以及HMM 方法,去除冗余序列后,在石榴基因組中鑒定到7 個CBF 候選基因家族成員。通過多序列比對(圖1),7個PgCBF 成員與擬南芥和水稻CBF 基因家族成員類似,除AP2結構域外,也包含有2個保守氨基酸序列(PKK/RPAGRx KFxETRHP和DSAWR),分別位于AP2結構域上下游,表明鑒定到7個PgCBFs為石榴CBF基因家族成員。

圖1 擬南芥、水稻、石榴CBF家族基因蛋白質多序列比對Fig.1 Multiple alignment of CBF protein from A.thaliana,O.sativa and P.granatum
根據PgCBF基因家族成員的染色體位置信息將它們命名為PgCBF1~PgCBF7(圖2-A)。7個成員分布在2 條染色體上,PgCBF1~PgCBF5集中分布在1號染色體下部,聚集在一起形成基因簇(圖2-B),PgCBF6和PgCBF7分別位于4號染色體的上部和下部。

圖2 石榴7個CBF基因染色體分布Fig.2 Distribution of seven CBF genes in pomegranate chromosome
2.2 石榴CBF基因家族成員序列理化性質分析
如表2 所示,除PgCBF1和PgCBF3外,其余5個PgCBF m RNA 片段大小各不相同,長度約為1 119~2 763 bp。PgCBF1、PgCBF3和PgCBF5編碼區(714 bp)和蛋白質(238 aa)長度一致,蛋白質分子質量(MW)大小相近,PgCBF7與它們相似。7個成員理論等電點(pI)為4.89~5.60;亞細胞定位預測與轉錄因子蛋白功能定位一致,均處于細胞核內。

表2 PgCBF基因家族成員理化性質分析Table 2 Analysis of physical and chemical properties of family members of PgCBF gene family
2.3 CBF基因家族進化樹構建
使用石榴等8個物種的56個CBF基因家族氨基酸序列構建進化樹。將CBF 基因家族分為4個亞組,分別包含9、8、19和20個成員(圖3)。水稻除OsCBF2 外,其他9 個成員全部位于GroupⅠ,表明GroupⅠ中的CBF成員在單子葉植物中可能有獨特的作用[23]。與其他研究結果一致,擬南芥6 個成員分布于Group Ⅲ和Ⅳ中[23],PgCBF2、PgCBF4~PgCBF6與At-DREB1F和AtDREB1E具有較近的親緣關系,PgCBF7與AtCBF1~AtCBF4、MdCBF1、PpCBF6、SlCBF1、CsCBF3等 均 歸 屬 于GroupⅢ。PgCBF1和PgCBF3具有較近親緣關系,均歸屬于Group Ⅱ。

圖3 石榴與擬南芥、水稻、大豆、番茄、桃、蘋果及茶樹CBF基因家族進化樹分析Fig.3 Phylogenetic tree analysis of CBF gene family from pomegranate(Pg),Arabidopsis(At),rice(Os),soybean(Gm),tomato(Sl),peach(Pp),apple(Md)and tea(Cs)
2.4 CBF家族基因結構、保守基序及結構域分析
為進一步分析CBF基因家族進化關系,綜合分析包括PgCBF 在內的50個CBF 基因結構和保守基序(圖4)。通過基因組GFF 注釋文件對CBF基因家族CDS分布以及外顯子和內含子統計分析,結果顯示大部分CBF成員基因結構相似僅有1個外顯子,不包含內含子,為內含子缺失型(圖4-A)。僅有少部分成員含有1~2個內含子,如:At DERB1E、PgCBF2、PgCBF4和PgCBF5。
通過MEME在線工具預測CBF基因家族成員的10個保守基序(圖4-B),各保守基序的氨基酸長度為11~25 aa(表3)。除了少部分成員外,大多數CBF蛋白含有motif 1~7,且在不同CBF亞組中排列較為一致(圖4-B),表明這些motif在CBF家族成員高度保守。除與其他物種CBF成員基本一致的motif組合與排列順序外,7個PgCBF成員中有6個成員含有motif 9(圖4-B)。

表3 石榴和其他6個物種CBF蛋白保守基序分析Table 3 Analysis of conserved motif of CBF protein in pomegranate with other six species

圖4 CBF基因家族基因結構和保守基序分析Fig.4 Analysis of gene structure and conserved motif elements of CBF gene family
2.5 石榴CBF家族基因蛋白質二級和三級結構預測
對7個石榴CBF 基因家族成員蛋白二級結構預測發現該家族包含α-螺旋、延伸鏈、β-轉角和不規則卷曲(表4)。在7個編碼蛋白中主要是α-螺旋(22.56% ~34.50%)和不規則卷曲(44.50%~59.02%),延伸鏈(9.05%~15.79%)和β-轉角(0.42%~5.50%)較少。
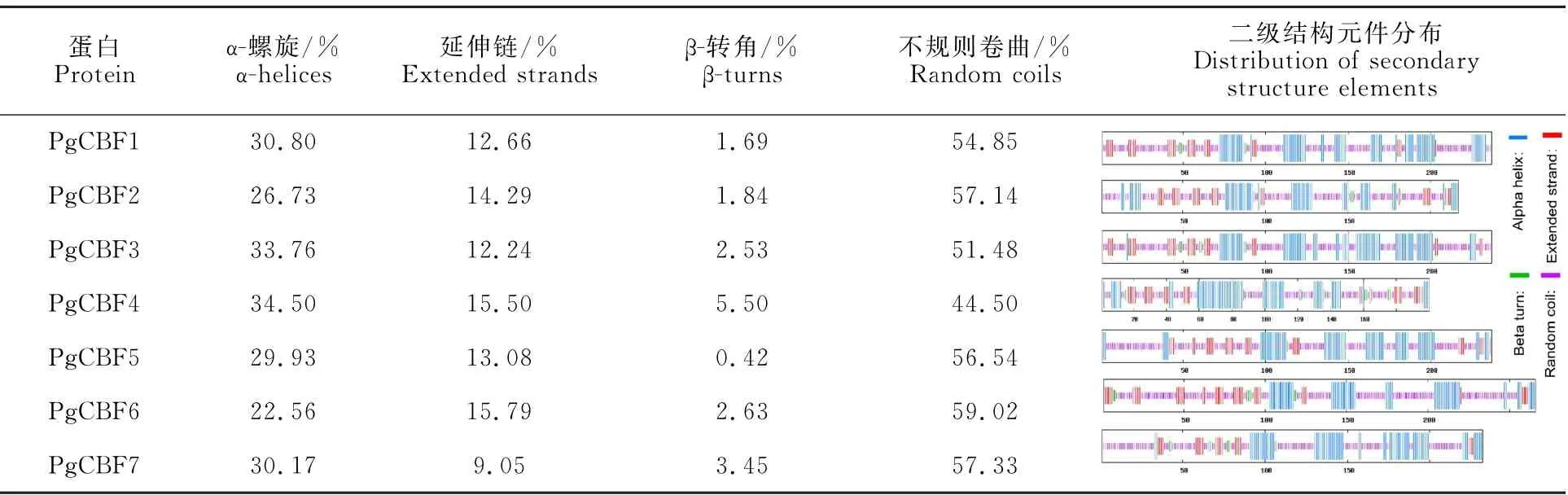
表4 石榴CBF基因家族蛋白二級結構分析Table 4 Secondary structure of protein sequence of CBF gene family in pomegranate
7個石榴CBF 基因家族成員蛋白三級結構與模型5wx9.1.A 較為相似(42.65%~46.58%)(圖5),且7個PgCBFs蛋白的三級結構較為相似,均由1個α-螺旋和2個β-折疊構成。

圖5 7個石榴CBF基因家族蛋白三級結構預測Fig.5 Tertiary structure of seven CBF proteins in pomegranate
2.6 石榴PgCBF基因共線性分析
如圖6所示,在石榴PgCBF家族間存在1個串聯復制基因對(PgCBF4/PgCBF5)。并鑒定到石榴共與擬南芥、蘋果和桃的10個CBF 基因家族成員存在共線性關系。其中,PgCBF1分別與AtDREB1F、MdCBF4、MdCBF3和PpCBF1具有共線性關系;PgCBF2與At DREB1E具有共線性關系;PgCBF4與PpCBF3具有共線性關系;PgCBF6與MdCBF3以及PpCBF1具有共線性 關 系;PgCBF 7與MdCBF 1、MdCBF 2和PpCBF6具有共線性關系。
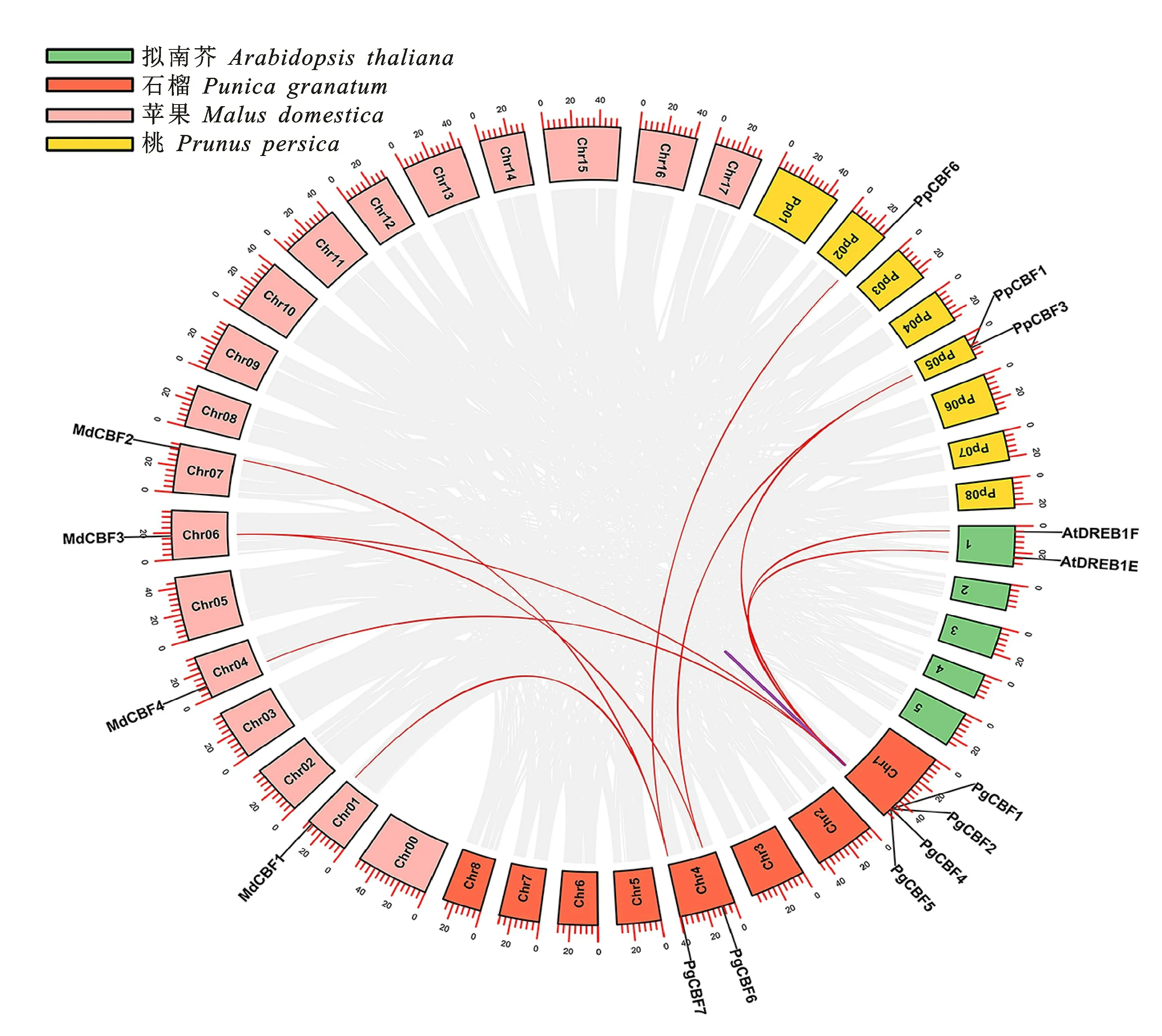
圖6 石榴、擬南芥、蘋果和桃的CBF基因家族共線性分析Fig.6 Synteny analysis of CBF genes family of P.granatum,A.thaliana,M.domestica and P.persica
2.7 石榴PgCBF基因GO 注釋
利用GO 注釋分析發現7個PgCBF 可被劃分在分子功能、細胞組分、生物過程3 個大類的26個亞類中(圖7)。分子功能主要涉及順勢作用元件結合與轉錄調控(10個亞類),細胞組分主要參與細胞核內組分形成(1個亞類),生物過程中涉及冷馴化(GO:0009631)、響應低溫、高溫與環境脅迫刺激(GO:0050826、GO:0009408、GO:0006950、GO:0050896)、參與赤霉素、ABA 等激素合成與信號轉導過程(GO:0009686、GO:0046885、GO:0009738、GO:0071215、GO:0010817、GO:0009737、GO:0009737、GO:0032870)。GO注釋表明7個PgCBF參與轉錄調控與脅迫及植物激素刺激應答,尤其是溫度脅迫。

圖7 石榴PgCBF基因家族成員GO 注釋分析Fig.7 Information from gene ontology(GO)annotation of PgCBFs
2.8 石榴PgCBF家族基因順式作用元件分析
位于上游啟動子區的順勢作用元件是植物內部信號和脅迫刺激響應的重要調控區域。對7個石榴CBF基因家族起始密碼子上游2 kb堿基序列進行分析。鑒定到一系列與脅迫相關(低溫、干旱、厭氧)、激素相關(ABA、MeJA、GA、SA、生長素)以及光誘導相關順勢作用元件(圖8)。在脅迫誘導的啟動子元件中,除PgCBF6外,6個成員含有16個厭氧誘導元件ARE;有5 成員含有6個低溫應答元件LTR;4個成員含有7個干旱誘導元件MBS。在激素響應元件中,在7個成員中鑒定到36個ABA 響應元件ABRE,40個MeJA應答元件CGTCA-motif和TGACG-motif,2 個GA 應答元件P-box,7 個生長素應答元件Aux RR-core、TGA-element、TGA-box,5 個SA應答元件TCA-element。在PgCBF 啟動子區還存在大量光誘導元件,其中G-box 元件最多,共在7個成員共鑒定到40個。結果表明PgCBF參與響應石榴脅迫應答、激素和光誘導。
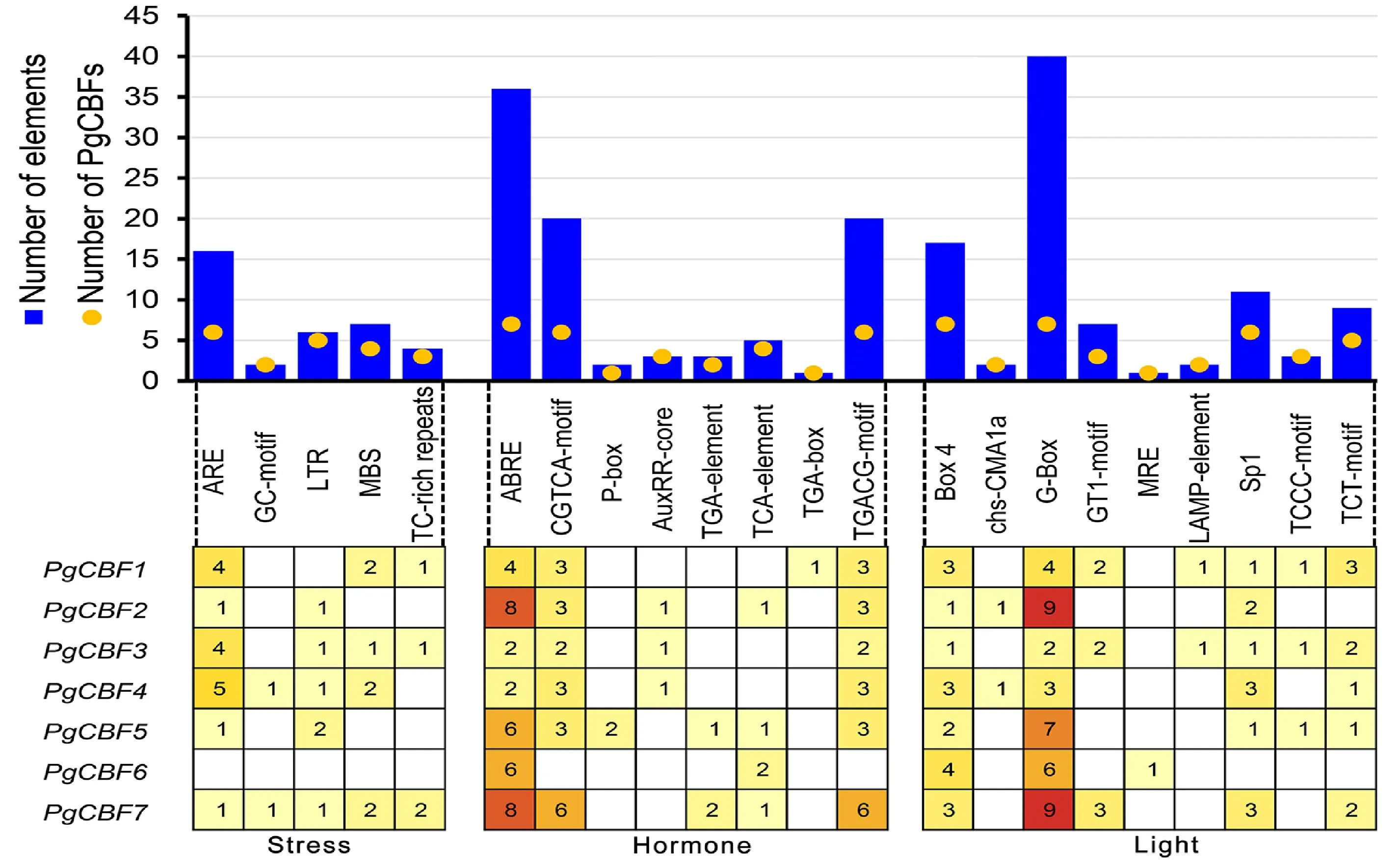
圖8 石榴CBF基因家族啟動子順式作用元件分析Fig.8 Analysis of putative cis-regulatory elements in promoter regions of CBF genes family in pomegranate
2.9 石榴PgCBF家族基因表達分析
通過石榴轉錄組數據[19],在石榴根、葉片、花和果實不同組織中對7個PgCBF 成員的表達模式進行分析(圖9)。結果顯示,除PgCBF6外,其余6個基因在根中顯著表達,其中PgCBF5的表達量最高。PgCBF2和PgCBF5在花和果皮中表達顯著高于其余成員。PgCBF6僅在果皮中有顯著表達。與其他成員相比,PgCBF7在果實內種皮和外種皮中表達上調。但所有PgCBFs在葉片中只有痕量表達或不表達。結果表明PgCBF 成員在不同組織中發揮功能存在差異。
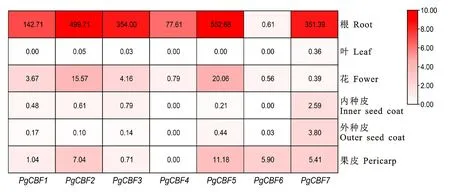
圖9 PgCBFs在石榴不同組織中的表達熱圖Fig.9 Heatmap of expression level of PgCBFs in different tissues of pomegranate
在低溫處理后的石榴根和枝條韌皮部中PgCBF基因家族成員表達量均顯著上調,但存在差異(圖10)。在根中,PgCBF1、PgCBF2、PgCBF4和PgCBF5在低溫處理后0~6 h表達量逐步升高并達到最大值,其中PgCBF2、PgCBF4和PgCBF5在6 h表達量均為0 h的10倍以上,而PgCBF1僅為0 h的2.2倍。在枝條韌 皮 部,PgCBF2、PgCBF3、PgCBF4和PgCBF5表達量均在處理后6 h 達到最高峰。PgCBF6在根和韌皮部表達趨勢一致,在處理24 h后達到最大值。PgCBF7在低溫處理1 h后表達量顯著升高并達到最大值,分別較0 h根和韌皮部升高35.3和23.3倍。在低溫處理12至24 h后,PgCBF7在根和韌皮部中仍顯著上調,上調8.3~13.0倍,表明PgCBF7能夠快速響應低溫誘導并持續應答低溫脅迫。
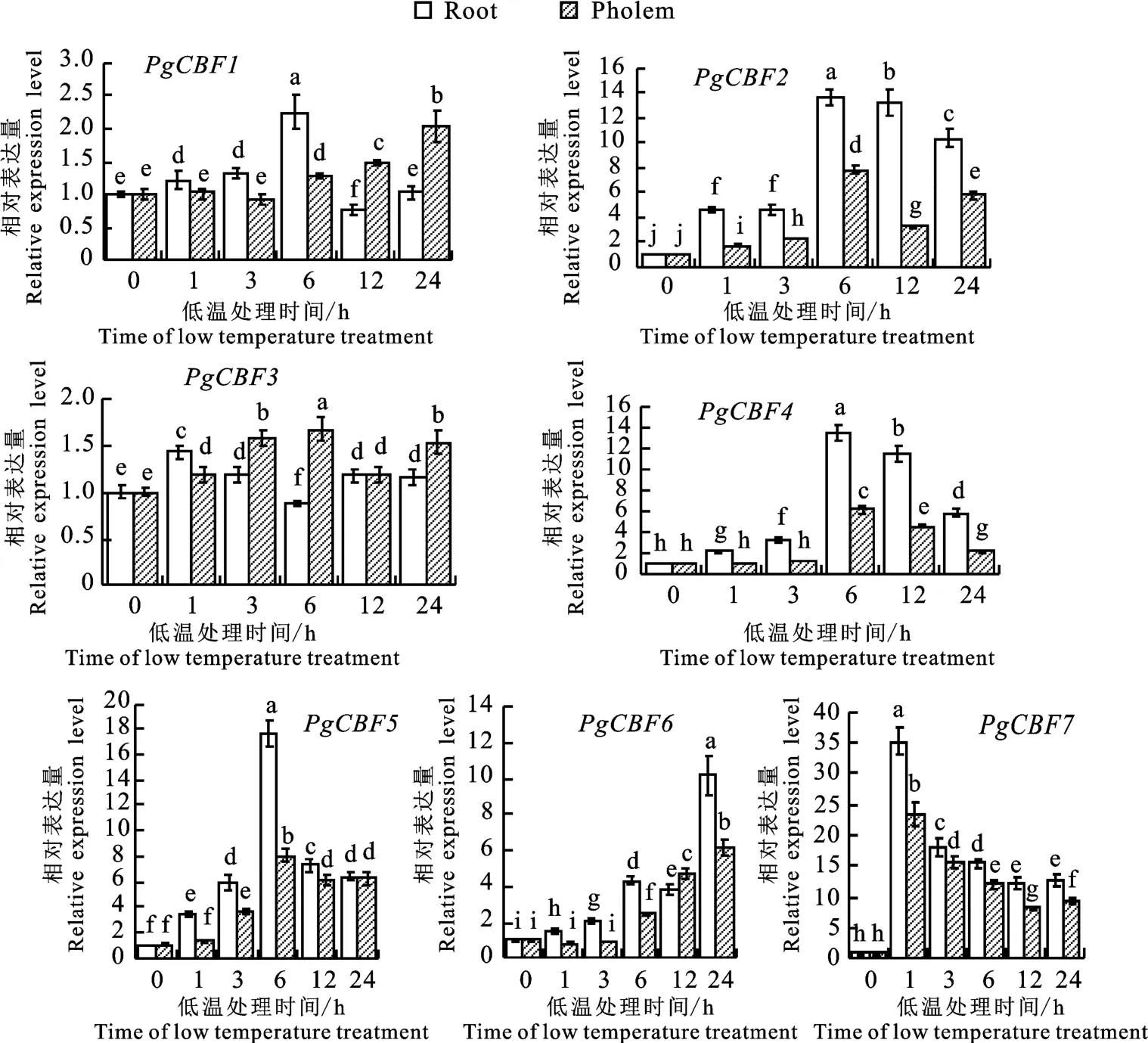
圖10 低溫處理后PgCBF在石榴根和枝條韌皮部中的相對表達量Fig.10 Relative expression of PgCBF in pomegranate roots and phloem under low temperature treatment
3 討論
低溫是制約植物生長發育的一個主要非生物脅迫因子。植物遭受冷脅迫時,約有12%的轉錄組基因參與CBF轉錄因子轉錄表達調控過程,以增強植物耐寒能力[24]。隨著植物全基因組測序、組裝和注釋研究的進一步發展,通過比較基因組方法在不同的植物物種中識別和鑒定到越來越多的CBF基因家族。通過分析發現CBF是一個小的基因家族,如在擬南芥(A.thaliana)、水稻(O.sativa)、油 菜(B.rapa)、大 豆(G.max)、蘋 果(M.domestica)、桃(P.persica)、梨(Pyrus bretschneideri)、葡萄(Vitis vinifera)[23]、鵝掌楸(Liriodendron chinense)[25]和茶樹(C.sinensis)[14]中分別鑒定到6、10、9、11、4、6、11、4、14和6個CBF成員。同樣的,本研究共在石榴全基因組中僅鑒定到7個CBF基因家族成員(表2),包含1個串聯復制基因對(PgCBF4/PgCBF5)(圖6),但在染色體間未鑒定到片段重復(圖6),這可能是CBF家族成員較少的一個原因。鑒定到的7個PgCBF蛋白序列中均含有由59個氨基酸殘基組成的高度保守AP2結構域和2個特征序列(圖1),其中位于N 端的特征序列內的PKK/RPAGRx KF屬于細胞核定位信號(nuclear localization signal,NLS)序列,表明PgCBF 定位在細胞核 內(表2),與 棉 花 GhDREB1[26]、茄 子Sm CBF[27]、茶樹CsCBF[14]亞細胞定位分析結果一致。
構建系統進化樹有利于分析基因家族成員蛋白質結構與功能。與其他雙子葉植物CBF 成員分布一致(圖3)[23],PgCBF成員分布在GroupⅡ~Ⅳ。在石榴中,PgCBF2、PgCBF4、PgCBF5、PgCBF6與At DREB1F、At DREB1E共 同 歸 屬于Group Ⅳ,且均含有ABRE 順勢作用元件(圖8),推測它們可能參與ABA 信號轉導或脅迫應答過程(圖7)。在Group Ⅲ中多數成員能夠顯著響應低溫脅迫,如:蘋果MdCBF1轉基因至擬南芥能夠顯著誘導AtCOR15a、AtRD29A和At RD29B表達,低溫脅迫抗性明顯增強[12];PpCBF6在桃遭遇低溫脅迫時表達量顯著升高,能夠結合至Pp VIN2啟動子區抑制其表達,降低液泡轉化酶(vacuolar invertase,VIN)活性,提高蔗糖含量,從而緩解低溫對果實的傷害[28];在低溫脅迫條件下,番茄slcbf1突變體更易遭受低溫脅迫傷害,電導率和丙二醛含量明顯升高[29];茶樹CsCBF3在擬南芥中過表達后,能夠顯著誘導AtCOR15a和At RD29A表達上調,明顯改善了轉基因植株在-4 ℃和-8 ℃的生長狀態[14]。在石 榴 中,PgCBF7與MdCBF1、MdCBF2、PpCBF6均歸屬Group Ⅲ,并與它們存在共線性關系(圖6),GO 注釋(圖7)、LTR 順序作用元件(圖8)以及基因表達分析(圖10)等結果表明PgCBF7能夠直接并快速響應低溫脅迫,其可能在低溫脅迫中發揮重要功能,但具體分子機制還需進一步研究說明。
PgCBF基因家族成員與已鑒定到的大部分CBF基因家族成員平均蛋白質長度、分子質量均值類似[14-15]。同時,不同基因亞組在基因結構和motif組成與排列上并沒有明顯的區分(圖4),PgCBF蛋白二級和三級結構類似(表4,圖5),表明CBF基因家族在進化過程中相對保守[13]。
一些轉錄因子已被證實能夠結合至CBF 啟動子區參與其表達調控,如ICE1/2、CAMTA1-3、PIF4/7、EIN3、MYB15 等[15]。在石榴PgCBF啟動子區鑒定到大量脅迫應答、激素響應與光誘導元件(圖8)。雖然PgCBF1和PgCBF6啟動子區并未鑒定到LTR 元件,但在低溫處理誘導后顯著誘導表達量上升(圖10),表明在石榴中可能存在不依賴LTR 元件調控CBF表達途徑。如研究指出,PIF4/7與CBF1和CBF2基因啟動子中的G-box結合后能夠在長日照條件下抑制CBF1和CBF2表達,而在短日照條件這種抑制可被解除,從而導致CBF表達上調,增加耐寒性[30]。此外,CBF還參與植物激素信號轉導。如外源Me-JA 處理能夠顯著誘導蘋果愈傷組織中Md-CBF1、MdCBF2、MdCBF3表達,增強愈傷組織低溫抗性[31]。在敲除番茄slcbf1后,吲哚乙酸的含量明顯增加,但MeJA、ABA 和玉米素核苷的含量減少[29]。需要注意的是,雖然在7個啟動子中共鑒定到36個ABRE元件(圖8),但研究發現ABA可直接結合至COR基因啟動子區的ABRE元件,而提高植物耐寒性并不依賴CBF 途徑[32];然而,目前仍然缺少ABA與CBF基因表達調控相關報道,但可能存在某些機制,它可以通過與其啟動子結合來調節CBF表達[33]。仍需要進一步研究以闡明ABA和CBF基因之間的復雜調節關系。
本研究共在石榴基因組中鑒定到7 個CBF基因家族成員,它們的蛋白質序列與結構相對保守;通過GO 注釋與順式作用元件分析,發現PgCBF基因家族與石榴激素和脅迫響應緊密相關,尤其在低溫條件下各基因表達均明顯上調,特別是PgCBF7能夠快速響應低溫誘導并持續高表達,表明PgCBF7可能在石榴幼苗低溫脅迫應答中發揮重要調控功能。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
