基于合成生物學策略創制微生物天然產物
2022-09-27 11:01:54梁淑芳
四川師范大學學報(自然科學版) 2022年5期
梁淑芳
(四川大學 生物治療國家重點實驗室, 四川 成都 610041)
合成生物學是綜合分子和細胞生物學、進化系統學、生物化學、信息學、數學、計算機和工程學等多學科技術體系發展起來的交叉新學科.作為21世紀初新興的生物學研究領域,合成生物學是在闡明并模擬生物合成的基本規律之上,基于標準化、去耦化和模塊化達到人工設計并構建新的、有特定生理功能的生物系統,從而建立藥物、功能材料或能源替代品等的生物制造途徑.隨著基因組測序技術的飛躍發展,大量微生物、真菌甚至植物的基因組序列測序完成并公開共享,這也推動了合成生物學在微生物領域的應用.一系列微生物遺傳操作工具和技術方法被不斷發展和優化,進而靶向改造現有的微生物天然產物生物合成途徑,甚至創造潛在的有特定生理功能的生物合成途徑,最終實現目標產物的高效制造[1].
微生物源天然產物不僅包括抗生素(如鏈霉素、達托霉素)[2]、免疫抑制劑(如雷帕霉素)[3]、抗真菌藥(如哈霉素)[4]、抗腫瘤藥物(如博萊霉素、紫杉醇)[5]等醫療藥物,還有能源、農業抗蟲劑、食品、芳香劑等用途[6-7].微生物因為擁有較快的生長速度、較高的生產效率、簡單的遺傳操作而被廣大研究者關注和研究.除此之外,微生物還擁有豐富的輔因子、天然產物前體代謝產物及復雜的翻譯后修飾系統.因此,微生物被不斷改造以便提供最優的天然產物合成途徑.部分微生物,如大腸桿菌、酵母、枯草芽孢桿菌、鏈霉菌等,因其廣泛的碳源利用能力、復雜的翻譯后修飾系統、高效的分泌系統而被不斷改造成工程菌,為高效生產高附加值的天然產物提供良好的平臺[8].隨著人工智能的快速發展,機器人技術和集成軟件也被廣泛應用于自動化標準工作流程,從而促進合成生物學工作效率的顯著提升,進而推動天然產物發掘工作的飛躍發展.
本文對合成生物學在微生物領域的最新研究內容及應用案例進行了概述,并對提升微生物天然產物產量的合成生物學策略和工具進行了總結和討論,還對人工智能應用于微生物天然產物產量的提升和發掘進行了總結和展望.
1 設計構建微生物天然產物的合成路徑
1.1 理性設計天然產物生物合成通路通過文本挖掘工具進行文獻挖掘、通過網絡模型進行模型預測、通過合成生物學進行人工途徑合理設計重構,來探索和設計有效生產所需化合物的合成途徑(圖1A).生物學研究前沿和路徑設計中數據源的積累仍然記錄在文獻中,這些文獻提供了大量的生物學信息.文獻挖掘已經從簡單的術語識別發展到相互作用關系的綜合分析,并從蛋白質相互作用的單一認知擴展到路徑合作的系統調控.一方面,建立了以Pubmed和Medline為主的海量文獻高效處理平臺;另一方面,一系列的文本挖掘工具,如Textpresso[9]、LitMiner[10]等,不僅可以識別、提取、整合和分析文獻數據,還能發現新的、隱藏的或不可預測的信息.
隨著基因組學及相關技術的發展,大量的微生物基因組序列分析解讀用于產生高質量的基因組規模的代謝重建,可用于產生綜合代謝模型.因此,一系列數據庫隨之興盛,為微生物代謝譜解析及代謝重構提供參考[11].其中,antiSMASH作為一個網絡服務器和獨立的工具,已成為鑒定和分析細菌、真菌及植物基因簇最廣泛使用的工具[12].除此之外,BIGG作為高質量基因組規模代謝模型的集中存儲庫,已完成了165個代謝模型(http://bigg.ucsd.edu),涵蓋78種微生物[13].這些基因組規模代謝模型的應用從理論研究到實用研究都是為了彌合基因型和表型之間的差距,其收錄的代謝產物及代謝反應依然為微生物的代謝重建提供參考.總之,通過文獻調研、生物信息學分析、計算機建模等,合理設計科學、可行的生物合成通路.
1.2 構建和優化生物合成通路大多數次級代謝產物生物合成基因簇在常規培養條件下是轉錄沉默的,不利于天然產物的表達積累[14].因此,激活沉默基因簇是發現新活性天然產物的有效途徑之一.天然產物的生物合成通常是包含多種基因及其控制元件完成的多步驟途徑,對這些基因或操控元件進行改造優化,可實現天然產物產量的提升或新產物的發掘.基因表達的精確調控是天然產物產生的關鍵步驟,如通過對影響產物合成過程中關鍵途徑關鍵酶的活性或表達豐度的提升以最大限度地合成目標產物.啟動子、轉錄因子工程是實現基因精準調控的重要途徑,模塊化工程則是實現目標產物修飾的高效途徑之一(圖1B).
一方面,內源性啟動子由于不能及時控制和持續最大化細胞內的轉錄水平而受到限制,通過位點選擇突變增強啟動子強度,或以核心啟動子區域和上游激活序列為重點構建啟動子文庫,篩選高活性啟動子[15].總之,啟動子工程作為合成生物學不可或缺的一部分,是調節遺傳回路和協調生物合成途徑的關鍵因素.另一方面,因為啟動子只能精準控制一個元件或基因,所以存在一定局限性.而轉錄因子因能同時控制代謝途徑中多種酶的豐度或活性廣泛應用于提高目標產物的產量[16].研究人員在不斷尋找高效正調控因子的同時,對已有轉錄因子的結構域或模塊進行編輯或融合來顯著提高轉錄因子的活性[17].另外,轉錄因子誘餌通過模擬與負調控因子結合從而阻止負調控因子結合相關DNA靶點,以此來干擾基因調控,實現天然產物合成通路的激活[18].
此外,基于遺傳操作優化生物合成通路,從而產生目標化合物的各種衍生物也成為篩選活性天然產物的重要途徑之一.Kudo等[19]基于CRISPR-Cas9及Gibson組裝技術對雷帕霉素(rapamycin)合成基因簇中的模塊進行刪除及替換,從而獲得一系列活性較好、毒性較低的rapamycin衍生物.
1.3 生物合成通路異源高效表達除了通過優化生物合成途徑提高天然產物合成效率,提高微生物發酵的產率也很重要.許多微生物內源次級代謝產物積累的水平不能滿足工業化生產的需求,是因為部分天然產物在本宿主中沒有充足的碳氮源、前體或代謝中間產物的供給來滿足目標代謝途徑的需求,因而產量極低.相反,生產工程菌在經過一系列的基因組優化、轉錄調控、蛋白質工程和代謝工程優化后,有著高效的調控系統、高性能的催化系統及豐富的前體和輔因子,還擁有最優的流向目標化合物同時降低或消除競爭途徑消耗的代謝流[20].因此,適宜規模化生產的工程菌為可再生和可持續的化工替代品的生產提供了高效環保的細胞工廠(圖1C).

圖1 合成生物學技術挖掘微生物天然產物
工程菌在經過一系列的靶向代謝重編程后,其代謝流集中流向目標化合物.因此,眾多天然產物合成基因簇在轉入工程菌后,其產量被提高數倍甚至數千倍[21].例如,將灰紫紅菌素(griseorhodin)生物合成通路轉入變鉛青鏈霉菌(Streptomyceslividans,S.lividans)ΔYA10工程菌后,其產量比野生型提高了5倍;而在轉入白色鏈霉菌(Streptomycesalbus,S.albus)Del14底盤細胞后,其產量更是提高了11倍[22].不僅如此,越來越多的植物天然產物也嘗試構建微生物工程菌生產,大大縮短了生產周期并提高了產量[23].例如,青蒿素、人參皂甙在轉入糧酒酵母工程菌后,其產量都有了質的飛躍[24-25].
2 幾種合成生物學有關的技術
2.1 基因編輯技術源源不斷的基因組信息促進了一系列基因組編輯工具的發展,而這些不斷更新的DNA編輯工具則進一步促進了沉默基因簇的重構和激活[26],最終提高目標產物的產量.目前,聚類規則間隔短回文重復序列(Clustered regularly interspaced short palindromic repeats, CRISPR)/CRISPR相關蛋白(Cas)系統廣泛應用于眾多生物體的基因功能研究.CRISPR/Cas9系統在引導RNA(Guide RNA,gRNA)的介導下,靶向基因組DNA,并招募Cas9蛋白對靶DNA進行切割形成雙鏈斷裂(Double-strand break,DSB),并以外源導入的同源DNA為模板,通過體內同源重組酶進行同源修復(Homology-direct repair,HDR),從而達到基因敲除、插入及突變的目的(圖2A).
CRISPR/Cas9系統極大地促進了對微生物高效率的基因組編輯[27],但即使在減少gRNA的情況下,Cas9毒性仍然存在,并且導致Cas9逐漸丟失[28].因此,其他基因組編輯工具如Cas12a等也在不斷被開發優化[26],以期獲得更高的基因編輯效率和更低的毒性.通過失活Cas9的核酸內切酶活性而獲得dead Cas9(dCas9)[29].dCas9應用于無DSB的、單核苷酸刪除的基因組編輯系統.dCas9連接DNA脫氨酶以進行基因編輯,該系統包括一個胞苷和一個基于腺苷脫氨酶的堿基編輯器(Base editor),利用dCas9-gRNA復合物發現并結合目標DNA,胞苷脫氨酶通過胞苷脫氨基作用將目標C轉化為U(圖2B).類似地,該系統也能在約7個核苷酸框內將A:T堿基對轉化為G:C堿基對[30].
文獻[31]還發展了將多種嵌合的dCas9-DNA脫氨酶融合進行精確的基因編輯和位點特異性突變的生成,然后用于選擇耐藥表型.堿基編輯器降低了DSB對染色體的壓力和Cas9的毒性,除了引入終止密碼子使基因失活外,還具有巨大的應用潛力,如通過糾正不理想的點突變或使假基因恢復功能狀態,通過在體內交換關鍵殘基進行蛋白質工程改造,以及通過在一個結構中復用gRNAs來進行全路徑工程優化[30].總之,堿基編輯器是對CRISPR-Cas基因組編輯工具技術的補充.
除此之外,將dCas9與轉錄增強子(如RNA聚合酶因子σ因子、bEBPs結合蛋白、AsiA)結合,在gRNA的介導下靶向目標序列,從而使轉錄增強子增加目標基因的轉錄水平[32](圖2C).類似地,失活的dCas9在gRNA的介導下準確地靶向基因組DNA后,與dCas9形成復合物,在空間上阻止RNA聚合酶的進展并抑制目標基因的轉錄,從而達到抑制基因表達的效果(圖2D).目前,dCas9與不同的酶活性蛋白聯系起來,從而廣泛應用至轉錄調控、表觀遺傳修飾和熒光基因組追蹤等[33].
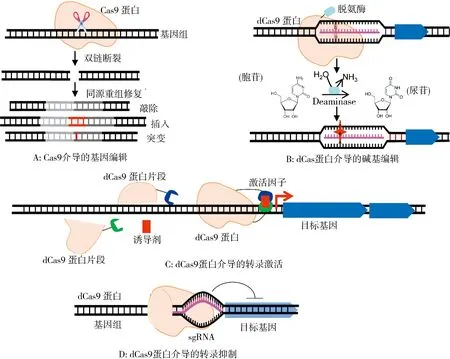
圖2 CRISPR/Cas9基因編輯系統
2.2 基因簇克隆技術在一個可遺傳的宿主中異源表達單個基因、一串基因甚至整個生物合成基因簇是挖掘相應天然產物的一種有效途徑[34].為了將
這些天然產物生物合成基因簇引入異源宿主,快速獲取這些基因簇都是必不可少的.獲取生物合成基因簇的方法可歸結為2種方法:一是直接克隆方法,二是體內外重構方法.
直接克隆方法中經典且廣泛應用的是篩選基因組文庫,即通過限制性內切酶酶切基因組,將基因組片段化,隨后將基因組片段克隆至能承載大基因組片段的載體,如細菌的BAC載體系列和P1噬菌體的PAC載體系列[35]、大腸桿菌-鏈霉菌穿梭質粒pStreptoBAC V[36],來獲得目的基因簇.BAC載體系列是基于大腸桿菌F-可繁殖質粒,能容納長達300 kb的基因組大片段,使其適合于大多數基因簇的捕獲[37].通過這種方法可建立一個基因組文庫,并可通過高通量篩選新化合物(圖3A).然而,這種基于限制性內切酶的基因組消化,沒有靶向性,即使有高通量篩選助力,也增加了研究者的工作量.
此外,部分直接克隆方法是基于整合酶介導的位點特異性重組來克隆獲得目的基因.Du等[38]開發了一種φBT1整合酶介導的位點特異性重組系統,并應用該系統克隆了鏈霉菌中長達157 kb的基因簇.盡管該方法可抓取大的DNA片段,獲得較高的克隆效率,然而,該方法需要經過一輪單交叉和一輪雙交叉,耗時耗力.文獻[39]結合限制酶和RecET重組(exonuclease combined with RecET recombination,ExoCET)直接將細菌和哺乳動物基因組中任何選擇的區域克隆到目標質粒中(圖3B).ExoCET結合了體外核酸內切酶和退火以及全長RecET同源重組(HDR)從基因組DNA制備物中提取特定區域.該方法結合限制性內切酶和同源重組,靶點明確,顯著降低盲目性,且目標基因簇及載體借助RecET同源重組酶在大腸桿菌中進行同源重組,一周內就可完成一輪基因簇的克隆抓取,顯著縮短工作周期,提高工作效率,但該方法往往受限制性內切酶限制.因此,該團隊隨后嘗試結合CRISPR-Cas9酶切和RecET同源重組酶進行基因簇抓取克隆,然而這個嘗試并沒有獲得良好的克隆效率,甚至效率很低[40].
目前也開發了多種體內外DNA組裝工具以便高效、快速地重構優化生物合成通路[41](圖4).Gibson assembly及其他一些基于重組酶的體外重組、體內酵母重組[42],在生物合成通路重構方面都取得好的進展.其中,基于5′外切酶、DNA聚合酶和DNA連接酶協同作用的Gibson assembly,已被用于組裝高達900 kb的DNA片段[43](圖4A).體內重組,主要是基于酵母及大腸桿菌體內的重組酶介導具有相同同源臂的DNA片段連接成環狀DNA(圖4B).iCOPE則是基于Cas12a突變體及其gRNA、T4連接酶、可在一個反應中編輯或組裝DNA片段的方法.Cas12a突變體擁有廣泛的識別位點,在體外轉錄的一對相應gRNA的引導下,在特定位點上切割靶DNA,隨后在T4連接酶作用下,將具有相同黏性末端的DNA片段鏈接,從而將DNA片段組裝成環狀DNA(圖4C).iCOPE不僅高效,且在操作大質粒方面也很有前景[44].
隨著DNA組裝工具的不斷發展,開發了眾多體外重組試劑盒且應用廣泛.酵母作為高效且廣泛應用的體內重組系統,用于天然產物合成通路的重組及優化.如基于酵母重組,利用即插即用方法激活多個沉默天然產物基因簇[45].除了將基因簇各元件擴增后再重組而激活目標基因簇,在原始基因組或者直接克隆的帶有目標基因簇的載體上進行修飾進而優化通路,同樣可以達到激活基因簇的目的.除此之外,通過在宿主基因組的主要結構基因前插入強啟動子、激活通路特異性正向調控因子等措施同樣可以有效提高天然產物的產量[46-47].
2.3 酶工程蛋白質是生命活動的主要承擔者,在自然界中發現的蛋白質傳統上是最常用的生物催化劑,用于生產從商品化學品到藥物等多種天然產物.酶工程在代謝工程的發展中,特別是在天然產物的生物合成中,已經成為一個強大的生物技術工具箱.近年來,酶工程已成為天然產物生物合成中提高酶活性、提高酶穩定性、擴大產物譜的一種有效方法[48].
從蛋白活性方面入手激活天然產物.天然產物生物合成的主要瓶頸之一是涉及的催化或合成酶活性有限.將外源酶整合到微生物底盤中可能會降低其催化活性,甚至導致功能喪失.提高酶活性以加速生產過程是這一領域的主要目標.通過隨機突變提高酶對特定底物的催化活性是蛋白質工程中使用的典型策略.如通過隨機突變,釀酒酵母異戊烯基二磷酸異構酶的催化活性提高了2.53倍,從而使番茄紅素產量(1.24 g/L)比野生型提高2.1倍[49].然而,這種隨機突變策略產生的酶庫通常是巨大的,而且大多數變異體已降低到無活性,使得篩選工作耗時耗力且效率低下.因此,首選的方法是分析目標酶的結構,并在結合位點或活性域附近設計位點定向突變,以提高其催化活性.
酶活性固然重要,酶穩定性依然不可忽視.酶的穩定性是多方面的,包括熱穩定性、pH耐受性、溶解度以及對鹽和有機溶劑的耐受性.提高熱穩定性可以使酶在高溫下發揮作用[50].分子分析表明,熱穩定性的提高是由于表面電荷增加和結構柔韌性降低所致[51].在結構導向一致性概念的指導下,葡萄糖1-脫氫酶的熱穩定變體在65 ℃下產生并顯示出極大的半衰期改善(3.5 d),后來發現它也能耐受高濃度的鹽和有機溶劑[52].酶也被設計來適應所需的環境pH值.例如,在木質素降解中起主要作用的漆酶,喜歡更高的溫度和pH值.一種來自Botrytis aclada的真菌漆酶經過定向進化獲得了更高的最適pH值,在中性pH值下其活性增加了5倍[53].
3 人工智能加速微生物天然產物創制
合成生物學技術大大提高了發掘創造活性化合物的速度.隨著人工智能的快速發展,機器人技術和集成軟件也被廣泛應用于自動化標準工作流程,從而使合成生物學工作的效率得以顯著提升.合成生物學是由多種非線性工作流程組成的“按需制造”,而生物學實驗存在異質性和動態性,使得工作流程因批次間的變化而更加復雜[54].
近年來,生物鑄造廠(Biofoundries)已經發展成為自動化設計—建造—測試—工程周期一體化,解決了生物工程過程中緩慢、昂貴且重復性低的幾大難題[55].已有機器人系統結合機器學習算法的應用,該應用可完成完全自動化生物系統的設計、構建、測試和學習過程[56].這個全自動的機器人平臺BioAutomata評估了不到1%的可能變異,比隨機篩選高出77%,并且研究者利用機器人平臺優化了番茄紅素生物合成途徑,提高其產量.
4 展望
在過去的20年,全基因組測序的突破揭示了微生物尚未開發的生物合成潛力,從而奠定了深入研究微生物和挖掘微生物天然產物的基礎.蛋白質組測序則有助于找出微生物應對體內外刺激所做出的生化反應,從而為研究和揭示相關蛋白功能及生化過程提供重要參考[57-59].利用多組學數據和基因組規模的代謝網絡模型進行系統分析,預測代謝工程的目標,并重新設計代謝通量,以最大限度地提高目標天然產物的產量[58,60].新的分子操作工具如DNA組裝、重組和基因組編輯技術的發展減少了克隆和編輯基因簇所需的時間,提高了克隆和編輯基因簇的效率.新技術的出現也促進了異源宿主的工程化,從而使天然產物在遺傳上易于產生.此外,通過合成基因的模塊進行重新編程,可以生產出新的天然化合物衍生物,擴展天然產物的生物活性.
對天然或改進的基因進行模塊化重構,無論是天然產物修飾還是產生新分子,都是一種非常有效的方法.這些技術的集成將有助于我們理解復雜的基因調控網絡和代謝網絡,從而通過基因簇編輯、通路改造、代謝通量優化、酶工程、調控電路重連和宿主修飾來發現和生產新的天然產物.隨著知識和信息的增加,天然產物“谷歌地圖”也將隨之產生,類似于Lee等[61]報道的“生物化學地圖”,突出了單個或多個生化反應的策略和途徑,為設計和生產特定的感興趣的天然產物提供重要依據.
近年來,人工智能技術和集成軟件也逐漸拓展到生命科學研究.機器人的應用,避免了人為因素導致的誤差或低效率,在節約成本的同時,快速精準地完成項目內容.盡管目前還未廣泛應用機器人等人工智能系統,但隨著人工智能的快速發展,相信在不久的將來,人工智能系統會被廣泛應用于天然產物的發掘和生產,更多更優的天然產物來源產品更快地問世,以滿足人們的需求.
總之,代謝工程和合成生物學技術的進展將為發現和生產多種有價值的活性化合物提供強大的解決方案,并有望開啟天然產物發掘的新黃金時代.合成生物學技術及產品因其高效性、精準性已被廣泛應用于市場,其對人口健康、生物醫藥等領域將產生深遠影響[62-63].
致謝本文得到四川大學生物治療國家重點實驗室和四川省人民醫院張丹博士在圖片構思編輯上的大力協作,在此一并致謝.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
現代畜牧科技(2021年4期)2021-07-21 06:13:00
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
數學物理學報(2017年2期)2017-06-05 09:12:30
