銀黑狐MC1R基因核心啟動子區的鑒定
2022-07-27 03:26:40劉華云徐桂利張天浩謝遇春段玲欣劉錚鑄鞏元芳
中國畜牧獸醫 2022年7期
劉華云,張 磊,徐桂利,張天浩,謝遇春,段玲欣,劉錚鑄,鞏元芳
(河北科技師范學院動物科技學院,河北省特色動物種質資源挖掘與創新重點實驗室,秦皇島066004)
狐貍是中國重要的毛皮動物,分狐屬和北極狐屬2個屬,銀黑狐是狐屬狐中的一個種[1]。狐貍毛色不僅是識別狐種的重要表型性狀,也是衡量其毛皮質量的重要經濟性狀。黑色素是哺乳動物毛色形成的重要物質基礎,分為真黑色素和褐黑色素,真黑色素使毛色呈黑色,褐黑色素使毛色呈黃色或紅色,真黑色素和褐黑色素的分布決定哺乳動物毛色的深淺。黑色素的合成是一個復雜的調控網絡,受多個基因的調控[2]。黑素皮質激素受體1(melanocortin 1 receptor,MC1R)基因是參與黑色素合成的重要功能基因,該基因編碼區只有1個外顯子,通過調節cAMP通路促進黑色素表達中真黑色素的合成[3]。
研究表明,MC1R基因主要存在于哺乳動物毛囊和皮膚的黑色素細胞中,與皮膚或被毛色素的沉積緊密相關[4-5]。劉玲玲等[6]利用全基因組關聯分析發現MC1R基因與馬毛色性狀有關。王曉薇等[7]研究發現,MC1R基因在黑斑灘羊皮膚組織中表達量極顯著高于純白灘羊、顯著高于褐斑灘羊,說明MC1R基因的高表達有利于真黑色素的形成,進而促進細胞中黑色素含量的增加。南瑞鵬等[8]研究發現,MC1R基因在羊駝毛囊干細胞中高效表達,間接促進小眼畸形相關轉錄因子(microphthalmia-associated transcription factor,MITF)和酪氨酸酶(tyrosinase,TYR)基因的表達,說明羊駝毛色形成受多個基因的調控。因此,MC1R基因在動物毛色合成過程中有著重要的調控作用,其通過促進真黑色素的合成繼而影響動物深毛色的形成。
基因在真核生物中的表達是一個復雜的過程,依賴于多層次的調控,其中轉錄水平的調控是影響真核生物基因表達的重要一步。啟動子是基因的一個組成部分,控制基因表達(轉錄)的起始時間和表達程度,通過對啟動子區的研究有助于了解基因的結構、功能及表達調控機制[9-11]。目前,MC1R基因啟動子區在人、小鼠中研究較為成熟[12],但在狐貍中鮮有報道。鑒于此,本研究利用PCR擴增、生物信息學預測及雙熒光素酶報告基因檢測等技術對銀黑狐MC1R基因核心啟動子區進行了鑒定與分析,以期為深入研究該基因對狐貍毛色的調控機制提供參考。
1 材料與方法
1.1 材料
1.1.1 試驗動物 銀黑狐來自秦皇島市昌黎金島育種場,隨機選取1只健康成年銀黑狐,剪去背部被毛,用取膚器采集背部皮膚組織樣品并立即放入液氮中,后轉入―80 ℃保存備用。
1.1.2 主要試劑 大腸桿菌DH5α感受態細胞、pMD19-T載體、T4 DNA連接酶、限制性內切酶SacⅠ和Hind Ⅲ均購自TaKaRa公司;pGL3-Basic表達載體、pRL-TK載體和雙熒光酶檢測試劑盒Dual-Luciferase?Reporter Assay System均購自Promega公司;皮膚黑色素瘤細胞株(B16細胞)購自北京協和細胞資源中心;DMEM培養基(高糖型)購自HyClone公司;胎牛血清購自Gibco公司。
1.2 基因組DNA提取
參照Sambrook等[11]方法從銀黑狐皮膚組織樣中提取基因組DNA,TE溶解,―20 ℃保存備用。
1.3 銀黑狐MC1R基因5′-UTR區序列獲取
1.3.1 引物設計及合成 根據GenBank中犬MC1R基因5′-UTR區序列(登錄號:NM_001014282)和銀黑狐MC1R基因編碼區序列(登錄號:KJ489060),利用Primer Premier 3.0引物設計軟件對銀黑狐MC1R基因5′-UTR區設計引物,F:5′-GAGCTCTGTTCTCTCAGCCTCCTCAC-3′;R:5′-AAGCTTGAAGTGAGGGGTGGTTGG-3′,預計擴增片段大小為2 781 bp。引物由生工生物工程(上海)股份有限公司合成。
1.3.2 PCR擴增 PCR反應體系50 μL:銀黑狐基因組DNA 1 μL,10×PCR Buffer 5 μL,2.5 mmol/L dNTPs 4 μL,上、下游引物各1 μL,Taq酶1 μL,ddH2O 37 μL。PCR反應條件:94 ℃預變性5 min;98 ℃變性10 s,55 ℃退火15 s,68 ℃延伸3 min,共30個循環;72 ℃延伸10 min;4 ℃保存。PCR擴增產物用1.5%瓊脂糖凝膠電泳檢測,取鑒定正確的擴增產物純化回收后克隆至pMD19-T載體,連接體系10 μL:載體 1 μL,DNA回收產物 4 μL,Solution Ⅰ 5 μL。離心混勻后,16 ℃過夜連接。連接產物轉化大腸桿菌DH5α感受態細胞,37 ℃搖床培養,將菌液PCR鑒定為陽性的樣品送生工生物工程(上海)股份有限公司進行測序。
1.4 銀黑狐MC1R基因核心啟動子區預測分析
利用3個在線生物學軟件Neural Network Promoter Prediction(http:∥www.fruitfly.org/seq_tools/promoter.html)、Promoter 2.0(http:∥www.cbs.dtu.dk/services/Promoter/)和TSSW(http:∥www.softberry.com/berry.phtml?topic=tssw&group=programs&subgroup=promoter)依次對銀黑狐MC1R基因啟動子核心區進行預測。
1.5 銀黑狐MC1R基因核心啟動子區確定
在軟件預測的基礎上,利用熒光素酶報告基因檢測技術進一步確定銀黑狐MC1R基因的核心啟動子區。
1.5.1 引物設計及PCR擴增 根據軟件預測結果,在起始密碼子上游依次截取10個片段(圖1),分別設計相應的引物(表1),在上、下游引物的5′-端分別引入SacⅠ、Hind Ⅲ酶切位點及相應的保護堿基以便于克隆。引物均由生工生物工程(上海)股份有限公司合成。缺失片段PCR擴增體系與程序同1.3.2,PCR產物用1.5%瓊脂糖凝膠電泳檢測。
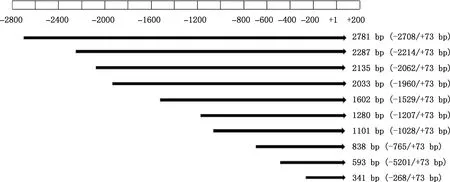
圖1 銀黑狐MC1R基因啟動子缺失片段示意圖

表1 銀黑狐MC1R基因啟動子區缺失片段引物
1.5.2 目的片段克隆及鑒定 PCR產物經瓊脂糖凝膠電泳純化回收后連接至pMD19-T載體上,具體方法同1.3.2,將菌液PCR鑒定為陽性的樣品送往華大基因公司進行測序。
1.5.3 重組質粒構建及鑒定 對于不同長度的重組載體pMD19-T和pGL3-Basic,利用SacⅠ和Hind Ⅲ進行雙酶切。酶切體系50 μL:質粒 13 μL,Buffer 5 μL,SacⅠ和Hind Ⅲ各3 μL,ddH2O補足體系。37 ℃酶切6 h。酶切產物進行切膠回收并進行雙酶切驗證,驗證后選取相應擴增片段與pGL3-Basic連接,連接體系10 μL:目的片段質粒4 μL,pGL3-Basic 1 μL,Solution Ⅰ 5 μL。16 ℃過夜連接。共構建10個熒光素酶報告基因重組質粒,分別命名為:pGL3-MC1RP0(-2 708/+73 bp)、pGL3-MC1RP1(-2 214/+73 bp)、pGL3-MC1RP2(-2 062/+73 bp)、pGL3-MC1RP3(-1 960/+73 bp)、pGL3-MC1RP4(-1 529/+73 bp)、pGL3-MC1RP5(-1 207/+73 bp)、pGL3-MC1RP6(-1 028/+73 bp)、pGL3-MC1RP7(-765/+73 bp)、pGL3-MC1RP8(-520/+73 bp)和pGL3-MC1RP9(-268/+73 bp)。菌液經PCR擴增及酶切鑒定后,選取陽性重組質粒測序。對陽性菌液提取無內毒素質粒,并檢測其濃度和純度。
1.5.4 細胞培養與重組質粒瞬時轉染 取出B16細胞復蘇后,接種于含10%胎牛血清的高糖DMEM培養基中,置于37 ℃、5% CO2恒溫細胞培養箱中培養,細胞每1~2 d用胰酶消化傳代。復蘇后的細胞培養3代后用于重組質粒的瞬時轉染(瞬轉)試驗。轉染前12~24 h,將生長狀態良好的B16細胞接種于24孔板內,每孔接種5×104~10×104個細胞。當細胞匯合度為80%~90%時,按照Xfect Transfection Reagent操作步驟進行相應質粒轉染;轉染時每孔加入質粒總量為5 μg(pGL3相關重組質粒與內參質粒pRL-TK質量比為100∶1),轉染試劑量為15 μL。對照組為pGL3-Basic與pRL-TK共轉染,試驗組為pGL3相關重組質粒與pRL-TK共轉染。每組進行3個復孔的轉染,且進行3次以上重復試驗。轉染48 h后收集細胞,用于雙熒光素酶的活性檢測。
1.5.5 雙熒光素酶活性檢測 根據雙熒光酶檢測試劑盒Dual-Luciferase?Reporter Assay System說明書進行報告基因表達水平的檢測,具體方法為:棄掉24孔板中的細胞培養液,用1×PBS洗滌細胞2次;加入150 μL細胞裂解液(1×PLB),輕輕吹打完全細胞;室溫放置15 min后,收集細胞裂解產物;取20 μL細胞裂解產物至熒光測定管中,加入100 μL檢測試劑(firefly luciferase,LARⅡ),混勻后使用GloMax-MultiJr單管型多功能檢測儀檢測螢火蟲螢光素酶活性;迅速加入100 μL Stop&GLO試劑,測定作為內標的Renilla熒光素酶活性。螢火蟲熒光素酶活性/Renilla熒光素酶活性的比值即為熒光素酶的相對活性(relative luciferase activity,RLA)。
1.5.6 統計分析 利用SPSS 22.0統計軟件對數據進行單因素方差分析(One-Way ANOVA),數據用平均值±標準差表示,P<0.05代表組間存在統計學差異,具有統計學意義。
2 結 果
2.1 銀黑狐MC1R基因5′-UTR區序列獲取
以銀黑狐基因組DNA為模板進行PCR擴增,擴增產物經1.5%瓊脂糖凝膠電泳檢測后,有一條長約2 781 bp的清晰條帶(圖2),與目的片段大小相符。PCR擴增產物克隆到pMD19-T載體后送測序,結果顯示,克隆所獲目的片段與GenBank中犬MC1R基因5′-UTR區序列(登錄號:NM_001014282)相似性達93.97%,表明已成功獲得銀黑狐MC1R基因2 781 bp(-2 708/+73 bp)的5′-UTR區序列。
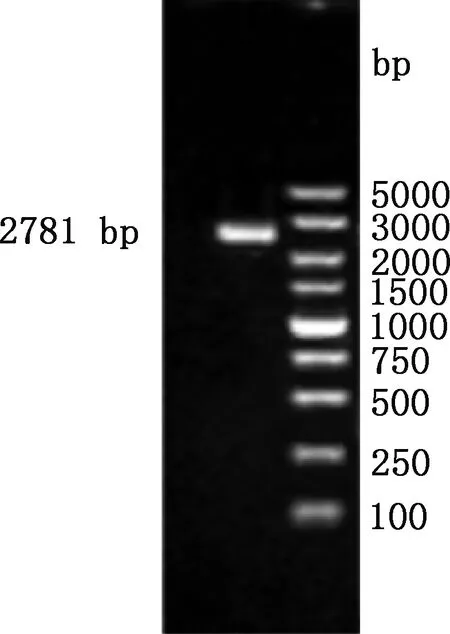
圖2 銀黑狐MC1R基因5′-UTR區PCR擴增結果
2.2 銀黑狐MC1R基因啟動子活性區預測分析
通過在線軟件對銀黑狐MC1R基因5′-UTR區(-2 708/+73 bp)進行預測分析。利用Neural Network Promoter Prediction預測到4個啟動子活性區域:-1 964/-1 915 bp(得分0.93)、-1 834/-1 785 bp(得分0.97)、-1 414/-1 365 bp(得分0.82)和-596/-548 bp(得分0.92);利用Promoter 2.0軟件預測啟動子位置為-1 209 bp(得分0.70)和-309 bp(得分0.71);利用TSSW軟件預測啟動子位置為-2 323 bp(得分17.88)和-359 bp(得分4.86)。3個軟件是基于不同算法對啟動子區的預測,提示-596/+73 bp可作為參考的候選核心啟動子區域。
2.3 銀黑狐MC1R基因啟動子缺失序列重組質粒酶切鑒定
以2 781 bp(-2 708/+73 bp)PCR產物為模板進行缺失片段的PCR擴增,所得片段大小分別為2 781、2 287、2 135、2 033、1 602、1 280、1 101、838、593和341 bp(圖3),均與預期大小相符。PCR擴增產物純化回收后分別克隆到pGL3-Basic質粒中,用SacⅠ與Hind Ⅲ雙酶切進行重組質粒鑒定,均獲得與預期相符的目的片段(圖4)。選出陽性重組質粒測序,結果顯示,克隆到的不同長度目的片段與所得狐貍MC1R基因序列相似性達99%。說明成功構建了10個包含銀黑狐MC1R基因不同長度序列的重組質粒。
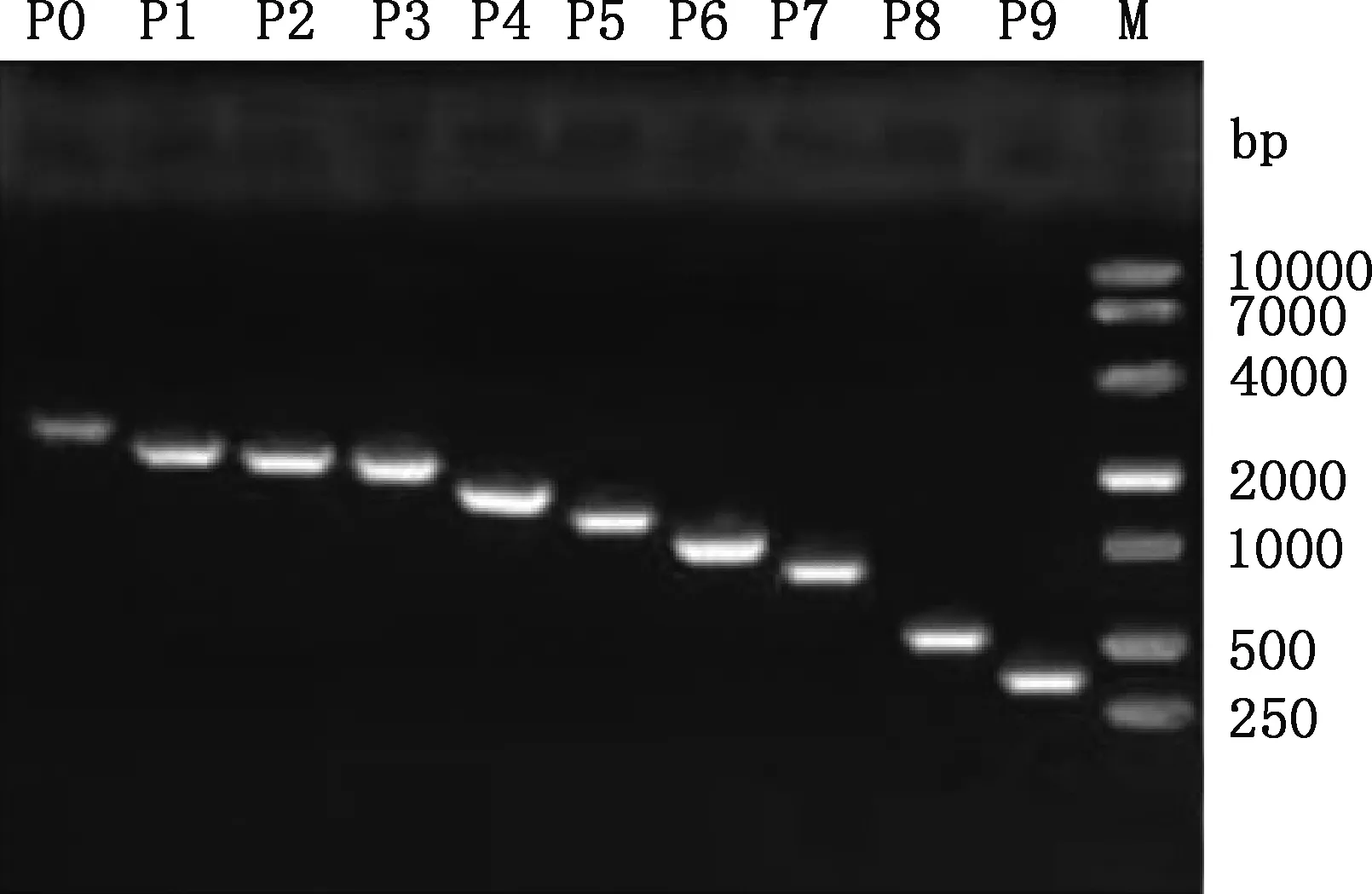
P0~P9,MC1R基因啟動子不同缺失片段;M,DM10000 DNA Marker
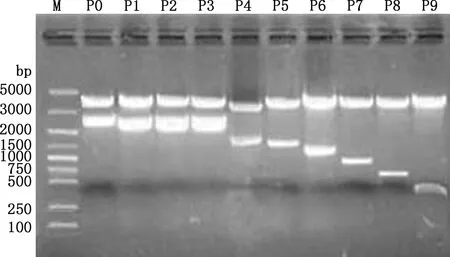
M,DM5000 DNA Marker;P0~P9,不同缺失片段重組質粒的SacⅠ與Hind Ⅲ雙酶切產物
2.4 雙熒光素酶活性檢測
將10個啟動子缺失片段的重組質粒、pGL3-Basic質粒分別與phRL-TK質粒瞬時共轉染B16細胞48 h后收集細胞,計算RLA,分析各啟動子片段的活性,進而確定MC1R基因的啟動子位置,結果見圖5。由圖5可知,9個啟動子缺失片段的重組質粒轉染后的熒光素酶相對活性均較pGL3-Basic空質粒轉染組顯著或極顯著增加(P<0.05;P<0.01),表明報告基因在B16細胞中具有啟動活性,啟動子片段可啟動熒光素酶的表達。通過比較10個不同長度啟動子片段的活性值發現,pGL3-MC1RP8(-520/+73 bp)組的熒光素酶活性值最高,說明銀黑狐MC1R基因的核心啟動子區存在于pGL3-MC1RP8(-520/+73 bp)區域內。

與pGL3-Basic相比,*,差異顯著(P<0.05);**,差異極顯著(P<0.01)
3 討 論
銀黑狐是赤狐在自然條件下產生的毛色突變體,被毛黑白相間,有一層霧狀的針毛,是裘皮服裝的主要飾品。近年來,隨著人們生活水平的不斷提高以及綠色環保理念的普及,生產天然優質彩色狐皮成為養狐業的終極目標,因此對其毛色調控基因的研究成為熱點。目前發現調控狐貍毛色的基因有MC1R、TYRP1、Agouti、TYR、CBD、Extension、SILV、KIT、MITF等,其中MC1R是最早被發現且證實可調控動物毛色的基因之一[13],前人陸續在狐貍[14-16]、水貂[17]、家犬[18-19]、兔[20-22]、馬[23]、羊[24-26]、豬[27-29]、牛[30-33]和羊駝[34]等物種中進行了相關研究。基因在真核生物中的表達調控可分為多個水平,其中最主要的就是轉錄水平調控,而啟動子調控也是轉錄環節的關鍵,因此針對啟動子功能區域開展相關研究對了解基因的表達調控機制具有重要意義[2]。
本研究成功獲取了銀黑狐MC1R基因5′-UTR區2 781 bp的DNA序列,構建了10個不同缺失長度的啟動子熒光素酶表達載體,活性檢測表明,10個片段均有一定的啟動子活性。研究表明,克隆片段僅含有TSS上游的區域很多情況會導致假陰性結果,若克隆片段包含TSS、ATG及部分編碼區會使陽性率大大提高[2]。本研究克隆的10個片段均符合上述要求。利用3種不同算法的在線生物學軟件對銀黑狐MC1R基因的啟動子區進行預測發現,-596/+73 bp可能為該基因的核心啟動子區。雙熒光素酶活性結果顯示,pGL3-MC1RP8(-520/+73 bp)的活性值最高,提示-520/+73 bp為銀黑狐MC1R基因的核心啟動子區,與軟件預測結果相符。
研究發現,在狐貍MC1R基因5′-UTR區的-36/-41、-100/-105、-123/-128、-278/-283和-507/-512 bp區域各存在1個E-box,在-91/-96 bp區域存在1個GC-box,在-451/-457 bp區域存在1個BRE調控元件[16]。E-box和GC-box是上游調控區重要的順式作用元件,參與促進基因的轉錄調控。BRE是基因核心啟動子區重要的調控元件,是TFIIB的特異性識別元件,而TFIIB基因的一個基本轉錄起始因子可促進RNA聚合酶正確定位,起“定位因子”的作用[35]。研究中E-box、GC-box及BRE調控元件均位于-520/+73 bp區域內,提示通過雙熒光素酶檢測技術確定的活性區域(-520/+73 bp)是銀黑狐MC1R基因的核心啟動子區。研究發現,黑腹田鼠MC1R基因編碼區上游500 bp左右為其核心啟動子區[36];人MC1R基因最小啟動子區域在-517/-282 bp[37],均與本研究結果基本吻合。
真核生物的表達是順式作用元件和反式作用因子共同作用的結果,此過程復雜而有序。轉錄水平調控是基因表達的第一步,是轉錄因子與轉錄因子結合位點進行特異性結合,參與調控靶基因轉錄效率的過程[38]。本研究構建的10個缺失報告基因載體中pGL3-MC1RP8(-520/+73 bp)的活性值最高,提示-520/+73 bp區域內含有重要的調控元件,可調控狐貍MC1R基因啟動子的活性,本試驗結果為深入研究該基因奠定了理論基礎,也為其他動物毛色基因的研究提供了參考。下一步應利用點突變、染色質免疫共沉淀、DNA足紋分析和凝膠阻滯分析等技術驗證啟動子區轉錄因子的真正功能,進而深入了解狐貍MC1R基因的轉錄調控機制。
4 結 論
本研究利用PCR擴增、生物學軟件預測及雙熒光素酶報告基因檢測等技術成功確定了銀黑狐MC1R基因的核心啟動子區(-520/+73 bp)。
