大鼠陰莖海綿體內(nèi)皮細胞的分離培養(yǎng)與鑒定*
2022-09-18 15:15:48瑾阿爾孜古麗吐爾遜劉鳳霞阿地力江伊明斯依提阿木提侯鵬程
中國男科學(xué)雜志 2022年4期
關(guān)鍵詞:生長
靳 瑾阿爾孜古麗吐爾遜劉鳳霞阿地力江伊明斯依提阿木提侯鵬程
1.新疆醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院人體解剖學(xué)教研室(新疆烏魯木齊830011);2.新疆醫(yī)科大學(xué)第一附屬醫(yī)院臨床研究院
陰莖海綿體組織主要由陰莖海綿體內(nèi)皮細胞(corpus cavernosum endothelial cells,CCECs)、平滑肌細胞等組成[1]。 在勃起過程中,CCECs 和平滑肌細胞在調(diào)節(jié)海綿體竇血流充盈和維持血管張力中起關(guān)鍵作用[2]。 內(nèi)皮功能受損是導(dǎo)致勃起功能障礙(erectile dysfunction,ED)的重要原因之一,不同類型ED 的研究結(jié)果提示,陰莖海綿體組織均存在不同程度的內(nèi)皮細胞功能改變和平滑肌纖維化現(xiàn)象[3-5],而在血管性ED的發(fā)生發(fā)展中,CCECs 的損傷更是造成ED 的主要危險因素[6]。 以往在ED 的體外研究中,通常以人臍靜脈內(nèi)皮細胞作為研究對象,但由于生長于不同器官和血管的內(nèi)皮細胞具有異質(zhì)性,其表型、功能、代謝特征等方面均有所不同[7],因此無法準(zhǔn)確反應(yīng)CCECs 在ED 的發(fā)病過程中所發(fā)揮的作用。 而CCECs 覆蓋于基質(zhì)表面形成陰莖海綿體竇成為陰莖組織中獨特的血管床[8],而血管損傷引起的ED 通常還作為全身內(nèi)皮功能障礙的指標(biāo),甚至還有可能是心血管疾病的預(yù)警信號[9],因此,探索原代培養(yǎng)CCECs 的方法,將更有助于研究血管性ED 等疾病的發(fā)病機制,為后續(xù)體外ED 模型制備及藥物實驗提供新載體。
資料與方法
一、資料
(一)實驗動物
SPF 級性成熟期雄性Sprague-Dawley 大鼠,體重(250±10)g,購于新疆醫(yī)科大學(xué)實驗動物中心。 該實驗在處理動物過程中,嚴(yán)格遵循新疆醫(yī)科大學(xué)實驗動物倫理章程。
(二)主要實驗儀器
CO2培養(yǎng)箱(美國Thermo 公司);高速臺式低溫離心機(德國Eppendorf 公司);酶標(biāo)儀(美國Bio-Rad 公司);熒光倒置顯微鏡(日本OLYMPUS 公司);激光共聚焦(德國Leica 顯微圖像采集系統(tǒng));精密電子天平(德國sartorius 公司);-20℃冰箱(中國海爾公司);培養(yǎng)瓶及培養(yǎng)皿(美國康寧公司)。
(三)主要試劑
EGM-2(微血管內(nèi)皮細胞毛長介質(zhì)-2)內(nèi)皮細胞生長培養(yǎng)基(CC-3162,Lonza,瑞士)、胎牛血清(04-001-1ACS,BI 公司,以色列)、抗CD31 抗體(ab24590,Abcam,英國)、抗VWF 單克隆抗體(AF3000,Affinity Biosciences,美國)、Cy3 ?山羊抗兔IgG H&L(ab6939,Abcam,英國)、FITC 山羊抗小鼠IgG H&L(ab6785,Abcam,英國)、Ⅱ型膠原酶(17101015,Gibco 公司,美國)、無菌PBS、0.25%胰蛋白酶-0.02%EDTA 消化液、青/鏈霉素雙抗均購自于美國Gibco 公司。
二、方法
(一)大鼠CCECs 的原代培養(yǎng)
SD 大鼠適應(yīng)性飼養(yǎng)1 周,2%戊巴比妥鈉麻醉,5min大鼠癱軟。 在無菌環(huán)境下,暴露大鼠陰莖及陰莖頭,沿包皮剪開,分離陰莖組織,緊貼陰莖根部取下陰莖組織后將大鼠進行脫頸處死。 將組織放入含2%青/鏈霉素的PBS中浸泡5min。 剪除陰莖頭和陰莖軟骨,分離筋膜、結(jié)締組織,剝離尿道海綿體、陰莖背深靜脈,分離陰莖海綿體外部白膜。 用含雙抗的PBS 反復(fù)沖洗海綿體組織后,剪成約1mm3的組織塊,移至5mL EP 管中,加入1mL 0.5%Ⅱ型膠原酶消化液,組織塊剪成肉糜狀,移入15mL 離心管內(nèi),加入組織塊3 倍體積的0.5%Ⅱ型膠原酶消化液,于37℃環(huán)境下進行消化,每隔15min 震蕩2min。 至組織塊完全消化后,加入EGM-2 完全培養(yǎng)液終止消化,無菌200目濾網(wǎng)過濾細胞懸液,1000rpm 離心6min,棄上清,加3mL PBS 吹打混勻,1000rpm 離心5min,棄上清,加入5mL EGM-2 完全培養(yǎng)基吹打混勻后接種于培養(yǎng)瓶,37℃、5%CO2的培養(yǎng)箱中培養(yǎng)。
(二)大鼠CCECs 的純化
原代培養(yǎng)48h 時首次換液,繼續(xù)培養(yǎng)至96h 時,鏡下觀察細胞并在培養(yǎng)皿底部標(biāo)記CCECs 細胞團位置,刮除其他部位的雜細胞團,PBS 清洗2 遍,加入EGM-2培養(yǎng)基繼續(xù)培養(yǎng)。
(三)大鼠CCECs 的傳代培養(yǎng)
待單層細胞融合至70% ~80%,1mL 胰酶消化,鏡下觀察貼壁細胞收縮變圓脫落后,加2mL EGM-2 完全培養(yǎng)液終止消化,1000rpm 離心5min,棄上清,PBS 清洗一遍,1000rpm 離心5min 棄上清,加EGM-2 完全培養(yǎng)基吹打混勻后,1 ∶2 進行傳代培養(yǎng)。
(四)HE 染色觀察大鼠CCECs 細胞形態(tài)
取第2 代細胞爬片后,4%多聚甲醛室溫固定細胞2h,預(yù)冷的1 ×PBS 洗5min×3 次,蘇木素染色90s,自來水沖洗10min,鹽酸分化1s,氨水反藍20s,自來水沖洗5min,伊紅染色90 ~120s,自來水沖洗5min,梯度酒精脫水透明,封片后進行鏡下采圖。
(五)繪制大鼠CCECs 生長曲線
取第2 代CCECs 制備細胞懸液,按照5×104個/mL的濃度均勻接種于96 孔板,每24h 取6 孔細胞加入10μL CCK8 溶液,37℃孵育4h 后在450nm 處讀取OD值,連續(xù)測量10 天,繪制CCECs 生長曲線。
(六)大鼠CCECs 免疫熒光鑒定
取第2 代CCECs 細胞爬片后,4%多聚甲醛室溫固定細胞2h,預(yù)冷的1 ×PBS 洗3 遍,滴加0. 5% Triton X-100 通透15min,PBS 洗5min ×3 次,滴加山羊血清,室溫封閉30min,傾去封閉液后即加入抗體vWF 和CD31,4℃孵育過夜,PBS 洗滌后加入熒光二抗,37℃避光孵育1h,室溫滴加DAPI (4',6-二脒基-2 苯甲吲哚)染核10min,PBS 洗5min×3 次,熒光倒置顯微鏡下觀察并采圖。
(七)流式細胞術(shù)檢測第1 ~4 代大鼠CCECs 的細胞周期變化
取1 ~4 代大鼠CCECs 細胞,按照1×106個/mL 濃度制備細胞懸液并在預(yù)冷的70%乙醇中4℃固定過夜,1000rpm 離心5min,棄上清,PBS 清洗后收集細胞沉淀,加 入100μg/mL 的 RNAnase 酶37℃孵 育30min,50μg/mL的PI(碘化丙定)染液避光孵育30min,200 目濾網(wǎng)過濾后上機檢測。
(八)激光共聚焦檢測第1 ~4 代大鼠CCECs 中vWF 的表達變化
取1 ~4 代大鼠CCECs 細胞,方法步驟同(六)進行免疫熒光染色,激光共聚焦顯微鏡觀察第1 ~4 代內(nèi)皮細胞中vWF 的表達并采圖。
結(jié) 果
(一)倒置顯微鏡下觀察細胞培養(yǎng)結(jié)果
原代培養(yǎng)至第4 天時可在鏡下觀察到細胞團呈發(fā)散狀向四周生長,貼壁細胞呈短梭型、多觸角型(圖1A);培養(yǎng)至第7 天時鏡下可見貼壁細胞鑲嵌排列于同一平面,呈典型的“鋪路石樣”成團生長(圖1B);培養(yǎng)至第11 天時,可見細胞融合到70%左右,隨細胞數(shù)目增加而形成單層鋪路石樣排列(圖1C),可進行傳代。

圖1 原代培養(yǎng)大鼠CCECs 結(jié)果
(二)HE 染色觀察細胞形態(tài)
HE 染色結(jié)果顯示,正常2 代細胞形態(tài)較均一,細胞單層排列,保持“鋪路石樣”外觀(圖2A),細胞呈多角形,核大而清晰,呈卵圓形居中,有較為明顯的2 ~4枚核仁(圖2B)。
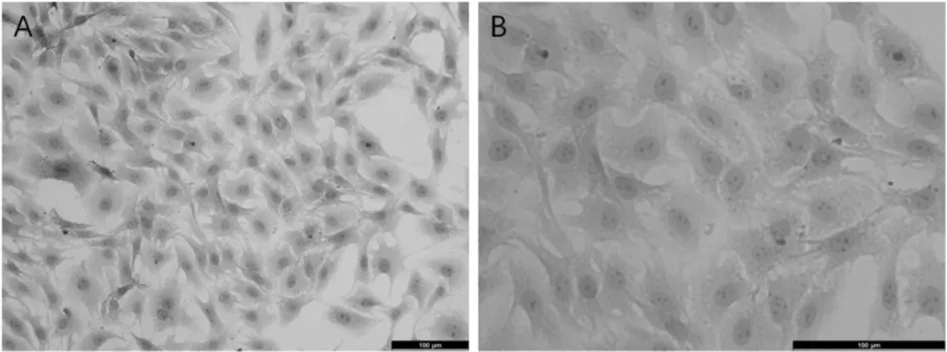
圖2 傳代培養(yǎng)大鼠CCECs HE 染色
(三)大鼠CCECs 生長曲線
第2 代內(nèi)皮細胞增殖速度明顯增快,生長曲線為“S”形,在細胞培養(yǎng)的第1 天細胞開始生長,但速度較慢;第2 ~4 天生長加快,進入對數(shù)生長期;第4 天后內(nèi)皮細胞增殖緩慢;第6 天開始細胞進入平臺期(圖3)。

圖3 大鼠CCECs 生長曲線
(四)vWF、CD31 免疫熒光染色鑒定結(jié)果
取第2 代細胞進行vWF 和CD31 免疫熒光共染,檢測內(nèi)皮細胞標(biāo)志物表達情況,結(jié)果顯示,vWF 在細胞質(zhì)中穩(wěn)定的表達為紅色熒光(圖4B),CD31 在細胞質(zhì)中穩(wěn)定的表達為綠色熒光(圖4C),通過分析,vWF 和CD31 免疫熒光共染的雙陽性表達率為(86. 19 ±1.14)%,符合內(nèi)皮細胞特征,提示大鼠CCECs 原代分離、培養(yǎng)成功。

圖4 vWF、CD31 免疫熒光染色結(jié)果
(五)第1 ~4 代CCECs 的細胞周期變化檢測結(jié)果
結(jié)果顯示,隨著傳代次數(shù)的增加,靜止期(G0/G1期)細胞數(shù)量逐漸增加,S 期(DNA 合成期)細胞數(shù)量依次減少,當(dāng)傳至P4 代時,細胞增殖周期逐漸阻滯在G0/G1 期(圖5),增殖速度減慢,提示,第2、3 代細胞生長周期穩(wěn)定,可用于后續(xù)實驗研究。

圖5 流式細胞術(shù)檢測細胞周期結(jié)果圖
(六)第1 ~4 代CCECs 中vWF 表達變化激光共聚焦檢測結(jié)果
結(jié)果顯示,vWF 穩(wěn)定表達于各內(nèi)皮細胞的細胞質(zhì)中,呈現(xiàn)紅色,同時可見在第2 ~3 代,內(nèi)皮細胞標(biāo)志物vWF 的表達較高,而在第4 代細胞vWF 的表達明顯降低,同時可見細胞形態(tài)不一,并出現(xiàn)細胞萎縮、胞質(zhì)邊界部位的形態(tài)學(xué)改變,提示第4 代細胞出現(xiàn)了衰老,并進而導(dǎo)致內(nèi)皮細胞標(biāo)志物vWF 表達降低(圖6)。 該結(jié)果與第1 ~4 代CCECs 的細胞周期變化結(jié)果相對應(yīng),進一步揭示了大鼠CCECs 的生長規(guī)律,并提示采用第2、3代細胞進行后續(xù)實驗可行。

圖6 傳代培養(yǎng)大鼠CCECs 中vWF 熒光結(jié)果
討 論
陰莖組織主要由一對陰莖海綿體和一個尿道海綿體構(gòu)成,在接收到性刺激信號時,CCECs 釋放一氧化氮(nitric oxide,NO)并作用于平滑肌細胞,激活可溶性鳥苷酸環(huán)化酶(solubleguanylatecyclase,sGC),增加細胞內(nèi)環(huán)磷酸鳥苷(cyclic guanosine monophosphate, cGMP)濃度,降低平滑肌細胞內(nèi)Ca2+濃度,使平滑肌松弛,血液經(jīng)海綿體動脈流入海綿體,在白膜的壓迫下,海綿體充血膨脹,發(fā)生勃起[10,11]。 NO/sGC/cGMP 作為促進陰莖勃起的經(jīng)典通路,NO 的持續(xù)穩(wěn)定輸出在信號通路傳導(dǎo)過程中至關(guān)重要[12]。 NO 作為許多細胞過程和生物學(xué)功能的重要信號分子,主要由三種一氧化氮合成酶(nitric oxide synthase,NOS)的同工酶,即內(nèi)皮型一氧化氮合酶(endothelial nitric oxide synthase,eNOS)、神經(jīng)型一氧化氮合酶(neuronal nitric oxide synthase,nNOS)和誘導(dǎo)型一氧化氮合酶(inducible nitric oxide synthase,iNOS)催化產(chǎn)生[13]。 iNOS 作為鈣非依賴性的NOS,其誘導(dǎo)的NO 的生成量主要是在轉(zhuǎn)錄水平上控制的[14],nNOS 催化產(chǎn)生的NO 受神經(jīng)傳遞信號的調(diào)控[15],而在內(nèi)皮細胞中,eNOS 在將L-精氨酸轉(zhuǎn)換為L-瓜氨酸的反應(yīng)中產(chǎn)生NO,因此,內(nèi)皮功能主要基于eNOS 的功能和活性[16]。 當(dāng)CCECs 結(jié)構(gòu)受損時,NO 的產(chǎn)生減少,進而可導(dǎo)致ED 的發(fā)生。 在我們以往開展的糖尿病性ED、復(fù)合應(yīng)激性ED 等相關(guān)研究結(jié)果均顯示,海綿體內(nèi)皮細胞均存在不同程度的功能損傷和平滑肌纖維化現(xiàn)象[17,18],亦有報道認為[19,20],高血壓、心血管疾病,甚至新冠肺炎患者發(fā)生ED 的原因均與陰莖海綿體內(nèi)皮功能受損相關(guān),上述結(jié)果均表明血管內(nèi)皮系統(tǒng)的完整性是正常勃起功能所必需的,CCECs 在陰莖勃起的過程中占有不可或缺的地位[21],更由于我們在后續(xù)研究中發(fā)現(xiàn),在復(fù)合應(yīng)激ED 的發(fā)生發(fā)展過程中,氧化應(yīng)激、慢性炎癥和神經(jīng)內(nèi)分泌功能改變均與內(nèi)皮細胞功能障礙有著密切的聯(lián)系和可能的因果關(guān)系[22-24],因此,建立一種簡便高效的大鼠CCECs 原代培養(yǎng)方法,對我們進一步開展氧化應(yīng)激、慢性炎癥等因素產(chǎn)生的相關(guān)特異性分子和內(nèi)皮細胞功能障礙之間的關(guān)系研究成為必須,同時對研究內(nèi)皮相關(guān)性功能改變與ED 關(guān)系及其藥物篩選方面具有重要應(yīng)用價值。
目前關(guān)于CCECs 原代培養(yǎng)研究主要來源于人、兔和小鼠[25-27],但人源組織來源稀缺,兔的繁殖速度較慢,而小鼠的陰莖組織較小,現(xiàn)有的方法無法較好滿足實驗需求。 SD 大鼠因其價格低廉、繁殖力強等優(yōu)點常作為糖尿病ED、高血壓ED 和高血脂ED 等ED 模型的首選實驗動物[28-30],因此本實驗最終選擇了SD 大鼠,分離其陰莖海綿體組織,并探討了建立大鼠CCECs 的方法。
CCECs 的培養(yǎng)方法主要包括組織塊培養(yǎng)法[25]和酶消化法[31]兩種。 組織塊培養(yǎng)法雖然操作簡單、成本較低,但培養(yǎng)細胞所需的組織較多、培養(yǎng)周期長,容易污染,而以往的酶消化法采用多種消化酶,花費高昂、操作繁瑣,因此,在本研究中我們采用了Ⅱ型膠原酶消化法聯(lián)合機械刮除法獲取原代SD 大鼠CCECs,并開展了研究。 本實驗將0.5% Ⅱ型膠原酶消化時間控制在2 ~2.5h,每隔15min 震蕩2 ~3min,可減少整體酶消化時間,最大程度保證細胞活性。 在原代培養(yǎng)48h 后進行首次換液,以保證活細胞已全部貼壁,因細胞群在原代培養(yǎng)3 ~4 天時開始生長,故于原代培養(yǎng)96h 時,采用細胞刮除法清除獨立成片和散在分布的平滑肌細胞與成纖維細胞群,可有效避免成纖維細胞等雜細胞污染,獲得較高純度的內(nèi)皮細胞。 既往原代培養(yǎng)內(nèi)皮細胞的文獻報道中,多采用M199 培養(yǎng)基、DMEM 高糖培養(yǎng)基、DMEM/F12 培養(yǎng)基等為基礎(chǔ)培養(yǎng)液[32-34],添加血清及生長因子進行原代培養(yǎng),綜合考慮高糖培養(yǎng)液可能對實驗結(jié)果的干擾,以及保證細胞最佳活力等因素,我們采用添加多種生長刺激因子的EGM-2 內(nèi)皮細胞專用完全培養(yǎng)液,以促進內(nèi)皮的生長,保證原代內(nèi)皮細胞培養(yǎng)的成功率。 原代培養(yǎng)至第4 ~5 天時,細胞匯合成單層,呈鋪路石樣鑲嵌式排列,傳代培養(yǎng)至第2 代時繪制生長曲線,細胞在第2 ~4 天進入對數(shù)生長期,培養(yǎng)至第6天時進入平臺期,呈“S”形生長。 第2 ~3 代細胞形態(tài)均可保持內(nèi)皮細胞典型的“鋪路石樣”特征,HE 染色可見細胞呈扁平的多角形,細胞核卵圓居中,核仁可見,且vWF 在CCECs 的胞質(zhì)內(nèi)的表達和細胞生長周期較為穩(wěn)定,原代細胞傳至4 代后,細胞出現(xiàn)胞質(zhì)增大衰老、雜細胞增多等現(xiàn)象,且細胞逐漸阻滯于G0/G1 期,因此我們選擇第2 ~3 代CCECs 進行后續(xù)研究。 CD31又稱為血小板內(nèi)皮細胞黏附分子在血管內(nèi)皮細胞內(nèi)以及胞間連接處廣泛表達[35]。 vWF 即血管性血友病因子,作為一種多聚糖蛋白,在內(nèi)皮細胞中合成[36],CD31和vWF 共同陽性表達是目前對原代提取內(nèi)皮細胞鑒定其表型的通用做法,并得到學(xué)者們的廣泛認同[5,37]。 本研究結(jié)果顯示CD31 和vWF 免疫熒光共染鑒定內(nèi)皮細胞均為陽性表達率達(86.19±1.14)%,表明酶消化法結(jié)合機械刮除法可成功獲得CCECs,并且第2 ~3 代CCECs 可作為后續(xù)實驗研究的最佳選擇時期。
綜上,本研究采用酶消化法結(jié)合機械刮除法,運用EGM-2 培養(yǎng)液可成功分離培養(yǎng)大鼠CCECs,鏡下呈典型的“鋪路石”狀生長,vWF 和CD31 免疫熒光陽性反應(yīng),純度達到(86.19±1.14)%,并可穩(wěn)定維持3 ~4 代,為基于CCECs 基礎(chǔ)上開展相關(guān)研究提供可靠簡便的細胞模型建立方法,如能建立起穩(wěn)定傳代的CCECs 永生細胞系,可為男科領(lǐng)域開展基于CCECs 層面的基礎(chǔ)研究與藥物篩選帶來極大的便利和更多的選擇。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
