梅花鹿成纖維細胞因子受體2基因多態性及其與茸重性狀的關聯分析
2022-08-23 02:39:46張禾垟劉琳玲李浩東鄭軍軍王桂武
中國畜牧獸醫 2022年8期
周 雅,張禾垟,劉琳玲,李浩東,鄭軍軍,王桂武
(中國農業科學院特產研究所,長春 130112)
鹿茸具有抗疲勞、增強免疫力和改善性功能等多種藥理作用,是重要的動物藥材之一[1]。梅花鹿是中國茸鹿養殖的主體,其產茸量是直接關系到養殖業經濟收益的重要經濟性狀,因此該性狀在育種中備受關注。研究表明,鹿茸的再生是包括軟骨、皮膚、血管和神經等組織的完全再生,因而茸重性狀受多基因控制,依靠傳統的育種方法,存在周期長、效率低等問題[2]。隨著分子生物學技術的發展,梅花鹿育種進入分子育種是必然的趨勢,相關分子標記的研究對推動其分子育種進程、提高動物產品產量等具有重要意義。
成纖維細胞因子受體2(fibroblast growth factor receptor 2,FGFR2)是成纖維細胞生長因子受體家族(fibroblast growth factor receptor,FGFR)的一員,由3個胞外免疫球蛋白結構域、1個疏水跨膜結構域和1個胞內酪氨酸激酶結構域組成[3]。其中第二和第三免疫球蛋白結構域是配體、成纖維細胞生長因子(fibroblast growth factors,FGFs)和肝素的主要結合位點[4-5]。而第3個免疫球蛋白結構域的mRNA存在2個可變剪切體,形成FGFR2b和FGFR2c 2個亞型[6]。在上皮細胞中編碼FGFR2b亞型,主要與FGF7、FGF10結合,而在間質細胞中編碼FGFR2c亞型,主要與FGF2、FGF4、FGF6、FGF8、FGF9等親和力較高[6-7]。FGFR2與相關的FGF結合可激活多個信號通路調節血管內皮細胞、上皮細胞、成骨細胞和成纖維細胞等的增殖和分化,在皮膚和骨等多個組織發育中起重要作用,其中1個配體FGF2已被證明與梅花鹿鹿茸快速生長密切相關[8-9]。此外,FGFR2還可能通過與FGF21的受配體作用,產生相關信號分子,誘導毛囊由生長期和退化期進入靜止期,而FGFR2b缺失會導致毛囊形成遲鈍并減少[10-11]。FGFR2的多種功能與鹿茸生長過程相關,故推測該基因可能是促進鹿茸生長的關鍵基因。Hu等[12]通過梅花鹿產茸量相關的全基因組關聯分析(genome-wide association study,GWAS)篩選到該基因,推測為茸重性狀候選基因,但尚未進一步證明。
本研究以梅花鹿為研究對象,通過PCR直接測序檢測FGFR2基因外顯子上的突變,并使用飛行質譜法分析基因型,與茸重性狀進行關聯分析,旨在篩選梅花鹿茸重性狀相關的分子標記,為梅花鹿分子育種提供依據。
1 材料與方法
1.1 試驗動物
在吉林省長春市某鹿場選取314頭24月齡飼養管理條件一致的梅花鹿作為研究對象。對314頭梅花鹿進行頸靜脈采血,經EDTA抗凝處理,準確記錄編號后暫存至冰盒內帶回實驗室進行分裝,置于-20 ℃保存備用。使用電子秤稱量生長45 d左右的二杠茸重量,精確到1 g。所有樣本茸重經平均值±3倍標準差(μ±3σ)剔除異常值,近似符合正態分布。
1.2 主要試劑
血液DNA提取試劑盒和2×TransStart?FastPfu PCR SuperMix (-dye)均購自北京全式金生物技術有限公司;2×EsTaqMasterMix(Dye)購自北京康為世紀生物科技有限公司;DNA Marker、6×Loading Buffer和10×Loading Buffer均購自TaKaRa公司。
1.3 DNA提取與PCR擴增
按照血液DNA提取試劑盒操作步驟提取血液樣品DNA。用NanoDrop 2000檢測所提取DNA 的純度和濃度,并用1.2%瓊脂糖凝膠電泳檢測DNA完整性,將質量合格的DNA樣品保存備用。根據梅花鹿基因組中FGFR2基因序列[13],使用NCBI中的Primer-BLAST設計FGFR2基因第1~17外顯子擴增引物,擴增產物包含全部外顯子和部分內含子序列,引物序列見表1。引物均由生工生物工程(上海)股份有限公司合成。PCR反應體系25 μL:2×EsTaqMasterMix 12.5 μL,上、下游引物各0.5 μL,DNA模板1 μL,ddH2O補充至25 μL。PCR擴增程序:94 ℃預變性5 min;94 ℃變性30 s,退火(各引物退火溫度見表1)30 s,72 ℃延伸30 s,共35個循環;72 ℃延伸7 min;4 ℃保存。PCR產物用2.0%瓊脂糖凝膠電泳檢測合格后送生工生物工程(上海)股份有限公司測序。測序結果利用DNAMAN和Chromas 2軟件進行比對,篩選SNPs位點。

表1 梅花鹿FGFR2基因各外顯子的引物信息
1.4 基因分型
FGFR2基因候選位點利用MassARRAY?技術進行分型,該步驟交由北京閱微基因技術股份有限公司完成,相關分型引物信息如表2所示,具體操作步驟如下:①將質檢合格的DNA樣本放入384孔板進行PCR擴增,PCR反應體系5 μL/孔:PCR Buffer(含15 mmol/L MgCl2) 0.625 μL,MgCl2(25 mmol/L) 0.325 μL,dNTP Mix(4種dNTP 各自濃度為25 mmol/L) 0.1 μL,擴增引物Mix(上、下游引物各自濃度為500 nmol/L) 1 μL,HotStarTaq(5 U/μL) 0.1 μL,DNA模板(10 ng/μL) 1 μL,ddH2O 1.850 μL。PCR擴增程序:94 ℃預變性5 min;94 ℃變性20 s,56 ℃退火30 s,72 ℃延伸1 min,共45個循環;72 ℃延伸3 min;4 ℃保存。②PCR產物進行SAP純化反應,反應體系2 μL/孔:SAP Buffer(10×) 0.17 μL,SAP Enzyme(1 U/μL) 0.3 μL,ddH2O 1.53 μL。PCR反應程序:37 ℃溫育40 min,85 ℃熱失活5 min,4 ℃保存。③單堿基延伸反應,反應體系2 μL/孔:iPLEX buffer plus 0.2 μL,iPLEX terminator 0.2 μL,擴增引物Mix 0.940 μL,iPlex Enzyme 0.041 μL,ddH2O 0.619 μL。反應程序:94 ℃預變性30 s;94 ℃變性5 s,52 ℃退火5 s,80 ℃延伸5 s,共40個循環;72 ℃延伸180 s,4 ℃保存。④樹脂純化:將反應產物(共9 μL)稀釋3倍,使用樹脂進行脫鹽,脫鹽處理后的樣品點在樣品靶上,自然結晶。⑤上機進行質譜檢測,并收集數據。
1.5 數據統計分析
利用Excel 2019計算梅花鹿FGFR2基因的基因型頻率、基因頻率及相關遺傳多樣性參數:觀測雜合度(heterozygosity observed,Ho)、期望雜合度(expected heterozygosity,He)、有效等位基因數(effective number of alleles,Ne)和多態信息含量(polymorphism information content,PIC),并進行χ2適合性檢驗,檢測是否符合Hardy-Weinberg平衡定律。利用Haploview 4.2軟件進行單倍型分析。利用SPSS 25.0軟件中一般線性模型(general linear model,GLM)單因素方差分析對梅花鹿群體中各位點基因型以及單倍型與茸重性狀進行關聯分析,存在群體數量低于5%的基因型的位點采用非參數檢驗Kruskal-Wallis H檢驗方法進行分析,使用最小顯著差異法(least significant difference,LSD)進行多重比較,結果用平均值±標準誤表示,以P<0.05為差異顯著性判斷標準。模型:Yij=μ+Gi+eij。其中,Yij為茸重性狀觀察值,μ為群體均值,Gi為基因型固定效應,eij為隨機殘差效應。

表2 SNPs引物信息
2 結 果
2.1 PCR擴增
PCR擴增結果顯示,17對引物的PCR擴增產物均呈現清晰、明亮的特異性條帶(圖1),且與預期產物大小一致,可用于后續試驗。

M,DL2000 DNA Marker;1~17分別為FGFR2基因第1~17外顯子擴增產物M,DL2000 DNA Marker;1-17,Amplification products of exons 1 to 17 of FGFR2 gene圖1 梅花鹿FGFR2基因PCR擴增結果Fig.1 PCR amplification results of FGFR2 gene in sika deer
2.2 測序驗證
經檢測發現,梅花鹿FGFR2基因存在12個SNPs,其中g.80998742 G>A、g.80987708 G>A、g.80975864 T>G、g.80936928 C>T和g.80928230 C>T位點分別位于外顯子2、外顯子4、外顯子5、外顯子12和外顯子15上;g.80943673 T>C和g.80943683 C>A位點均位于內含子8;g.80940697 T>G、g.80938545 G>A、g.80938352 C>T、g.80928299 C>T和g.80928115 A>G分別位于內含子9、內含子10、內含子11、內含子14和內含子15(部分結果見圖2,外顯子2為反向測序其余均為正向測序)。80998742 G>A、g.80987708 G>A、g.80975864 T>G、g.80936928 C>T和g.80928230 C>T 5個外顯子上的突變均未引起氨基酸改變,為同義突變。

圖2 梅花鹿FGFR2基因部分SNPs位點測序結果Fig.2 Sequencing results of partial SNPs in FGFR2 gene in sika deer
2.3 質譜分型
分型結果顯示,g.80975864 T>G位點未分型成功,其余11個位點可做后續分析。其中g.80940697 T>G和g.80936928 C>T僅存在2種基因型,其余位點均存在3種基因型(部分結果見圖3)。

圖3 梅花鹿FGFR2基因部分SNPs位點分型結果Fig.3 Typing results of partial SNPs in FGFR2 gene in sika deer
2.4 梅花鹿FGFR2基因多態性分析
梅花鹿FGFR2基因各位點基因型頻率、基因頻率、相關遺傳多樣性參數及χ2檢驗結果見表3。由表3可知,各位點Ho和He基本一致,g.80943673 T>C、g.80943683 C>A及g.80938352 C>T位點屬于中度多態(0.25

表3 梅花鹿FGFR2基因SNPs位點的群體遺傳學分析
2.5 FGFR2基因多態性與梅花鹿茸重性狀的關聯分析
FGFR2基因11個SNPs位點的不同基因型與梅花鹿茸重性狀的關聯分析結果見表4。結果顯示11個位點各基因型之間茸重差異均不顯著(P>0.05)。
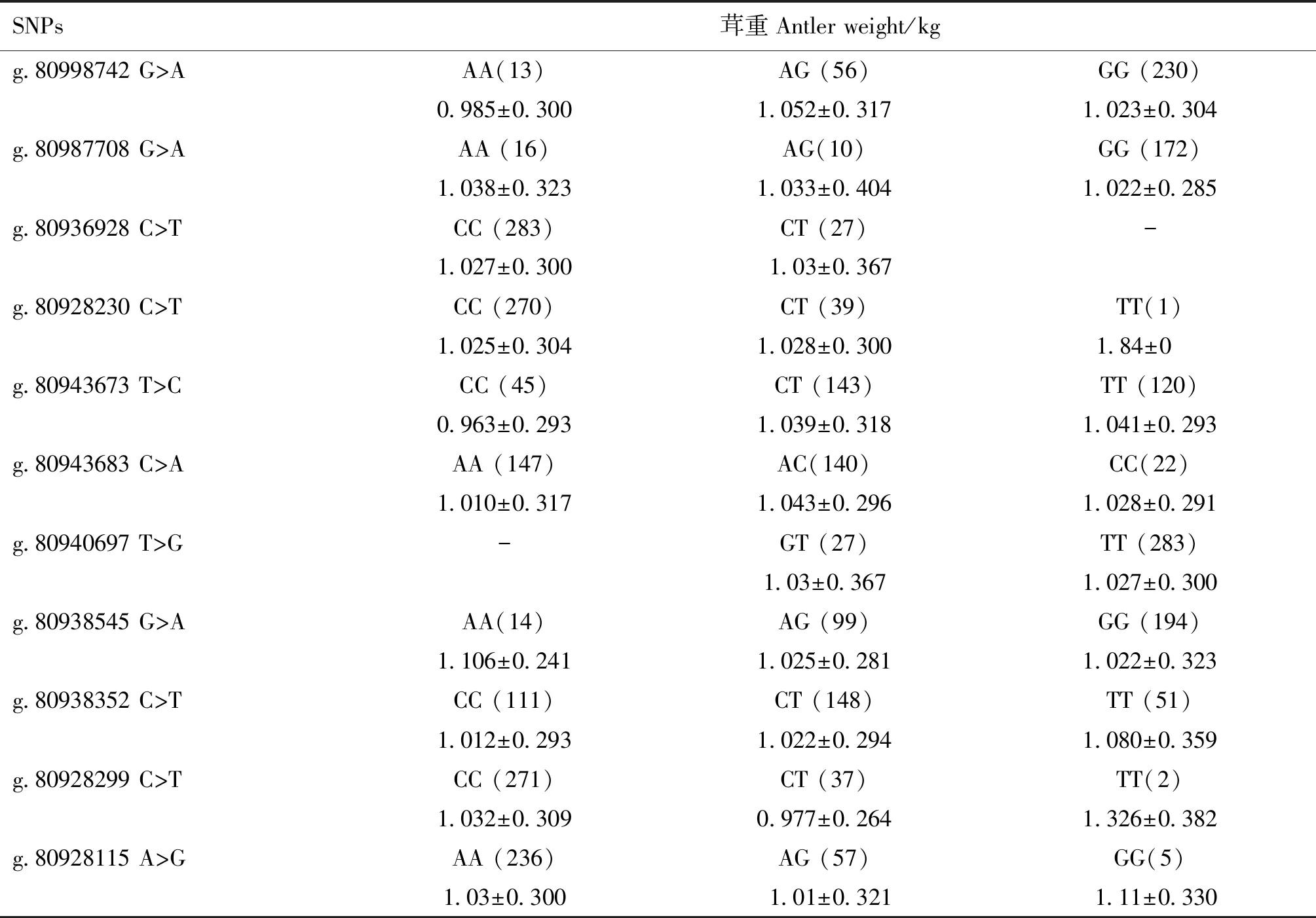
表4 梅花鹿FGFR2基因SNPs位點各基因型與茸重的關聯分析
2.6 FGFR2基因單倍型與梅花鹿茸重性狀的關聯分析
對梅花鹿群體FGFR2基因11個SNPs位點進行連鎖不平衡和單倍型分析,結果見圖4。FGFR2基因g.80987708 G>A位點檢出率較低,在分析中剔除,g.80938545 G>A、g.80940697 T>G、g.80943673 T>C和g.80943683 C>A位點之間處于強連鎖不平衡狀態(D′>0.96,r2>0.72)。
通過連鎖不平衡分析發現,梅花鹿群體中存在5種單倍型,分別為GTCA、GTTA、ATTC、GTTC和GGTC。由表5可知。FGFR2基因各單倍型間茸重差異均不顯著(P>0.05)。

圖4 梅花鹿FGFR2基因SNPs連鎖不平衡分析Fig.4 Linkage disequilibrium analysis of SNPs of FGFR2 gene in sika deer

表5 FGFR2基因單倍型對梅花鹿茸重的影響
3 討 論
鹿茸具有很高的藥用價值,價格昂貴,故茸重性狀也廣受關注。近年來,隨著分子生物技術的發展,分子育種技術已廣泛應用于畜禽育種中,并取得了良好的效果。鹿茸屬于限性性狀,通過傳統的育種方法具有很大的局限性,利用分子遺傳標記對梅花鹿鹿茸生長特性進行研究,可以有效地克服傳統育種方法的不足,提高梅花鹿鹿茸產量。鹿茸生長機制復雜,其相關分子標記也相當匱乏。胡鵬飛等[14]選用飼養條件基本一致的高、低產梅花鹿各50只做重測序的全基因組關聯分析,篩選出與茸重性狀顯著相關的SNPs位點96個。Jia等[15]通過轉錄組測序挖掘梅花鹿茸重性狀相關的表達序列標簽(expressed sequence tag,EST)微衛星,篩選出了8個EST微衛星,特別是M009和M027,可作為二杠茸茸重性狀的分子標記。二者分別從基因組測序和轉錄組測序挖掘與茸重性狀相關的分子標記,豐富了篩選高產梅花鹿的遺傳資源。杜志恒等[16]對生長激素(growth hormone 1,GH1)基因進行多態性檢測,結果表明,其內含子2上存在G→A突變,存在3種基因型,各基因型在第五鋸產茸量存在一定差異。Yang等[17]發現,褪黑激素Ⅰ型受體a亞型(melatonin receptor 1A,MTNR1A)基因外顯子2上存在G629C突變,存在3種基因型,CC基因型產茸估測值顯著高于GC基因型,GC基因型產茸估測值顯著高于GG基因型。
FGFR2基因位于梅花鹿第9號染色體,共17個外顯子,編碼區長2 460 bp,編碼819個氨基酸。既往研究發現FGFR2在骨發育中發揮重要作用,可促進間充質干細胞的募集和增殖,通過正向調節Wnt/β-catenin信號通路促進成骨細胞的分化,在骨損傷后修復中研究廣泛[18]。FGFR2還可調節癌癥干細胞的增殖和凋亡,在乳腺癌[19-20]、胃癌[21]、食管鱗癌[22]和膽管癌[23]等多種癌癥的發展中發揮重要作用。本研究采用直接測序法檢測FGFR2基因17個外顯子及部分內含子的多態性,共發現12個SNPs位點,進一步分析發現,g.80943673 T>C、g.80943683 C>A及g.80938352 C>T位點雜合度較高,多態信息含量為0.3597、0.3307及0.3655,為中度多態位點,這3個位點的群體遺傳變異程度較大,遺傳多樣性比較豐富,具有一定的選擇潛力,而其余8個位點雜合度較低,為低度多態位點,選擇潛力較小。χ2檢驗結果顯示,g.80998742 G>A和g.80987708 G>A位點偏離Hardy-Weinberg平衡狀態,其他位點均處于Hardy-Weinberg平衡狀態。連鎖不平衡和單倍型分析結果顯示,FGFR2基因4個位點處于強連鎖不平衡狀態,產生5種單倍型。關聯分析結果顯示,11個位點各基因型及5種單倍型之間茸重差異均不顯著。
4 結 論
本研究梅花鹿群體中FGFR2基因存在12個SNPs,其中5個為同義突變,有1個同義突變位點(g.80975864 T>G)未分型成功。該基因有4個SNPs處于強連鎖不平衡狀態,產生5種單倍型。FGFR2基因各位點基因型及單倍型間茸重差異均不顯著,說明這11個SNPs均不適用于梅花鹿茸重性狀的選育。
