天目山毛竹林皆伐后群落的恢復特征
2022-08-18 10:49:12劉宗悅徐鈞杰祁祥斌余樹全
浙江農林大學學報 2022年4期
陳 旭,劉宗悅,徐鈞杰,祁祥斌,余樹全
(1.浙江農林大學 林業與生物技術學院,浙江 杭州 311300;2.浙江天目山國家級自然保護區管理局,浙江 杭州 311311)
毛竹Phyllostachys edulis是竹亞科Bambusoideae剛竹屬Phyllostachys大型散生竹,具有繁殖容易、生長快、產量高、材質好、用途廣等特點[1]。20世紀90年代到2010年前,毛竹因其竹材、竹芛具有較高的經濟效益而被廣泛種植,其中浙江省毛竹林面積達100多萬hm2。但近年來,隨著毛竹種植面積增加以及勞動力成本的提高,竹材的市場價格大幅下降,許多地方毛竹林已無人管理,導致毛竹擴張速度加快[2-3]。研究顯示,1956—2014年,天目山毛竹林面積從55.10 hm2自然蔓延擴張到105.38 hm2,漲幅近1倍[4],對天目山國家級自然保護區原生植被的蠶食現象日益嚴重。毛竹因其無性繁殖能力強,可以靠地下竹鞭滲透使其具有很強的水平擴展能力,當地下竹鞭滲透到相鄰森林中,依靠其快速的營養生長而迅速成林,改變生境條件和群落結構,從而對森林的物種多樣性和生態系統穩定性產生嚴重威脅。白尚斌等[5]的研究表明:毛竹入侵相鄰的森林群落時會導致喬木和灌木層的物種豐富度、Simpson指數和Pielou均勻度指數顯著降低。目前,在許多地方毛竹林的無序自然蔓延擴張已造成嚴重生態退化,威脅到生物多樣性保護和區域生態安全,因此,控制毛竹林無序蔓延擴張,開展生態修復,受到全社會廣泛關注。
森林采伐是森林生態系統管理重要手段之一,皆伐作為一種森林經營管理措施,會對林地上物種組成和生境條件產生強烈的影響。森林皆伐后,喬木層已不存在,加上采伐過程中的影響,森林群落實際退化為以灌草為主的早期演替階段,地表被大量采伐剩余物所覆蓋,植物生長所需的光照、溫度、水分等都發生了明顯變化,環境異質性增加,為更多類型的植物萌發和生長提供有利條件。如河南寶天曼地區栓皮櫟Quercus variabilis林皆伐后群落喬木層物種豐富度和多樣性指數明顯增加[6],福建永安市馬尾松Pinus massoniana人工林皆伐跡地封育1 a后,新增天然闊葉樹種多達71種、密度高達64 547株·hm-2[7]。皆伐后群落自然恢復過程中樹種的功能特征組成會發生改變,比如反映群落外貌特征的常綠樹種相對落葉樹種的比例會增加[8];反映光耐受性的陽性樹種主導地位逐漸減弱,而中性及耐蔭樹種的優勢度明顯增加[9],因此研究樹種功能特征組成的變化可以反映群落恢復的效果。本研究通過對天目山毛竹純林進行皆伐處理,設置固定樣地進行定株定位觀測,研究分析毛竹林皆伐后自然恢復過程中群落的物種組成、樹種功能特征和物種多樣性的動態變化,旨在為天目山國家級自然保護區生物種多樣性保護與恢復提供指導,為亞熱帶地區毛竹林無序蔓延擴張區域的生態退化修復和生物多樣性保護提供借鑒。
1 研究地區與研究方法
1.1 研究區概況
天目山國家級自然保護區位于浙皖兩省交界處浙江省杭州市臨安區境內(30°18′30″~30°24′55″N,119°24′1l″~119°28′21″E),面積為 4 300 hm2,海拔為 300~1 556 m。天目山屬于中亞熱帶向北亞熱帶過渡地帶,受海洋暖濕氣流影響,四季分明,年平均氣溫為8.8~14.8 ℃,年平均降雨日為159.2~183.1 d,年降水量為1 390~1 870 mm,分布著典型的中亞熱帶森林生態系統[10]。保護區內植被的分布有著明顯的垂直帶譜特征:在海拔230~850 m分布常綠闊葉林,850~1 100 m分布常綠落葉闊葉混交林,1 100~1 350 m分布落葉闊葉林,1 400 m以上為落葉矮林[11]。毛竹林作為一種特殊的森林類型,主要分布在海拔350~900 m。由于不允許開展森林經營活動,保護區內毛竹林完全處于自然狀態下,立竹度高,林下植被生物多樣性較低,零星分布有豹皮樟Litsea coreanavar.sinensis、毛花連蕊茶Camelia fraterna、微毛柃Eurya hebeclados、細葉青岡Cyclobalanopsis gracilis和牛鼻栓Fortunearia sinensis等。
1.2 樣地設置與調查
1.2.1 樣地設置 2015 年對保護區內 54 hm2毛竹純林進行皆伐作業,皆伐時伐除全部毛竹,對毛竹純林中零星分布杉木Cunninghamia lanceolata、金錢松Pseudolarix amabilis、楓香Liquidambar formosana等高大喬木樹種全部保留,林下植物盡量保留。在皆伐后2 a內持續清除萌發的新竹,直至不再有新竹萌發。毛竹皆伐2 a后植物群落進行自然更新恢復。
在皆伐后自然更新群落區塊設置11個樣地(20 m×20 m),其中3號樣地有楓香2株、金錢松2株,5號樣地有楓香2株、杉木5株,8號樣地有4株杉木,其余樣地無喬木分布。受采伐工作影響,林下植物僅有零星少量分布。在選擇樣地時避免人為干擾較多、動物活動頻繁的地段,同時覆蓋各個海拔梯度,使得選取的樣方能夠代表毛竹林皆伐作業的整體情況,并用全球定位系統(GPS)和羅盤儀測量每個樣地的經度、緯度、海拔、坡度、坡向等環境因子(表1)。坡向采用4方向法表示,樣地4個角設置長期固定標志。
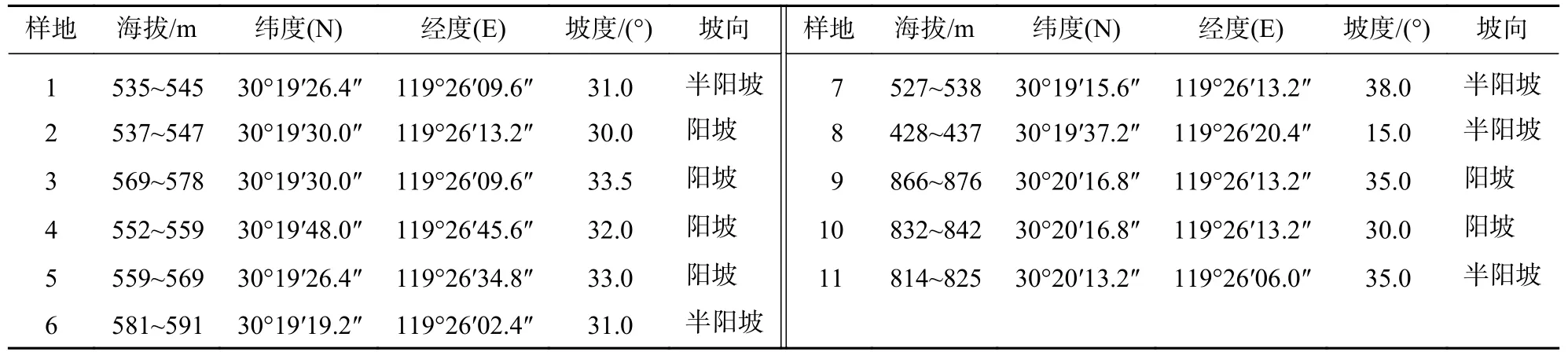
表1 樣地基本情況Table 1 Basic conditions of sample plot
1.2.2 樣地調查 在樣地 4 個角與中間設置 5 個 (5 m×5 m)小樣方,共獲得 55 個小樣方,并于 2018—2020年10—11月對每個小樣方的喬木、灌木和草本進行調查。調查內容包括喬木層:對胸徑(DBH)≥1 cm的木本植物掛牌并記錄其種名、胸徑、樹高、冠幅等;灌木層:調查種名、基徑、樹高、蓋度等;草本層:調查種名、株數、高度、蓋度等。
1.3 數據處理
1.3.1 重要值 重要值(VI)是森林群落研究中被常用來衡量某個樹種優勢程度大小的重要指標。①喬木層的重要值(VI喬)=[(相對多度+相對顯著度+相對高度)/3]×100%。②灌草層的重要值(VI灌草)=[(相對多度+相對蓋度)/2]×100%。其中:相對多度是某物種株數占所有物種總株數的比值;相對顯著度是某物種的胸高斷面積占所有物種的胸高斷面積的比值;相對高度指的是某物種的高度占所有物種總高度的比值;相對蓋度是指某物種的蓋度占所有物種總蓋度的比值[12]。
1.3.3 補員率與死亡率 ①死亡率M=(lnN0-lnSt)/t。②補員率R=(lnNt-lnSt)/t。其中:N0和Nt分別為第1次和第2次調查時的株數,St是第2次調查時的存活株數,t為2次調查時間的間隔,本研究為1 a。種群大小變化率采用補員率和死亡率的差值來表示,即種群大小變化率=補員率-死亡率[15]。
1.3.4 樹種功能特征組成 本研究依據《天目山植物志》和《浙江省植物志》對群落的樹種功能特征進行劃分[16-17]。根據葉生活型將樹種劃分為常綠和落葉2種類型,常綠與落葉樹種占比可以反映群落外貌特征變化;根據生長特性將樹種分為喬木、小喬木、灌木3種生長型;根據樹種對光的耐受性及生長環境,將樹種劃分為陽性、中性、陰性3種類型。
1.3.5 數據處理 采用 Excel 2019、SPSS 24.0、Origin 等軟件進行數據處理、分析與作圖。
2 結果與分析
2.1 物種組成動態
2.1.1 喬木層物種組成動態 由表2可知:2018—2020年,群落恢復過程中喬木層植物共有130種,隸屬于45科86屬。每年科、屬的數量變化不大,但物種數增加了4種,其中2018—2019年新增8種,退出7種,2019—2020年新增6種,退出3種,主要是一些偶見種,如云山八角楓Alangium kurzii、海州常山Clerodendrum trichotomum從群落中退出,化香Platycarya strobilacea、杭州榆Ulmus changii等在群落中新增。喬木層主要科有大戟科Euphorbiaceae、蕓香科Rutaceae等,以白背葉Mallotus apelta、朵花椒Zanthoxylum molle等為主要優勢種。
2.1.2 灌木層物種組成動態 從表2 可以看出:2018—2020 灌木層植物共有 224 種,隸屬于 65 科137屬。隨著時間推移,灌木層科、屬、種數量均顯著下降,其年降幅分別為17.2%、27.3%、27.0%。這是因為隨著密度增大、林分郁閉,群落光照條件變弱和養分競爭加劇,導致植株大量死亡減少,其中2018—2019年新增4種,退出75種,2019—2020年新增6種,退出41種。灌木層主要科有薔薇科Rosaceae、山茶科Theaceae等,并以油茶Camellia oleifera、毛花連蕊茶等為主要優勢種。
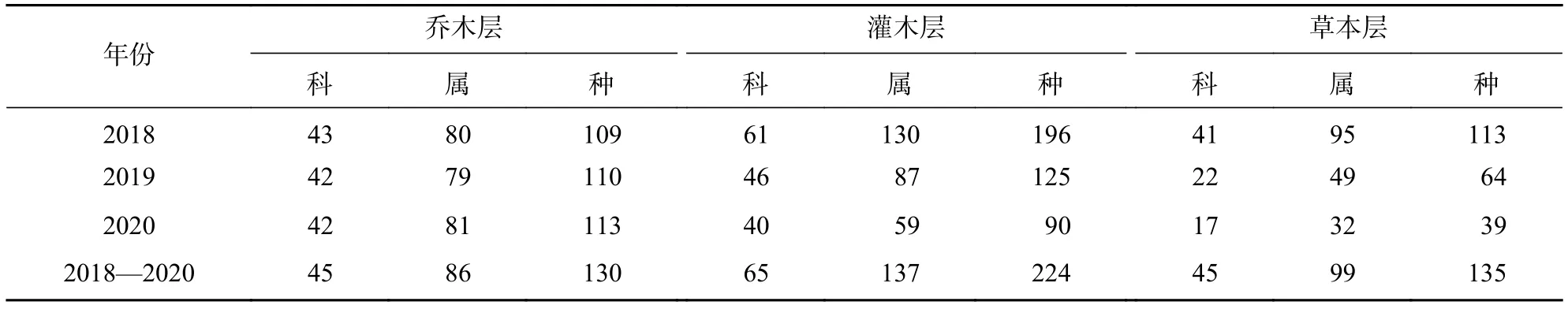
表2 2018—2020 年群落科屬種組成情況Table 2 Composition of families, generas and species of community from 2018 to 2020
2.1.3 草本層物種組成動態 從表2可以看出:2018—2020草本層植物有135種,隸屬于45科99屬,隨著時間推移,草木層科、屬、種的數量也均顯著下降,其年下降幅度分別為29.3%、33.2%、32.7%,其中2018—2019年新增19種,退出66種,2019—2020年新增8種,退出33種。草本層主要科有莎草科Cyperaceae、禾本科Gramineae等,并以栗褐薹草Carex brunnea、三穗薹草Carex tristachya、顯子草Phaenosperma globosa等為主要優勢種。
2.2 優勢種的動態變化
2.2.1 優勢種組成變化 由表3可知:喬木層排名前20位的優勢種重要值之和高達80,能代表喬木層主要樹種的變化。優勢度最明顯的種是朵花椒,其重要值大于20,白背葉、臭椿Ailanthus altissima、臭辣樹Euodia fargesii等樹種的優勢度次之,它們的重要值均高于5。不同優勢種重要值的變化在演替過程中有很大差異,如掌葉覆盆子Rubus chingii、朵花椒的重要值3 a內分別下降了4.00、5.90,而白背葉、小構樹Broussonetia kazinoki的重要值卻分別增加了3.16、1.43。排名前20位的優勢種僅有油茶、小葉青岡Cyclobalanopsis myrsinifolia、苦櫧Castanopsis sclerophylla等為常綠樹種,并且重要值均偏低。
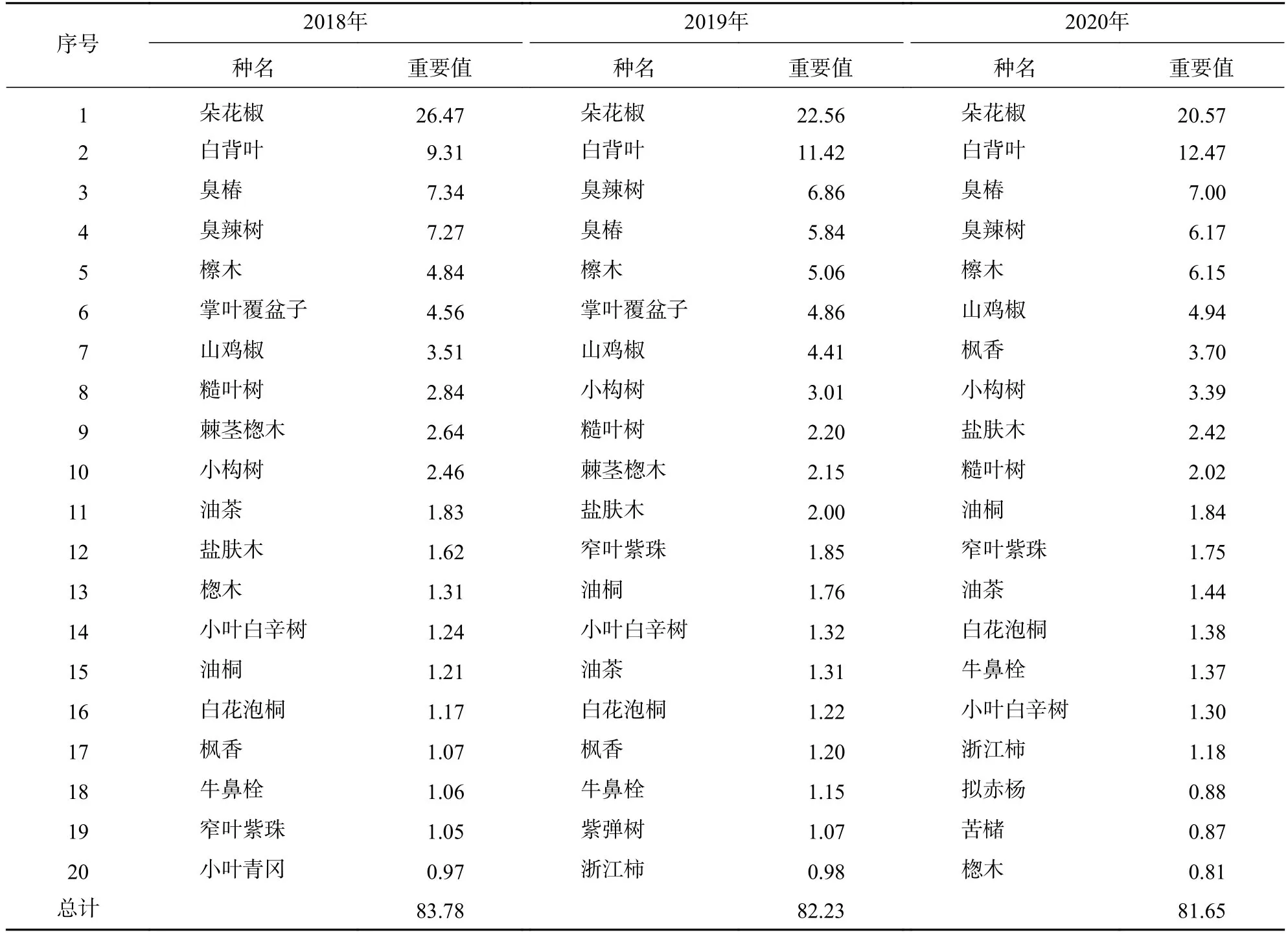
表3 2018—2020 年喬木層重要值排名前 20 的物種Table 3 Top 20 species ranked by importance values in tree layers from 2018 to 2020
灌木層排名前20位的優勢種的重要值之和為60.93~70.74。由表4可以看出:大部分優勢種重要值約為2,表明優勢種的優勢程度不明顯,但優勢種種類變化明顯,比如朵花椒、掌葉覆盆子、臭椿等樹種被微毛柃、紫麻Oreocnide frutescens、油茶等樹種取代。灌木層優勢度較高的種有茶Carex crebra、油茶、微毛柃等常綠樹種。這些優勢種在2019—2020年的優勢度明顯增加,尤其茶的重要值增加了9.25,增加了3倍左右。另外,豹皮樟、細葉青岡等常綠樹種也出現在灌木層,并且重要值呈現逐年小幅增加趨勢。
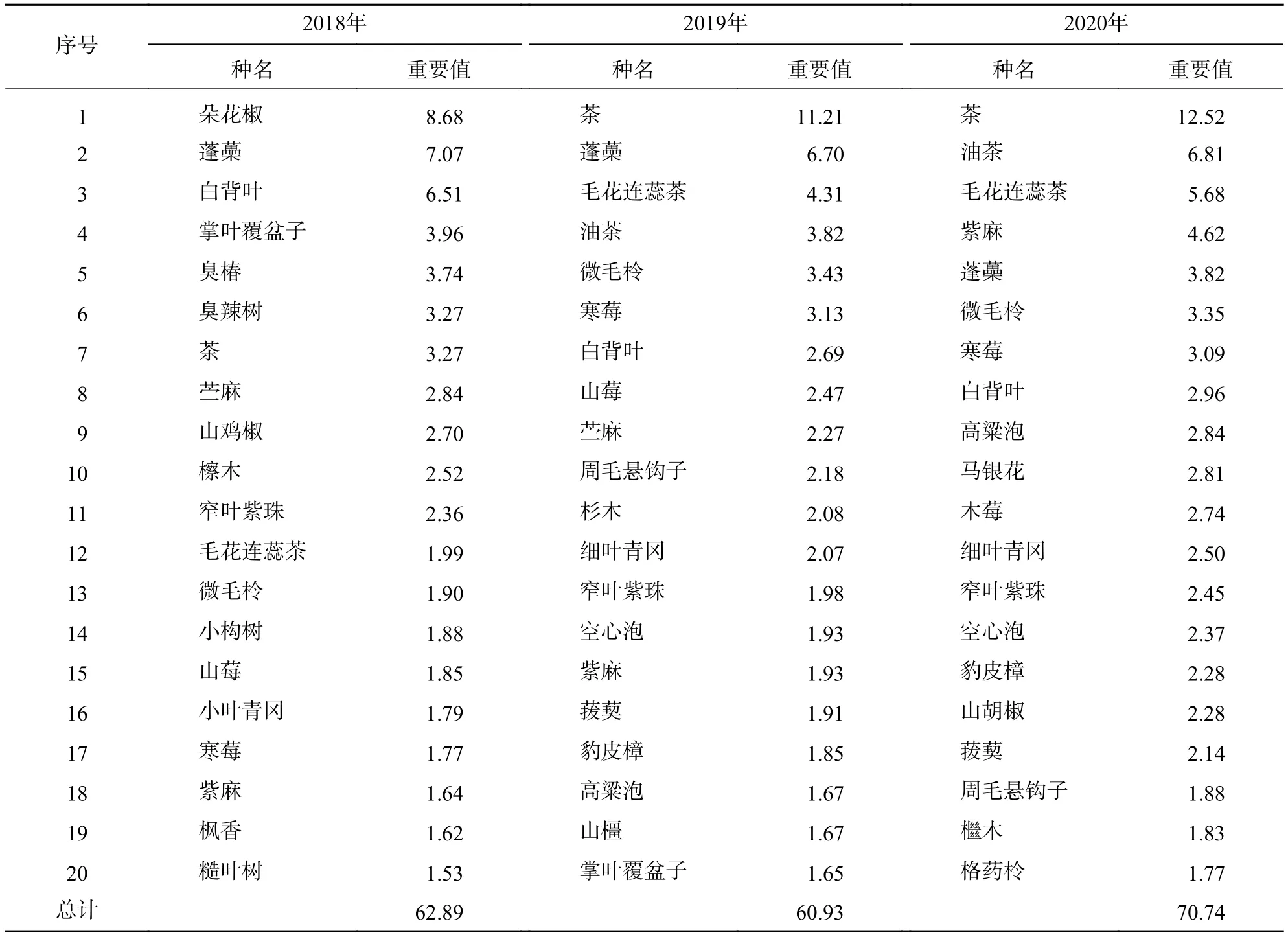
表4 2018—2020 年灌木層重要值排名前 20 的物種Table 4 Top 20 species ranked by importance values in shrub layer from 2018 to 2020
由表5可知:草本層排名前20的優勢種重要值之和為78.06~96.55,并且呈現逐年大幅上升的趨勢。草本層主要優勢種的種類沒有改變,但重要值的增長卻很明顯,如栗褐薹草、顯子草、三穗薹草的重要值3 a內分別增加了17.92、7.80、2.81,在2020年這3種植物的重要值之和高達59.73,這與莎草科、禾本科耐蔭、耐潮濕的生長特性有關。
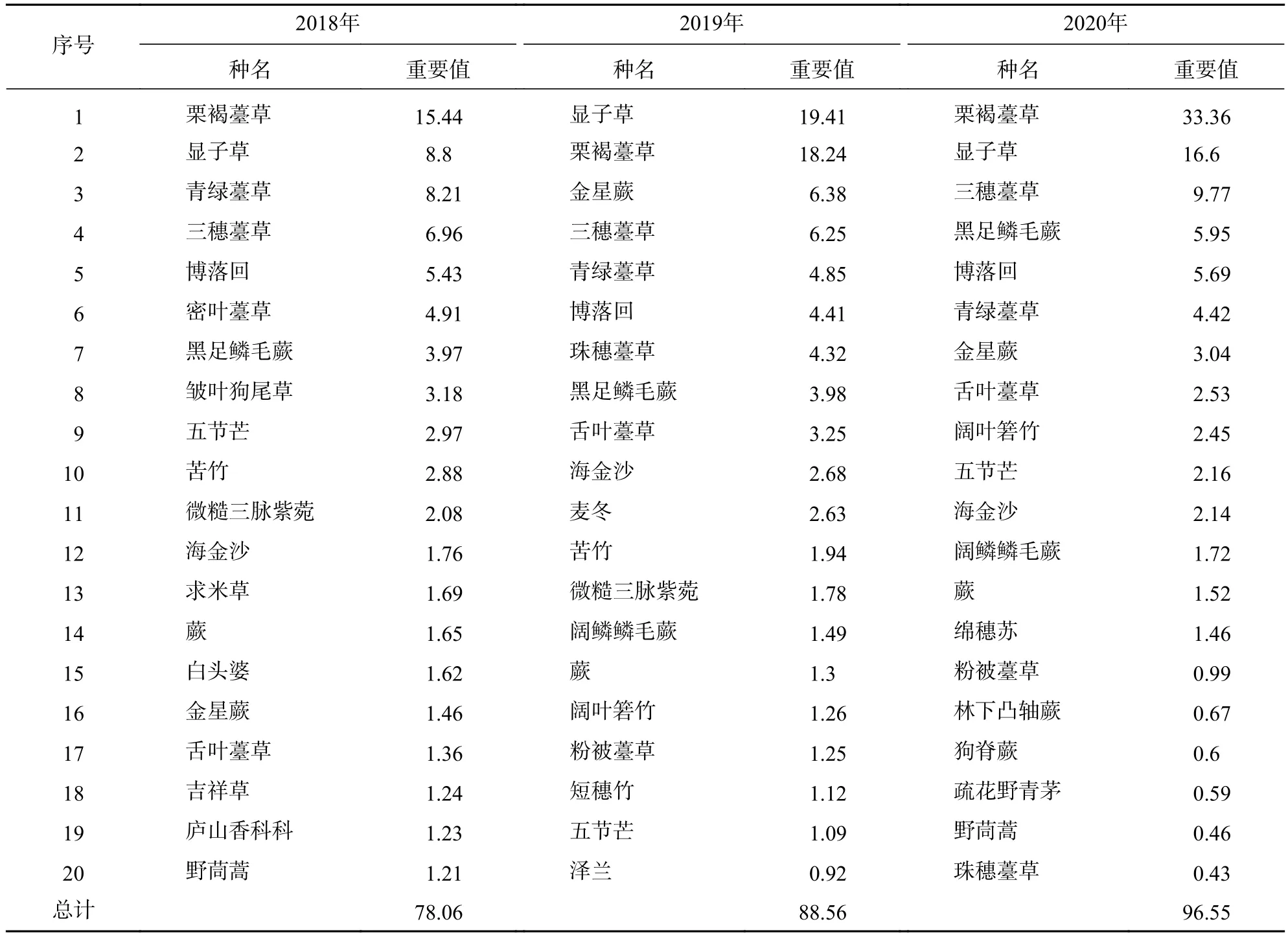
表5 2018—2020 年草本層重要值排名前 20 的物種Table 5 Top 20 species ranked by importance values in herb layer from 2018 to 2020
2.2.2 優勢種的種群動態 喬木層中的優勢樹種是森林群落的重要建造者,是森林群落的骨架[18]。由表6可知:喬木層死亡率一直大于補員率的優勢種有朵花椒、糙葉樹,其年均種群大小變化率分別為-10.9%、-11.2%,種群數量下降;補員率一直大于死亡率的優勢種有白背葉、檫木、小構樹和山雞椒,其年均種群大小變化率分別為26.7%、11.3%、32.6%、25.7%,種群數量上升;另外,有些優勢種的種群數量如臭椿、臭辣樹、掌葉覆盆子、棘莖楤木表現為先增后減,其中掌葉覆盆子在2019—2020年的死亡率高達359.0%。
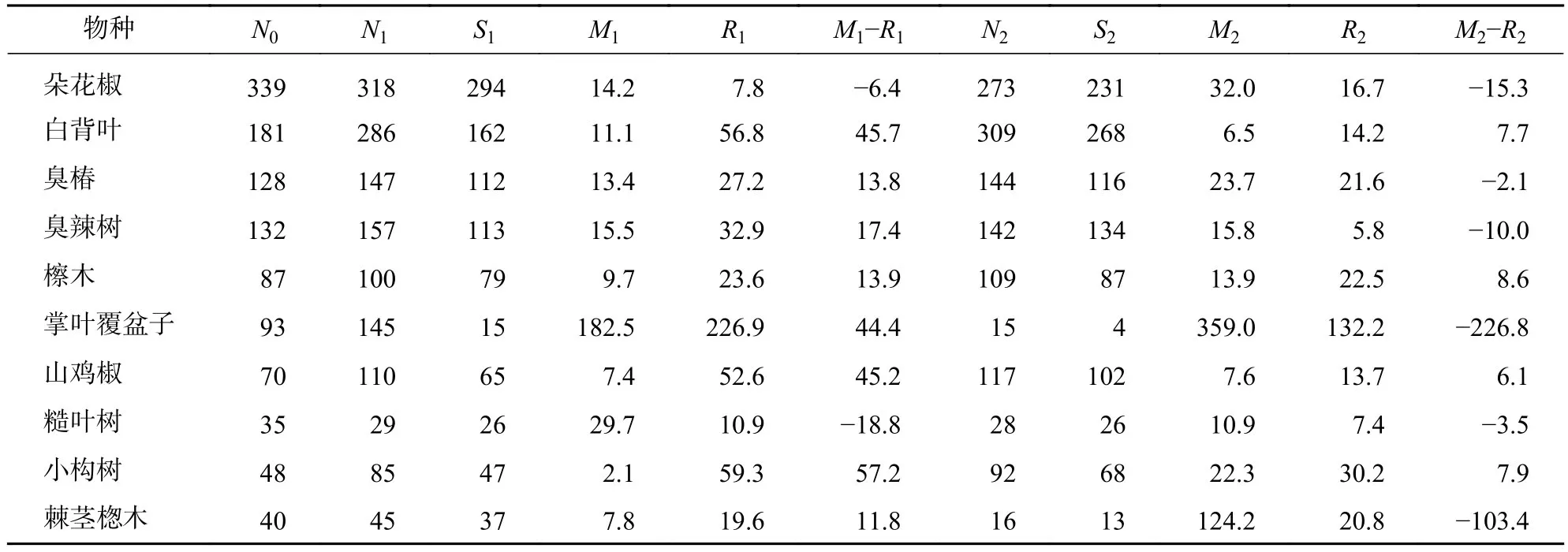
表6 優勢種的種群動態Table 6 Dynamics of dominant species
2.3 樹種功能特征的組成動態
2.3.1 葉生活型 由表7可知:2018—2020年常綠和落葉樹種的種數均增加2種,并且比例變化不大;從株數變化來看,常綠樹種的株數增加58株,所占比例逐年小幅上升,落葉樹種的株數增加278株,所占比例逐年小幅降低。這是因為落葉樹種的葉片氮、磷較高,光合作用能力較強。這些特征有助于其在光照、空間等資源充足的環境中生長發育,但隨著群落的恢復,耐受低資源環境特征的常綠樹種逐漸增多。
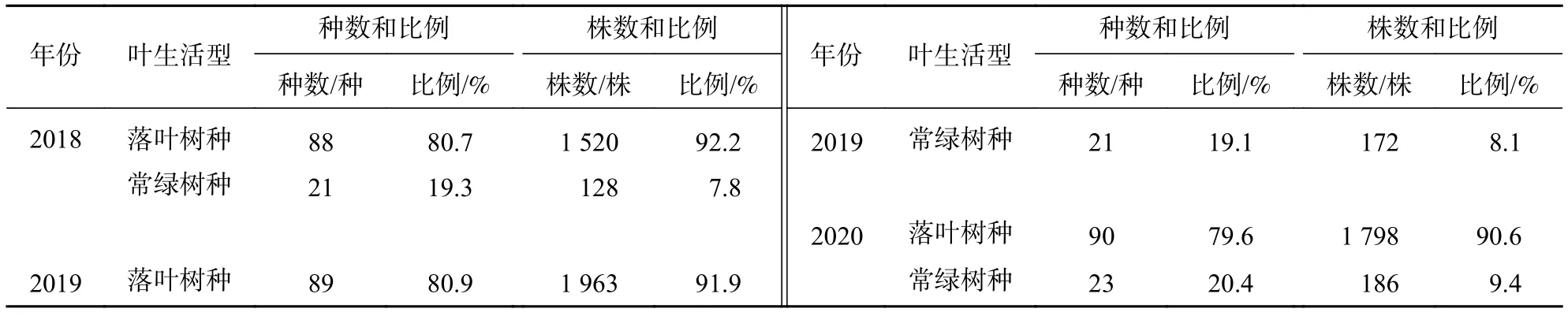
表7 2018—2020 年群落樹種葉生活型的組成情況Table 7 Composition of leaf life forms of tree species from 2018 to 2020
2.3.2 生長型 由表8可知:2018—2020年喬木樹種的種數增加6種,小喬木、灌木均減少1種,但各生長型樹種的種數占比變化不明顯。從株數變化來看,喬木、灌木的株數均先增后減,其株數占比分別下降了4.4%、1.9%,小喬木的株數一直增加,其株數占比上升6.3%。
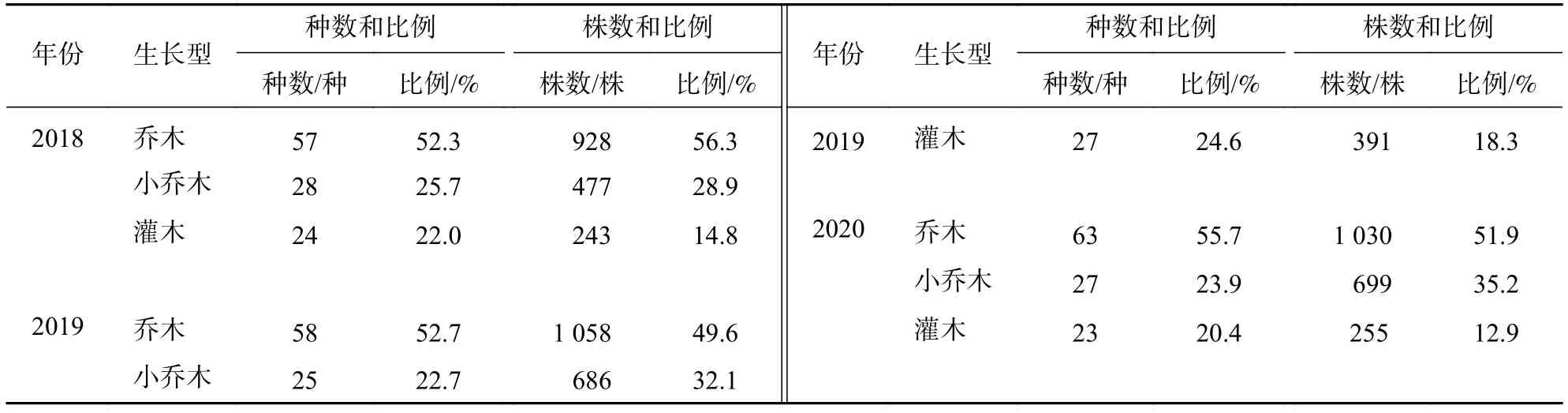
表8 2018—2020 年群落樹種生長型的組成情況Table 8 Composition of tree species growth form of community from 2018 to 2020
2.3.3 光的耐受性 由表9可知:不同光耐受性樹種種數年均變幅為1~2株,并且占比變化不明顯。從樹種的株數變化來看,陽性樹種在群落占據主導地位,其株數占比為87.7%~91.1%,并且呈現逐年小幅降低的趨勢;中性與陰性樹種株數均增加緩慢,3 a間其株數占比分別增加了1.5%、1.9%。
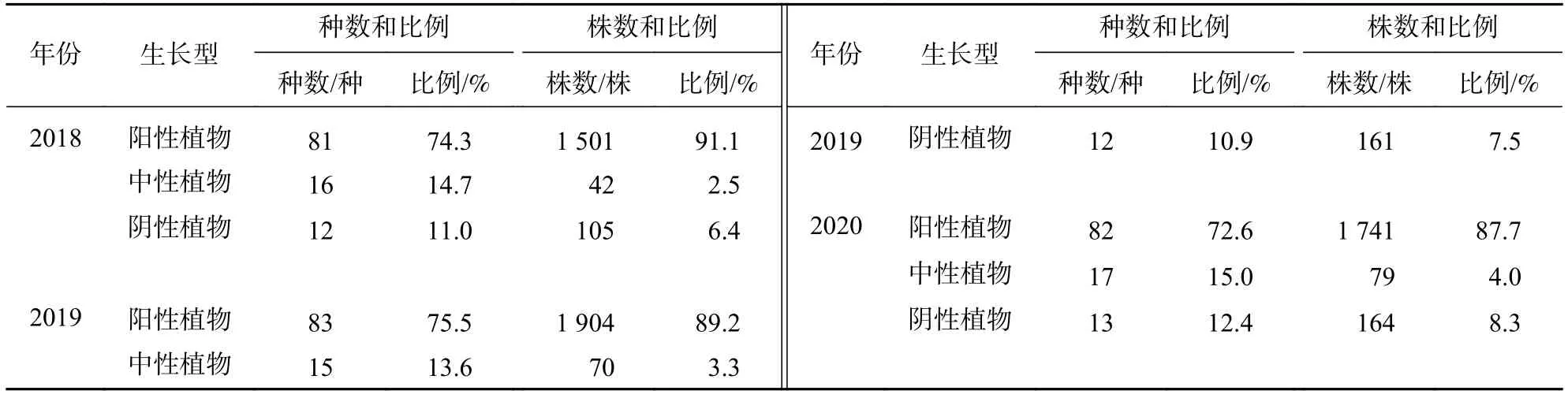
表9 2018—2020 年群落樹種光耐受性的組成情況Table 9 Composition of light tolerance of tree species from 2018 to 2020
2.4 物種多樣性的動態變化
2.4.1 喬木層物種多樣性的動態變化 由表10可知:喬木層各多樣性指數均無顯著差異,并且呈現逐年小幅上升的趨勢,具體表現為Shannon-Wiener指數、Simpson指數、Pielou均勻度指數、Margalef豐富度指數在2018—2020年的年均增幅分別為3.8%、1.3%、0.4%、7.9%。這表明在群落早期恢復過程中喬木層的物種組成與分布較為穩定。
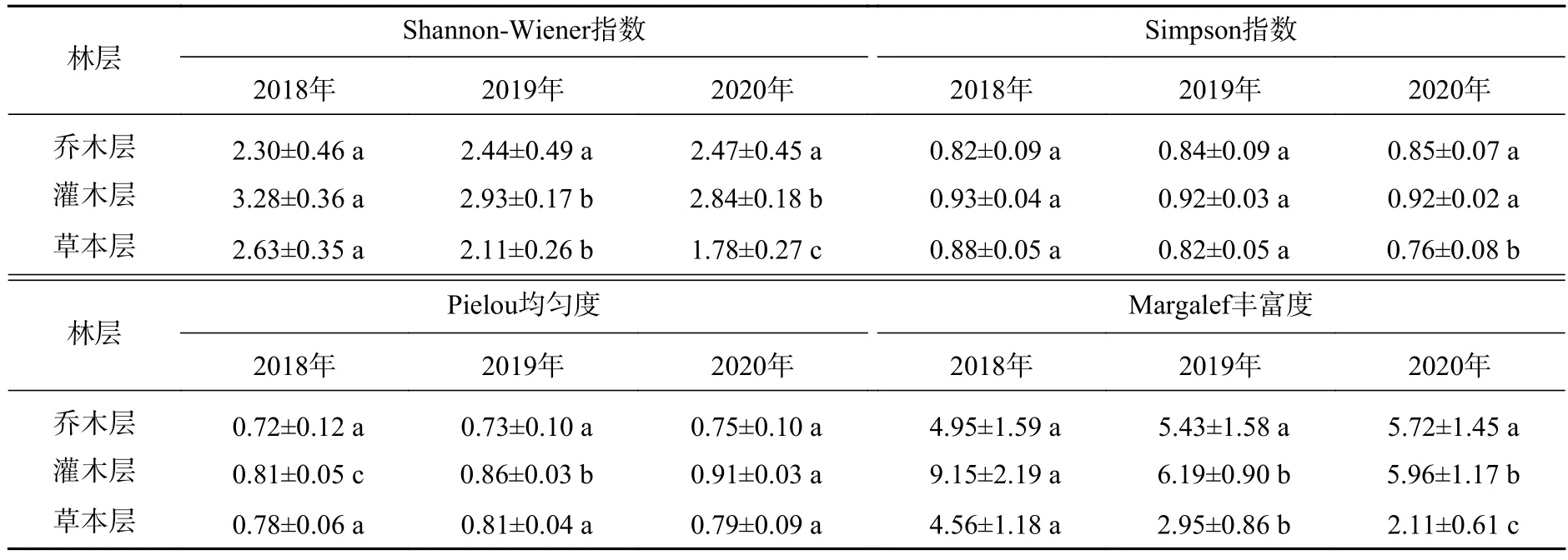
表10 物種多樣性的變化Table 10 Change of species diversity
2.4.2 灌木層物種多樣性的動態變化 灌木層 Shannon-Wiener指數和 Margalef豐富度指數在 2018—2019年顯著降低(P<0.05),降低值分別為0.35、2.97。Pielou均勻度逐年顯著增高(P<0.05),每年的增值分別為0.05、0.06。這說明某些先鋒種類獨占優勢的程度趨于逐漸減弱,物種的分布更趨于均勻。Simpson指數無顯著差異。
2.4.3 草本層物種多樣性的動態變化 草本層的Shannon-Wiener指數和Margalef豐富度指數每年均顯著降低(P<0.05),其年均降幅分別為16.2%、26.9%。這表明群落中的先鋒陽性草本植物逐漸受到灌木和喬木的遮蔭而逐漸退出群落。Simpson指數在2018—2019年無顯著差異,在2019—2020年顯著降低(P<0.05),降幅為7.5%。Pielou均勻度指數無顯著差異。
3 討論
3.1 群落物種組成豐富
群落科、屬、種和物種多樣性指數的動態變化表明:毛竹林皆伐后群落自然恢復過程中物種組成豐富。本研究喬木層植物共有45科86屬,與同區域的科屬數量相比偏高[19]。物種多樣性指數體現了群落的結構類型、生境差異以及植被的恢復程度。由于恢復演替過程中生物與環境、生物與生物間復雜的相互作用,不同物種會出現死亡或新增,物種多樣性隨之發生變化。本研究喬木層物種多樣性指數均呈現小幅增加的趨勢,與白尚斌等[5]去除針闊混交林中的毛竹時多樣性指數的變化一致,同時與亞熱帶常綠闊葉林在演替過程中物種多樣性的變化趨勢一致[8]。灌木層的Shannon-Wiener指數和Margalef豐富度指數逐年下降,這與岷江冷杉林Abies fabri皆伐后群落次生演替前期的變化趨勢一致,但草本層的Simpson指數變化與其相反[20]。毛竹林皆伐后群落的灌木、草本的多樣性指數較毛竹純林來說明顯增加[21],因此為恢復天目山國家級自然保護區的物種多樣性,首先要控制毛竹林的蔓延,然后進一步對毛竹林進行皆伐處理,同時注意清理竹筍、竹鞭,防止毛竹再生。
群落喬木層、灌木層、草本層均有只含1個種的單種屬,同時含有4種以上的屬,說明本研究區生境具有過渡性,不同屬的物種都能生長[22];另一方面,皆伐可以增加生境異質性[23]。通常認為不同樹種能在同一區域共存,是由于這些樹種的一些生態位有差異,且能協同進化形成更新特征的集聚性共存機制。這種共存機制由生物和非生物環境資源的空間異質性決定,空間異質性使樹種各自占據適生的空間而協同存在于同一區域[24]。
3.2 物種更替速率
森林群落的演替過程以各優勢種群的動態為其表現特征。群落喬木層優勢種重要值排名前10位的樹種均為落葉陽性樹種。這是因為在次生林早期演替階段,幼苗的新增主要依靠種子傳播。毛竹皆伐前種子通過風媒、水媒、動物傳播等方式進入研究區形成地下種子庫,所以皆伐后群落陽性落葉樹種以及耐蔭性較強的中性、陰性樹種均萌發。由于皆伐后研究區的地面光照強度增加、溫度升高、濕度降低,為群落中陽生性樹種的生長發展提供了機會,以至發展成為群落的優勢種[18]。
喬木層大部分優勢種的死亡率與補員率均偏高,說明群落物種更新速率較快,演替強度大。這可能是因為毛竹各器官通過化感作用釋放的物質使林木種子萌發困難,根系難以伸長,在毛竹皆伐后,大量種子得以萌發[25]。同時,林木個體為了爭奪更多養分,林木間自然稀疏現象明顯加劇[26]。除了喬木層優勢種更替速率較快外,灌草層的物種更替強度也很大,主要表現在株數的大幅下降和某些優勢種重要值的顯著增加。從常綠樹種的重要值變化來看,2018年常綠樹種的重要值總計為6.89,2020年重要值為7.12。這與沈琪等[27]關于浙江省生態公益林植被恢復過程中樹種重要值變化趨勢的研究結果一致,并且常綠樹種重要值增值較低,說明自然恢復為常綠闊葉林的速率慢。
3.3 群落結構和外貌特征的變化
群落外貌是是植物群落對各種生境因素綜合反應的體現。常綠和落葉樹種在耐旱性、耐蔭性、受干擾后的恢復能力等方面存在差異,其株數動態變化反映了群落的外貌特征[28]。調查結果表明:群落常綠樹種與落葉樹種株數占比變化不大,說明群落外貌在短期內無變化。常綠樹種的種數占總物種數的19.1%~20.4%,相對天童山[29]、古田山[30]常綠樹種物種數占比偏低。群落恢復過程中常綠樹種的種數和株數均緩慢增加,與天目山常綠落葉闊葉混交林喬木層群落動態研究結果一致[31]。這是因為隨著群落恢復,林分逐漸郁閉,為常綠樹種的萌發、生長提供有利條件[32]。
不同生長型及光耐受性樹種的動態變化反映了群落的結構特征,不同生長型樹種所占比例可以反映群落的垂直結構。群落恢復過程中,不同生長型樹種的種數與株數占比從大到小均為喬木、小喬木、灌木,說明喬木層物種豐富度與多度隨著高度的增加而增加。光耐受性是植物更新需求和生態位分化的重要指標之一[33],在演替初期主要體現在林冠下層的物種更新。本研究中陽性樹種株數占比下降3.3%,中性樹種上升1.4%,陰性樹種上升1.9%,表現為陽性樹種逐步由耐蔭樹種取代。這與汪殷華等[34]關于古田山常綠闊葉林主要樹種更新動態的變化規律基本一致。同樣,灌木層的優勢種表現出耐蔭種替代陽性樹種,草本層的優勢種如莎草科的栗褐薹草以及禾本科的顯子草等耐潮濕、耐蔭種的優勢度顯著增加。這是因為喬木作為群落的主要物種,在改變群落水分、養分、光照等環境條件中發揮著極其重要的作用。隨著林分郁閉度上升,耐蔭植物開始在林下萌發并擴張[35]。
4 結論
天目山毛竹林皆伐后的群落自然恢復過程中,物種組成豐富,物種多樣性恢復效果明顯,因此,毛竹林旨伐是恢復和保護天目山國家級自然保護區生物多樣性的可行方法,可為亞熱帶毛竹林蔓延擴張區域生態退化修復和生物多樣性保護提供參考。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
