西藏飛蝗的研究現狀及展望
2022-09-28 12:19:38王向向顧亞欣任麗娜母澤祺唐曉琴
廣西農學報 2022年3期
王向向 顧亞欣 任麗娜 母澤祺 唐曉琴, 3*
(1. 西藏農牧學院植物科學學院, 西藏 林芝 860000;2. 西藏高原資源昆蟲與應用昆蟲實驗室, 西藏 林芝 860000; 3. 西藏高原森林生態教育部重點實驗室, 西藏 林芝 860000)
西藏飛蝗(Locusta migratoria tibetensis Chen)屬于直翅目(Orthoptera)蝗亞目(Locustodea)飛蝗科(Locusta migratoria)飛蝗屬(Locusta), 是1963年陳永林先生在西藏自治區發現并命名的飛蝗新亞種。西藏飛蝗活動范圍主要在我國的西藏自治區、青海省、四川省等高原地區, 取食廣泛且食量大, 主要取食禾本科與莎草科的作物和牧草, 是一種高原重要害蟲, 具有繁殖能力強、密度大、為害面積廣和為害程度嚴重的特點。根據李婧研究發現, 在我國歷代古籍中有許多處蝗災的記載, 嚴重時導致莊稼顆粒無收和草場的寸草不生, 造成饑荒和出現難民, 引起社會混亂。吳志剛和秦萌研究西藏飛蝗對青稞產業的經濟損失進行評估時, 結果顯示西藏飛蝗每年造成的潛在經濟損失達0.713億~4.542億元。隨著全球氣候變暖, 青藏高原的生態環境也受到嚴重影響, 導致西藏飛蝗的繁殖和發育能力增強并且增加了越冬存活率, 使西藏飛蝗暴發更加容易, 為害更加嚴重。本文主要對西藏飛蝗的部分特征以及綜合防治措施等研究現狀進行綜述。
1 西藏飛蝗的形態學特征
西藏飛蝗的生長發育類型屬于漸變態發育。與其他漸變態發育昆蟲相似, 一生需要經歷3個發育階段。其中蝗蝻需要經過5次蛻皮、羽化才能完全發育為成蟲, 即一只西藏飛蝗一生共經歷5個齡期。觀察發現西藏飛蝗的受精卵形狀類似長橢圓形且每一個卵囊中卵的顆粒數不相等, 各齡蟲態的前胸背板后緣的形狀、翅芽的長度、觸角的節數和體色等也各不相同, 西藏飛蝗成蟲的體長、體色也會因雌雄不同有所差別。
西藏飛蝗成蟲分為頭部、胸部和腹部三個部分, 其頭部的額唇基區有1對絲狀觸角, 顱側區有1對復眼和3個單眼, 頰下區有1個咀嚼式口器。胸部的三個節分別有1對足, 共有3對足, 其前足、中足為爬行足, 后足為跳躍足;胸部含有前后2對通明翅, 前翅狹長, 具有斑紋, 后翅平常時呈折起狀態, 覆蓋前翅下;遷徙時, 展開進行平衡身體。成蟲腹部第1節具有鼓膜器, 能發出聲音, 成蟲末端含有生殖器官, 因雌雄性別不同, 其生殖器的形態特征也不相同。
2 西藏飛蝗的生物學特性
2.1 西藏飛蝗的生活史
西藏飛蝗以蟲卵的形式在土壤的表皮中越冬。西藏飛蝗的年生活史以四川甘孜州鄉城縣調查為 例, 如表1所示。由表1可知, 西藏飛蝗在四川部分地區第一年發生1~2代, 卵在土壤中越冬并于第二年3月開始孵化, 到4月為卵的孵化盛期, 5月和6月基本是以蝗蝻為主, 7月的蝗蝻開始羽化, 7月至8月為4齡蝗蝻的羽化盛期, 8月至9月為成蟲的產卵盛期。西藏飛蝗的活動范圍受外界溫度和光照的影響比較大, 其全世代發育的有效積溫相對偏高。當溫度過高和過低時都會影響其取食、蛻皮、羽化等活動;成蟲在光照不足或者溫度較低的場所會使其產下的卵孵化率大幅度降低。
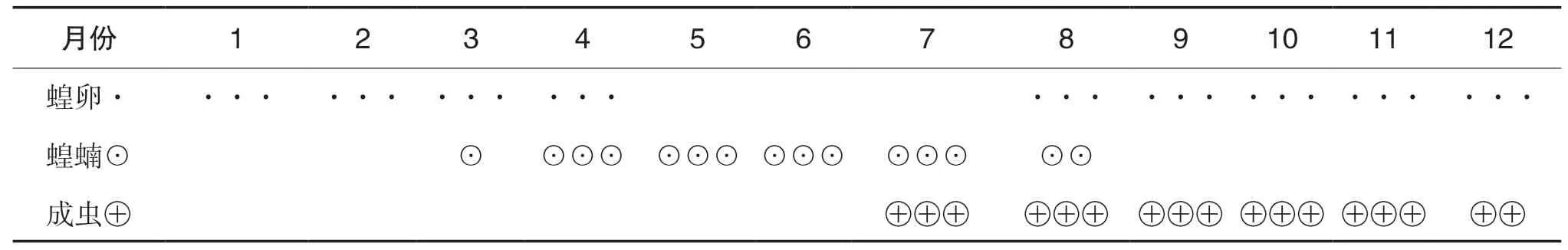
表1 西藏飛蝗的年生活史(四川甘孜州鄉城縣)
王思忠等利用培養箱在溫度不同, 其他條件相同的條件下飼養西藏飛蝗。測量蟲卵和各齡蟲態的發育起始溫度和有效積溫, 結果顯示:蝗卵、各階段的蝗蝻、成蟲的發育起點溫度和有效積溫各不相同。由此, 可以說明各齡蟲態發育歷經的時間也不相同。
2.2 西藏飛蝗的習性
西藏飛蝗成蟲產卵根據地形、土壤含水量和土壤理化性質等不同有著明顯的選擇性。李慶等研究表明西藏飛蝗會選擇堅實平坦、濕度合適的土壤產卵。
西藏飛蝗主要取食禾本科農作物和蒿草、披堿草等禾本科與莎草科雜草;在不同的寄主植物下, 其會優先取食含水量和營養價值高的植物。1~3齡蝗蝻喜歡取食幼嫩的禾本科雜草, 但是取食量相對較少;成蟲期會進入暴食期, 而因青稞和小麥的長勢和含營養含量此時比雜草要好, 成蟲將主要以青稞和小麥為食, 造成農作物的減產。有研究證明8月的成蟲密度大且是產卵的盛期, 能夠在禾本科植物上獲得生長發育所必需的營養物質, 但在沙棘和十字花科白菜上則不能完成整個生活史。
西藏飛蝗具有群居型和散居型兩種生活類型, 但兩種類型可以隨其生存環境與其種群密度的變化相互轉化;兩種生活方式類型下的蝗蟲體色、體型的大小、身體各個部分等形態學特征都有所不 同。封傳紅等研究證明西藏飛蝗具有群集遷徙的習性, 當西藏飛蝗蟲口密度達到一定數值時, 將會由分散型轉變為群居型進行遷徙。此外, 西藏飛蝗群集會受到取食的食物、蟲口密度和其他行為反應的影響。陳湖海等研究西藏飛蝗觸角的意義發現西藏飛蝗能夠通過群集行為增加遷徙距離、增加遠距離食物取食的概率。李彝利通過掃描電鏡對西藏飛蝗觸角外部形態結構特征觀察發現不同的西藏飛蝗具有不同的感受器, 共計有5種感受器類型。感受器因蝗蟲的雌雄、散居型、群居型的類型并無明顯差異, 僅在數量上不同。不同蟲態的西藏飛蝗對同一種人工合成化合物的觸角電位反應(EAG)總體呈現散居型強于群居型、成蟲強于蝗蝻、雄性強于雌性的趨勢。王海建等通過實驗對西藏飛蝗糞粗提物中化學聚集信息素進行測量分析, 結果發現西藏飛蝗兩種生活方式下的西藏飛蝗聚集信息素組成成分相一致但所含比例有所不同。
3 西藏飛蝗的生態學特征
西藏飛蝗具有很強的遷徙能力, 以性成熟前的遷徙能力最強。唐昭華等研究西藏飛蝗成蟲的生物生態學發現成蟲的生殖活動、飛行能力、交尾、產卵等特性受到周圍生態環境的影響。昆蟲在長途遷徙過程中需要高營養的食物來維持身體所需消耗的能量, 高質量的食物會增加其成蟲的大小和繁殖能力。因此, 可以改變周圍的環境減少西藏飛蝗的活動范圍以降低為害程度。
3.1 西藏飛蝗的耐寒性
西藏飛蝗屬于變溫昆蟲。白天隨著溫度的升高其活動也頻繁, 夜晚隨著溫度降低其活動減緩。西藏飛蝗與其他昆蟲一樣體內存在一種過冷卻的生理現象, 西藏飛蝗的耐寒能力與過冷卻點呈負相關, 西藏飛蝗蟲卵的過冷卻點要比其他蟲態低, 即蟲卵耐寒能力更強, 更容易越冬。研究發現不同發育階段西藏飛蝗過冷卻點、結冰點如表2所示, 并發現在當地冬季西藏飛蝗的存在形式只有蟲卵一種。李慶等在測定西藏飛蝗的耐寒性理化指標時, 發現過冷卻點和結冰點不僅各發育階段的飛蝗之間存在差異, 而且群居型和散居型、雌雄之間也不相同。

表2 不同發育階段西藏飛蝗過冷卻點、結冰點
3.2 西藏飛蝗耐寒性的理化物質
西藏飛蝗的耐寒能力與其體內的生理生化物質的含量有密切關系, 各發育階段體內的理化物質含量如表3所示, 研究證明昆蟲的過冷卻點隨著體內的含水量的增加而降低, 隨著甘油的增加而增 加。西藏飛蝗體內抗寒性系統是以甘油等物質組成的, 為適應干燥缺氧的高原環境。在低溫脅迫的條件下, 其成蟲體內脂肪轉化為甘油, 糖原分解成糖類小分子碳水化合物, 體內的大分子蛋白質被降解成丙氨酸等氨基酸。西藏飛蝗體內的物質通過氧化分解釋放能量, 用來抵御低溫的影響, 維持生長發育、生理代謝所需要的正常溫度。
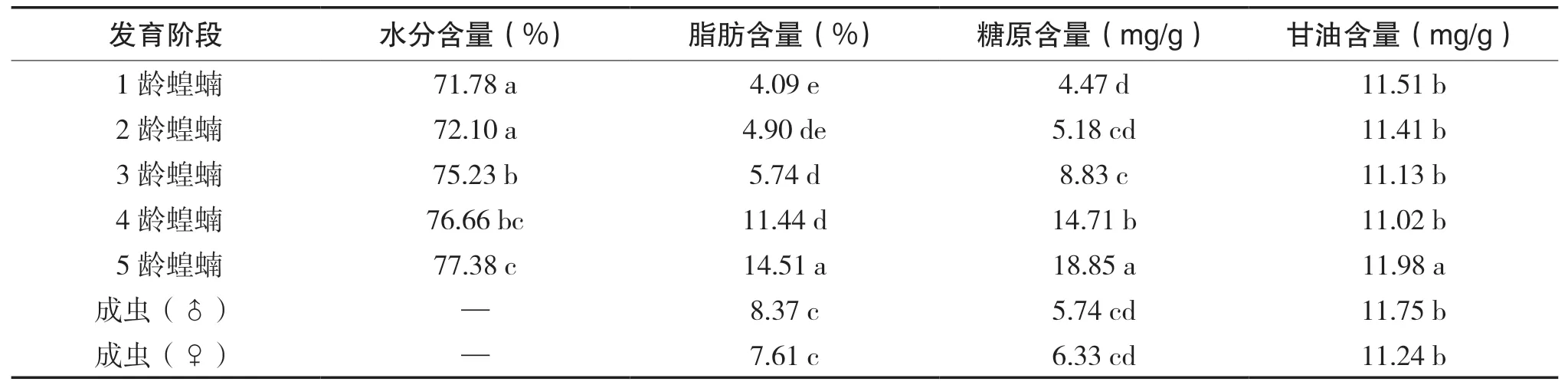
表3 西藏飛蝗(群居型)各發育階段體內的理化物質含量
4 西藏飛蝗受環境因子的影響
當前, 全球氣候還在不斷地升高, 未來全球的地表溫度也將繼續增加, 青藏地區作為氣候敏感區, 也會隨全球氣候的變化而變化。氣候的變化將會造成生態系統的不可逆的改變, 影響著西藏飛蝗的分布。由于溫度的升高導致西藏飛蝗代謝和繁衍能力都有所增強, 為害也隨之加劇。董兆克等分析了全球氣候變暖和局部氣候惡化不僅使昆蟲生存能力受到影響, 還能改變昆蟲的分布范圍, 是影響農作物和牧草等植物的病蟲害發生的重要原因之一。
4.1 西藏飛蝗受溫度的影響
西藏飛蝗的分布區域廣泛且海拔差異顯著, 是通過長期的適應和物種進化建立的新亞種群。溫度的變化將影響西藏飛蝗生長發育的改變, 有研究證明在一定的溫度范圍內, 西藏飛蝗各齡期的發育歷期與溫度呈負相關, 發育速率與溫度呈正相關, 具體見圖1 。在不同的溫度條件下, 其生長發育的全世代歷期會隨著溫度的升高而縮短, 說明溫度的升高將會加快西藏飛蝗的發育, 具體見圖2。吳雷等研究環境因子對西藏飛蝗產卵的影響, 發現西藏飛蝗體內的超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)的活性在低溫脅迫和高溫脅迫下會發生變化來應對低溫和高溫的環境影響;同時, 溫度的脅迫還會對丙二醛(MDA)含量影響進而影響生理代謝。韓政等分析了熱激蛋白與昆蟲的耐熱性關系時證實熱休克蛋白(HSP)是在不良環境下產生的應急蛋白, 控制應急蛋白的基因在個體受到不良環境的刺激后會轉錄表達, 從而提高昆蟲的抗逆性。通過對比西藏飛蝗在不同的溫度脅迫下的基因表達情況, 發現高溫脅迫和低溫脅迫條件下體內的轉錄基因組的表達量不同;低溫下的西藏飛蝗體內的幾丁質酶(CHT)活性更強, 從而提高西藏飛蝗的抗逆性能力。

圖1 西藏飛蝗各蟲態在不同溫度條件下的發育歷期
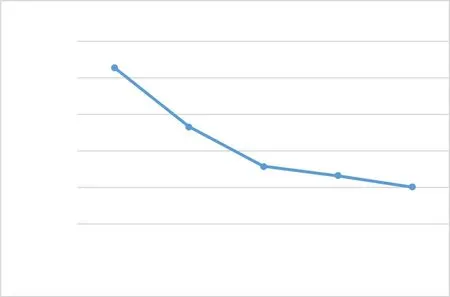
圖2 西藏飛蝗在不同溫度下的全世代歷期
4.2 西藏飛蝗受光照的影響
吳蕾用西藏飛蝗做實驗, 通過不同的紫外線輻射研究光照對西藏飛蝗的分布、分化和擴散, 發現用不同紫外線輻射后蝗蟲體內SOD、POD和CAT 活性以及MDA含量有明顯變化。西藏飛蝗能夠在光照逆境的條件下, 身體內的自由基清除系統和體內的抗氧化酶協調作用形成抗逆機制, 適應光照引起的環境變化。在測量不同光照下西藏飛蝗體內的過氧化物丙二醛(MDA)含量的實驗, 說明蝗蟲體內的過氧化反應與光照的時長、光的種類都有一定的聯系。西藏飛蝗能夠在青藏高原干燥的環境下生存, 是因為體內的抗氧化酶活性、含量和比例適應環境發生了改變。
4.3 西藏飛蝗受低氧的影響
西藏飛蝗的生存離不開氧氣(O), 當O過低時會導致組織氧化損傷。為適應青藏高原含氧量低的環境, 西藏飛蝗的呼吸模式中存在不連續氣體交換循環(DGC)呼吸模式。研究表明DGC呼吸模式與昆蟲呼吸失水性和缺氧的環境中進化有關;西藏飛蝗通過增加DGC呼吸速率吸收更多的氧氣(O)釋放出二氧化碳(CO)。鄧曉悅等通過測量和計算西藏飛蝗與東亞飛蝗的呼吸特征發現西藏飛蝗可以縮短DGC開放階段, 用來補償DGC頻率增強帶來的呼吸性水(HO)的損失。進一步證明西藏飛蝗的DGC呼吸模式與高原環境有關。
5 西藏飛蝗的綜合防治措施
蝗災被列為我國農業上的三大自然災難之一, 嚴重影響我國的農牧業經濟發展。在我國分布有三個種群的蝗蟲, 其中西藏飛蝗主要分布在青藏高原地區, 為害牧草和禾本科農作物。我國對蝗蟲的防治工作從以前的迷信、盲目治理到科學、安全、有效地防治, 到現在的生物技術防治、農業綜合防治, 堅持植物資源保護、生物資源保護、環境資源保護以及生態資源保護的理念, 開發綠色可持續防治技術。
5.1 監測技術研究進展
西藏飛蝗的遷飛能力強, 種群規模大, 導致在監測上造成很大的問題。20世紀初, 我國通過鑒定蝗蟲種類、研究蝗蟲的生長發育以及調查蝗蟲分布區域結合氣象因子分析蝗蟲發生規律從而建立檢測站觀測。隨著我國計算機技術的快速發展, 對蝗蟲的檢測技術有很大的進步, 在20世紀末我國開始利用遙感(RS)檢測蝗蟲, 利用地理信息系統(GIS)預測蝗蟲發生期和發生地點, 預防蝗災。到21世紀, 我國開始將野外調查和3S技術相融合, 雷達技術、高空燈和地面誘捕相結合;利用大氣環流建立我國蝗災的檢測預警體系, 實現蝗災的提前預測并制定相對完善防治措施。
5.2 防治技術研究進展
針對西藏飛蝗的發生規律和為害癥狀結合生態安全建設, 防控技術從以前單一的物理防治或者化學防治應急防治策略發展到現在的以生物防治和農業防治多項防控技術相結合的綠色可持續防治體系。防治技術的不斷改進與完善體現了人們想要控制西藏飛蝗的種群數量來調節西藏飛蝗的發生頻率, 減輕西藏飛蝗對農業和畜牧業的影響。隨著社會的進步, 人們在實踐中挖掘更經濟、環保的防治措施。
在我國傳統防治蝗蟲技術中一般人們通過飼養雞、鴨或者其他蝗蟲天敵來捕食蝗蟲, 一直從古延續至今, 具有綠色、環保的優點, 但當大面積發生蝗蟲時, 無法得到有效控制。經過人們多年的嘗試, 現在我國對蝗蟲的防治措施是采用常用的化學藥劑對蝗蟲進行防治, 這是蝗災發生時首選的防治手段, 具有見效快、成本低、防治效果好等優點。由于單一追求防治效果嚴重引起了環境污染, 化學藥劑使用方法技術不普及, 導致化學藥劑在農戶手中不能夠正確使用, 造成水污染、大氣污染、土壤板結等環境問題, 導致區域生態失衡, 物種多樣性驟減。化學藥劑的長期使用, 不但增加西藏飛蝗的抗藥性, 而且化學藥劑具有難降解的特性, 會經生物鏈和生物富集進入人體, 嚴重損害人體健康, 甚至有些化學藥劑參與地球化學物質循環, 嚴重破壞地球生態系統。尋求一種綠色、環保、安全、高效的防治技術是目前綠色可持續防治蝗災的首要任務。
21世紀以來, 我國對害蟲的防治遵循不破壞生態環境和生物多樣性的原則, 對傳統的防控技術進行改進。在探索過程中對生態環境友好的生物防治技術登上歷史舞臺, 因為該防治方法對蝗蟲不產生抗體, 且具有防治效果持久等特點。早在20世紀90年代, 蝗蟲生物防治技術主要開發寄生性天敵的應用和植物源農藥的開發利用兩種主要形式, 通過人們不懈努力研發出一系列的殺蝗細菌、殺蝗真菌、殺蝗病毒、蝗微孢子蟲、蘇云桿菌等病原微生物菌劑, 多項殺蝗劑的研發在蝗蟲的生物防治中起到十分重要的作用。隨著研究的深入, 人們發現植物次生代謝物具有間接或直接的殺蟲性, 有希望替代化學藥劑防治蝗蟲, 推動蝗蟲生物防治, 將低耗、多元化、綠色的發展方向與生物制劑、化合物制劑、微生物制劑、天敵生物制劑等協同應用, 實現西藏飛蝗的綠色可持續控制。
在我國提出“預防為主, 綜合防治”的植保方針以后, 西藏飛蝗得到有效控制, 生態環境破壞也得到緩解。隨著“綠色可持續防控害蟲”技術的提出, 西藏飛蝗的綠色防控技術也在不斷開發。目前在明確西藏飛蝗發生規律與植被物候期的關系的前提下, 利用現代發達的監測預警技術研究飛蝗種群遷徙路線及規律, 從而建立短期、中期及長期的實時監測預警體系。保證生態安全的前提下, 建立集多項技術的綠色可持續防控技術體系。深度研究西藏飛蝗與寄主、非寄主、天敵之間的關系, 利用自然界存在的“推—拉策略”對西藏飛蝗防控符合我國目前科學的植保方針。
6 結語
西藏飛蝗能夠適應全球氣溫變暖, 所以加強對西藏飛蝗實施綠色可持續防治技術依然是一項持續的課題。在青藏高原地區必須大力發展以生態治理為核心的環保控蝗技術, 利用RS、GIS、GPS等先進技術監控西藏飛蝗的遷徙。結合西藏自治區生態系統保護政策和當地的民俗建立應急防治策略和可持續治理策略相結合防控技術。利用現代科技研究產卵聚集信息素、聚集信息素和昆蟲生長調節劑等干擾蝗蟲的生命活動, 防止西藏飛蝗種群暴發。研究寄主植物中的化學物質對西藏飛蝗的趨勢或非寄主植物中化學物質的避勢, 為西藏飛蝗的綠色可持續防治探索新的方向。
