山羊RORα基因的克隆、表達載體構建及功能分析
2022-07-07 08:23:18高登科趙泓淙靳亞平陳華濤
畜牧獸醫學報 2022年6期
關鍵詞:小鼠
高登科,趙泓淙,董 浩,靳亞平,陳華濤*
(1. 西北農林科技大學動物醫學院,楊凌 712100; 2. 西北農林科技大學 農業農村部動物生物技術重點實驗室,楊凌 712100)
千百萬年來地球圍繞地軸自轉,形成了周期性的明/暗循環,賦予了地球表面光照時間、溫度和能量等外部環境的晝夜節律性變化。這些節律性變化使地球上的生物在進化壓力的長期驅動下產生了與之相適應的內在計時機制,從而導致機體大多數生理功能具有了以近似24 h為周期的節律性振蕩,被稱為晝夜節律。生物體用以預測時間變化和調節晝夜節律的內源性計時機制則被稱為生物鐘。作為一種重要的調控系統,生物鐘與機體的生長發育等生理功能的正常運轉密切相關。當生物鐘受到環境、基因突變等因素干擾時,機體的生理過程也會發生紊亂,從而導致疾病的發生。哺乳動物的生物鐘系統由位于下丘腦視交叉上核(suprachiasmatic nucleus,SCN)的中樞生物鐘和分布于機體幾乎所有細胞、組織與器官的外周生物鐘組成。SCN通過接收和整合來自視網膜下丘腦束傳來的光信號來調控機體中樞生物鐘的運轉,并通過神經和體液途徑將信號信息傳遞給大腦其他部位以及外周組織,驅動外周生物鐘的節律性振蕩,從而使機體維持生理穩態。
哺乳動物生物鐘調控機制雖然具有一定的物種差異性,但在分子水平上其主要依賴于1、、(1、2、3)、(1、2)和(α、β)等核心生物鐘基因的轉錄調控,幾乎所有的細胞都表達這些生物鐘基因,并具有產生晝夜節律性振蕩的潛能,不同物種的核心生物鐘調控機制在進化上具有高度保守性。維甲酸相關孤兒受體α(retinoic acid-related orphan receptors alpha,RORα)作為生物鐘系統的重要組成部分,在維持生物鐘的正常運轉過程中發揮重要作用。在生物鐘的轉錄反饋環路中,RORα通過與1基因啟動子上特定的DNA反應元件即維甲酸相關孤兒受體反應元件(RORE:由一段(G/A)GGTCA核心序列和一段6 bp富含A/T的前導序列構成)結合來激活基因轉錄。同時,REV-ERBs作為轉錄抑制因子,通過與RORα競爭性結合RORE元件來抑制1的轉錄。此外,α自身的轉錄也受到一些上游因子的調節,如DBP/NFIL3會與α基因啟動子上的D-box元件結合來調控其轉錄。反過來,和3的轉錄也分別受到BMAL1/CLOCK和RORα的調控。總之,α與1和等核心生物鐘基因一起形成了多個轉錄翻譯反饋環路,共同維持著生物鐘系統的正常運轉。除此之外,有研究發現RORα作為脂質和脂蛋白代謝、脂肪生成和血管炎癥的重要調節因子,在機體脂代謝和炎癥反應過程中發揮重要調控作用。研究表明,RORα還在細胞凋亡以及腫瘤發生過程中扮演著重要角色。
然而,上述文獻證據大多是基于模式動物小鼠來研究的,RORα蛋白的功能是否具有種屬特異性尚不清楚。山羊作為人類最早馴化的家畜之一,在人類農業文明和經濟發展中發揮重要作用,一直是畜牧獸醫領域關注的熱點。有研究報道,絨山羊的產絨性能受到晝夜節律生物鐘的影響。然而,山羊生物鐘系統的轉錄調控機制仍未被完全解析。作為生物鐘系統的重要組成部分,α基因的全長編碼區(coding sequence,CDS)還未被克隆和深入研究。基于此,本研究旨在克隆山羊生物鐘基因α的編碼區CDS片段并構建其真核表達載體,進一步對山羊α基因進行生物信息學分析和功能的初步研究,為后續探究RORα蛋白生物學功能及其在山羊生物鐘系統的轉錄調控機制奠定基礎。
1 材料與方法
1.1 試驗材料、主要試劑及儀器
山羊肝組織取自1只12月齡健康的雄性西農薩能奶山羊;HEK293T細胞購于中國科學院細胞庫;TRIzol、PrimeSTARMax DNA Polymerase、Quick CutR I、Quick CutI、Quick CutI購自日本TaKaRa公司;反轉錄試劑盒購自AG生物工程有限公司;THUNDERBIRD SYBRqPCR Mix購自日本ToYoBo公司;ClonExpressII One Step Cloning Kit購自南京諾唯贊生物科技股份有限公司;DH5α感受態細胞、膠回收試劑盒、無內毒素質粒小提中量試劑盒、基因組DNA提取試劑盒購自北京天根生化科技有限公司;胎牛血清、Opti-MEM培養基均購自美國Gibco公司;DMEM高糖培養基購自Hyclone公司;TurboFect Transfection Reagent購自美國Thermo Fisher公司;雙熒光素酶報告實驗試劑盒購于Promega公司;BCA蛋白濃度檢測試劑盒購自江蘇凱基生物技術股份有限公司;脫脂奶粉購于美國BD公司;HRP標記的鼠抗β-actin抗體購自中國三箭公司;兔抗HA抗體購自Abcam公司;HRP共軛山羊抗兔抗體購自中國中杉金橋公司;ECL HRP底物試劑盒購自北京迪寧生物科技有限公司。實驗室具有PCR擴增儀(美國Bio-Rad公司)、熒光定量系統(美國Bio-Rad公司)、電泳儀(美國Bio-Rad公司)、凝膠成像分析系統(英國Syngene公司);核酸蛋白測定儀(美國Thermo Fisher公司)和CO培養箱(美國Thermo Fisher公司)等儀器設備。
1.2 山羊肝組織cDNA的獲取及引物設計
稱取20 mg山羊肝組織,使用TRIzol法提取總RNA,反轉錄得到cDNA。根據NCBI數據庫中山羊α基因(NM_001285652.1)的CDS區,使用Primer Premier 6.0軟件設計PCR引物。同時在引物上下游的5′端添加與pcDNA3.1-Puro-N-3HA載體R I 酶切位點處互補配對的同源序列,設計帶有同源臂的引物。另外,根據NCBI數據庫中山羊1基因啟動子(1-Promoter)序列(NC_030822.1)與1D1啟動子(1D1-Promoter)序列(NC_030826.1)(兩個基因的啟動子序列均含有RORE元件),使用Primer Premier 6.0軟件分別設計擴增山羊1-Promoter區域和1D1-Promoter區域的引物。前者在引物上下游5′端添加與pGL4.10載體I酶切位點互補配對的同源序列,后者在引物上下游5′端添加與pGL4.10載體I酶切位點互補配對的同源序列,設計帶有同源臂的引物。引物序列信息見表1,所有引物均交由西安擎科生物科技有限責任公司進行合成。

表1 引物序列信息
下劃線部分為酶切位點
The underlined parts are the sequences of the added restriction enzyme sites
1.3 pcDNA3.1-3HA-gRORα真核表達載體的構建
1.3.1 山羊α基因CDS區的克隆 以山羊的cDNA為模板擴增帶有同源臂的α基因的CDS區片段。PCR反應程序如下:在反應離心管中加入cDNA模板4 μL(50 ng · μL),上、下游引物各1 μL(10 μmol·L),2×PrimeSTAR Max Premix 25 μL,ddHO 19 μL,建立50 μL的PCR反應體系;PCR程序為:98 ℃預變性5 min;98 ℃ 變性10 s,55 ℃退火15 s,72 ℃延伸105 s,循環35次;72 ℃ 總延伸5 min。PCR產物進行瓊脂糖凝膠電泳鑒定,對符合預期的條帶按照DNA純化回收試劑盒說明書操作進行純化回收。
1.3.2 pcDNA3.1-3HA-gRORα載體的構建及鑒定 使用限制性內切酶R I將pcDNA3.1-Puro-N-3HA載體進行酶切線性化處理,并通過瓊脂糖凝膠電泳試驗檢測酶切產物,將符合預期的條帶使用膠回收試劑盒回收純化。根據ClonExpress快速重組克隆試劑盒說明書,將膠回收得到的帶有同源臂的目的基因片段與酶切線性化的真核表達載體pcDNA3.1-Puro-N-3HA進行重組連接,重組反應體系:線性化載體1.5 μL,片段3.5 μL,Exnase Ⅱ 2 μL, 5×CE Ⅱ Buffer 4 μL,加水至20 μL。將反應混合液在37 ℃條件下反應15 min以完成重組反應,得到重組產物。將重組產物按照轉化反應操作說明轉至DH5α大腸桿菌感受態細胞中,之后涂布于含氨芐霉素30 μg · mL的固體培養基中。12 h后挑取單克隆菌落,于37 ℃搖床培養12 h,使用無內毒素小提中量試劑盒提取質粒并測定濃度。隨后使用限制性內切酶R I對重組質粒進行單酶切鑒定。同時以重組質粒為模板,使用α基因的引物進行PCR鑒定。將酶切與PCR鑒定結果符合預期的重組質粒送至西安擎科生物有限公司進行測序鑒定。返回序列與NCBI數據庫中已知序列進行比對,將比對正確的質粒樣本命名為pcDNA3.1-3HA-gRORα真核表達載體。
1.3.3 重組質粒轉染HEK293T細胞 復蘇HEK293T細胞并進行傳代,以每孔4×10個細胞接種至1個6孔板,以每皿8×10個細胞接種至6個60 mm細胞培養皿。待HEK293T細胞密度達到60%左右時進行轉染。將pcDNA3.1-3HA-gRORα作為試驗組,pcDNA3.1-Puro-N-3HA作為對照組。將兩組質粒按照TurboFect轉染試劑說明書的操作步驟分別轉染細胞。6孔板中,試驗組質粒和對照組質粒分別轉染3孔,轉染后24 h收樣,用于檢測山羊α基因在mRNA水平的表達。6個60 mm 皿中,試驗組質粒和對照組質粒分別轉染3皿,轉染48 h后收樣,用于檢測山羊α基因在蛋白水平的表達。
1.3.4 實時熒光定量PCR檢測山羊α基因在mRNA水平的表達 轉染24 h后,使用TRIzol試劑提取細胞樣品總RNA,反轉錄合成cDNA,并稀釋至5 ng·μL,作為模板備用。用Primer Premier 6.0設計人與山羊通用的α引物序列以及人的引物序列(表1)。使用ToYoBo的THUNDERBIRD SYBRqPCR Mix并按說明書操作進行實時熒光定量PCR(qPCR)反應。反應體系:cDNA 5 μL,qPCR Mix 10 μL,上、下游引物各 1 μL,去離子水3 μL。qPCR反應程序:95 ℃ 3 min; 95 ℃ 10 s;60 ℃ 30 s,共40個循環;95 ℃ 15 s,60 ℃ 15 s,95.5 ℃ 5 s。使用熒光定量系統進行反應,以人的基因作為內參基因,并采用2的相對定量法對樣本進行定量分析。
1.3.5 蛋白質免疫印跡檢測山羊α基因在蛋白水平的表達 轉染48 h后,提取細胞蛋白樣品進行蛋白質免疫印跡(Western blotting,WB)試驗。具體操作:將細胞收集至離心管中,按照RIPA裂解液操作說明處理細胞得到蛋白樣品。使用BCA蛋白含量檢測試劑盒測定總蛋白濃度后加入Loading Buffer,100 ℃水浴10 min后進行SDS-PAGE凝膠電泳。電泳結束后將蛋白轉印至PVDF膜上;使用10%的脫脂奶粉封閉2 h。封閉完成后,TBST溶液中洗膜4次,每次5 min。將PVDF膜置于兔抗HA抗體工作液(1∶2 000稀釋)中,室溫搖床孵育2 h;TBST洗膜4次,每次5 min。之后將PVDF膜置于HRP標記的山羊抗兔二抗工作液(1∶5 000 稀釋)中,室溫搖床孵育2 h;TBST洗膜4次,每次5 min。 稍晾干后,將膜置于3 mL ECL發光液中作用2 min,于暗室中曝光并保存目標蛋白結果圖。將曝光后的膜取出,TBST洗膜2次,每次5 min。加入適量抗體洗脫液,室溫孵育15 min,棄去洗脫液,TBST洗膜3次,每次5 min。隨后再次進行封閉,操作與上述相同。封閉完成后,將PVDF膜置于HRP標記的β-actin抗體工作液(1∶5 000稀釋)中,室溫搖床孵育2 h;TBST洗膜4次,每次5 min。之后將膜放入ECL發光液中作用2 min,于暗室中曝光并保存目標蛋白結果圖。最后使用ImageJ軟件對轉染組和對照組的蛋白條帶分別進行灰度分析,并與β-actin蛋白進行均一化處理。
1.4 山羊RORα基因生物信息學分析
利用Ensembl在線數據庫(https://www.ensembl.org/index.html)對山羊α基因全長序列進行分析。在NCBI數據庫中查找山羊和小鼠、大鼠、人、牛、綿羊、豬、馬、斑馬魚等多個物種的α基因CDS序列(表2)。使用生物信息學軟件DNAStar對各物種α基因的CDS序列進行同源性比對;應用MEGA7軟件對各物種α基因的CDS序列進行分析并構建系統進化樹;使用在線軟件 SignalP-5.0(https://services.healthtech.dtu.dk/service.php?SignalP-5.0)、TMHMM Server v.2.0(http://www.cbs.dtu.dk /services/ TMHMM/)、ProtParam(https://web.expasy.org/protparam/)、ProtScale(https://web.expasy.org/ protscale/)、SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL(https://swissmodel.expasy.org/)對山羊RORα蛋白的理化性質、親/疏水性、信號肽、跨膜結構、蛋白二級以及三級結構進行生物信息學預測和分析。最后,使用pyMOL軟件對山羊、小鼠和人三者的RORα蛋白三維結構進行相似性比較。
1.5 pGL4.10-BMAL1-Promoter-Luc與pGL4.10-NR1D1-Promoter-Luc熒光素酶報告質粒的構建
使用基因組DNA提取試劑盒提取山羊肝組織的基因組DNA。以基因組DNA為模板,利用設計好的帶有同源臂的引物(表1)分別擴增山羊1-Promoter片段以及1D1-Promoter片段,擴增產物用1%瓊脂糖凝膠進行凝膠電泳鑒定,目的條帶按照膠回收試劑盒操作說明分別進行膠回收。將膠回收產物分別與I或I線性化的pGL4.10載體進行同源重組連接,操作步驟同“1.3.2”。隨后分別使用限制性核酸內切酶I和I對連接有1-Promoter片段及連接有1D1-Promoter片段的重組質粒進行單酶切鑒定。同時以重組質粒為模板,使用1-Promoter和1D1-Promoter的擴增引物對相應的重組質粒進行PCR鑒定。將酶切及PCR鑒定結果符合預期的重組質粒送至西安擎科生物有限公司進行測序鑒定,返回序列與NCBI數據庫中已知序列進行比對。將測序正確的重組質粒分別命名為pGL4.10-BMAL1-Promoter-和pGL4.10-NR1D1-Promoter-。
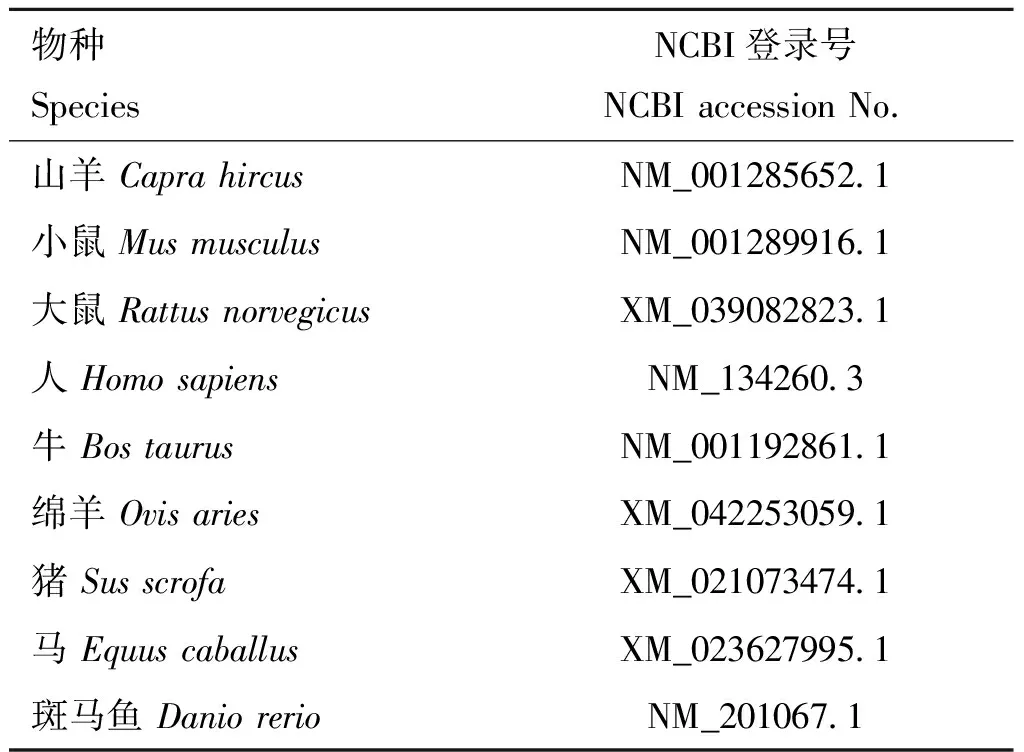
表2 各物種RORα基因CDS區序列來源
1.6 雙熒光素酶報告試驗
將HEK293T細胞以每孔5×10個細胞接種至1個24孔板中,置于細胞培養箱中培養24 h。隨后,使用TurboFect轉染試劑將pcDNA3.1-3HA-gRORα真核表達載體(試驗組)、pcDNA3.1-Puro-N-3HA空載體(對照組)分別與pRL-CMV海腎熒光素酶報告質粒(內參質粒)和pGL4.10-BMAL1-Promoter-(或pGL4.10-NR1D1-Promoter-)螢火蟲熒光素酶報告質粒共轉染至HEK293T細胞中,每組平行設3個重復孔。轉染36 h后,按照雙熒光素酶報告試驗試劑盒說明收集并處理細胞樣品,使用BioTek Synergy 2多功能酶標儀測定熒光值,計算相對熒光強度,測定重復3次取平均值。
1.7 數據統計分析
使用GraphPad Prism 6.0軟件進行統計學分析,結果以“平均值±標準誤”表示,兩組間樣本均數比較采取檢驗,*<0.05表示差異顯著,**<0.01表示差異極顯著。
2 結 果
2.1 山羊RORα基因的克隆及其真核表達載體的構建
對山羊α基因CDS區全長序列分析后發現,山羊α基因共包含10個外顯子,各個外顯子長度在28 ~ 396 bp之間,所有內含子的剪接位點序列均符合GT-AG法則(表3)。隨后,提取了山羊肝組織的總RNA,反轉錄得到cDNA,以cDNA為模板使用PCR技術成功擴增獲得山羊α基因CDS區片段,電泳結果如圖1A所示,在1 500 bp下方觀察到單一條帶,與預期結果一致(1 407 bp)。將擴增片段進行純化回收后,與R I酶切線性化的骨架載體pcDNA3.1-Puro-N-3HA進行同源重組連接,構建真核表達載體pcDNA3.1-3HA-gRORα。通過酶切和PCR鑒定重組載體,同時用R I對空載體進行單酶切作對照,鑒定結果如圖1B所示。重組載體經過R I酶切后獲得兩條目的條帶,上方條帶大小與空載體單酶切條帶一致(5 211 bp), 下方條帶大小與PCR擴增產物條帶大小一致(1 407 bp)。最后將鑒定正確的質粒進行測序,測序結果顯示pcDNA3.1-3HA-gRORα中插入片段的序列與NCBI庫中山羊α基因已知序列完全一致。以上結果表明成功獲得了山羊α基因CDS區全長片段,且真核表達載體pcDNA3.1-3HA-gRORα構建成功。
2.2 qPCR和WB檢測山羊RORα基因在mRNA及蛋白水平的過表達效果
qPCR結果如圖2A所示,與對照組相比,試驗組α基因的mRNA表達量升高約500倍(<0.01)。WB結果如圖2B所示,用HA抗體孵育后,試驗組在56 ku處有明顯的條帶,而對照組在相應位置無條帶。上述結果表明,pcDNA3.1-3HA-gRORα轉染HEK293T細胞后山羊α基因在mRNA和蛋白表達水平均明顯高于對照組。
2.3 山羊RORα基因的生物信息學分析
2.3.1 同源性比對及系統進化樹構建 使用DNAStar軟件對多個物種α基因CDS區進行相似性分析,結果如圖3所示。山羊α CDS區與小鼠、人、大鼠、牛、綿羊、馬、豬和斑馬魚的α CDS區相似性分別為:89.2%、94.1%、89.5%、97.1%、97.5%、94.0%、95.2%、70.7%。不同物種間基因相似性較高,進化過程中具有高度保守性。使用MEGA6軟件構建α基因的系統進化樹,結果如圖4所示,山羊與綿羊的親緣關系最近,其次是和牛、豬等其它哺乳動物,和斑馬魚親緣關系最遠。利用STRING數據庫檢索到RORB、RORC、NRIP1(nuclear receptor interacting protein 1)、SLC1A6(solute carrier family 1 member 6)、KAT5(lysine acetyltransferase 5)、GRM1(glutamate metabotropic receptor 1)、TRPC3(transient receptor potential cation channel subfamily C member 3)、ITPR1(inositol 1,4,5-trisphosphate receptor type 1)、ATP2A2(ATPase sarcoplasmic/endoplasmic reticulum Catransporting 2)、ATP2A3(ATPase sarcoplasmic/endoplasmic reticulum Catransporting 3)共10個蛋白可能和RORα蛋白存在相互作用(圖5)。
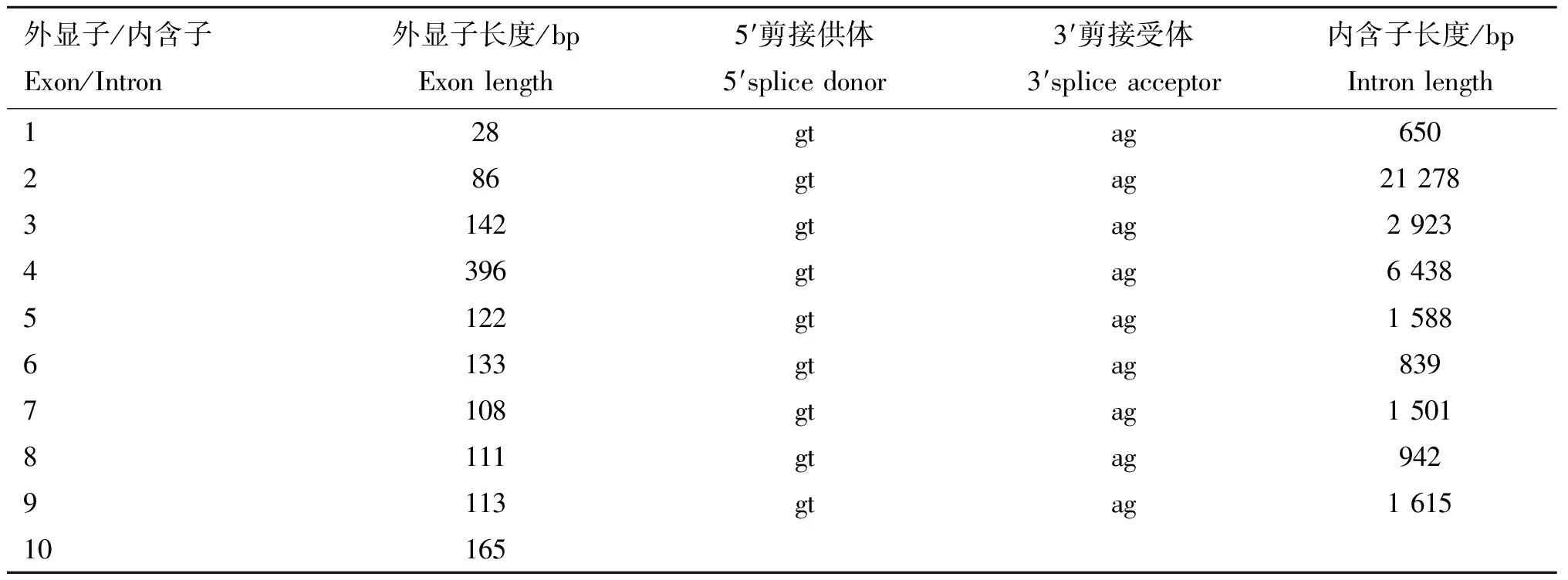
表3 山羊RORα基因的剪接位點
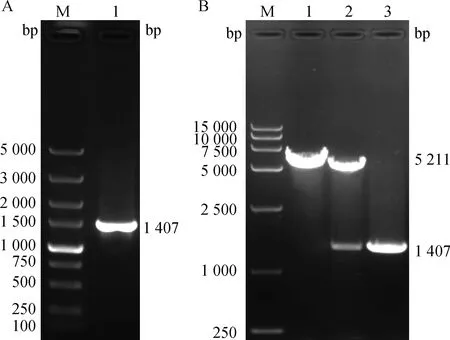
A. RORα基因CDS區擴增結果:M. DNA相對分子質量標準;1. RORα基因CDS區片段。B. pcDNA3.1-3HA-gRORα重組載體酶切鑒定結果:M. DNA相對分子質量標準;1. 空載體單酶切產物;2. 重組載體pcDNA3.1-3HA-gRORα單酶切產物;3. RORα基因質粒PCR目的條帶A. Amplification result of RORα gene CDS region: M. 5 000 bp DNA Marker; 1. The fragment of RORα gene CDS region. B. Restriction digestion result of pcDNA3.1-3HA-gRORα eukaryotic expression vector: M. 15 000 bp DNA Marker; 1. Single digested product of empty vector; 2. Single digestion product of recombinant vector pcDNA3.1-3HA-gRORα; 3. The PCR target band of RORα gene圖1 山羊RORα基因CDS擴增與pcDNA3.1-3HA-gRORα真核表達載體鑒定結果Fig.1 Results of goat RORα gene CDS amplification and identification of pcDNA3.1-3HA-gRORα eukaryotic expression vector

A.山羊RORα基因mRNA水平相對表達量;B. RORα蛋白WB鑒定結果。**表示差異極顯著(P<0.01)A. Relative mRNA expression of goat RORα gene; B. WB analysis of goat RORα protein expression. ** indicating that the difference is extremely significant (P< 0.01)圖2 qPCR及WB檢測結果Fig.2 Results of qPCR and WB analysis

圖3 RORα基因CDS區序列同源性比對Fig.3 Sequence homology alignment of RORα gene CDS region
2.3.2 山羊RORα蛋白理化性質分析 利用在線工具ExPASy中的ProtParam對山羊RORα蛋白進行理化性質分析,結果顯示山羊RORα蛋白理論分子質量為53 397.15,分子式為CHNOS,原子總數是7 430個,等電點(pI)為6.37,屬于堿性蛋白;該蛋白含有468個氨基酸,其中亮氨酸含量最高(8.3%),色氨酸含量最低(0.6%)(表4);在水中以280 nm預測消光系數是41 590;半衰期為30 h;不穩定系數為38.97;脂肪族系數為78.16。
2.3.3 親/疏水性預測 利用在線工具ExPASy對山羊RORα蛋白氨基酸序列的親水/疏水性進行預測分析,結果如圖6所示,山羊RORα蛋白氨基酸序列最低分值為-3.578(位于第115、116位氨基酸),親水性最強;最高分值為2.744(位于第369位氨基酸),疏水性最強。綜合來看,該蛋白整條肽鏈大部分屬于親水區域,由此推測山羊RORα蛋白為親水性蛋白。
2.3.4 信號肽及跨膜區預測 利用在線工具SignalP-5.0對山羊RORα氨基酸進行信號肽預測,結果顯示,該蛋白有12%的概率存在位于0~18氨基酸之間的信號肽,該蛋白有信號肽的概率較小。利用在線軟件TMHMM Serverv.2.0對山羊RORα蛋白進行跨膜結構預測,結果顯示,該蛋白不存在跨膜區。
2.3.5 山羊RORα蛋白二級結構預測 利用在線工具SOPMA對山羊RORα蛋白二級結構進行預測,結果如圖7所示,山羊RORα蛋白主要由α-螺 旋、延伸鏈、β-轉角和無規則卷曲等組成,其中α-螺旋占43.8%,延伸鏈占10.04%,β-轉角占4.91%,無規則卷曲占41.24%。
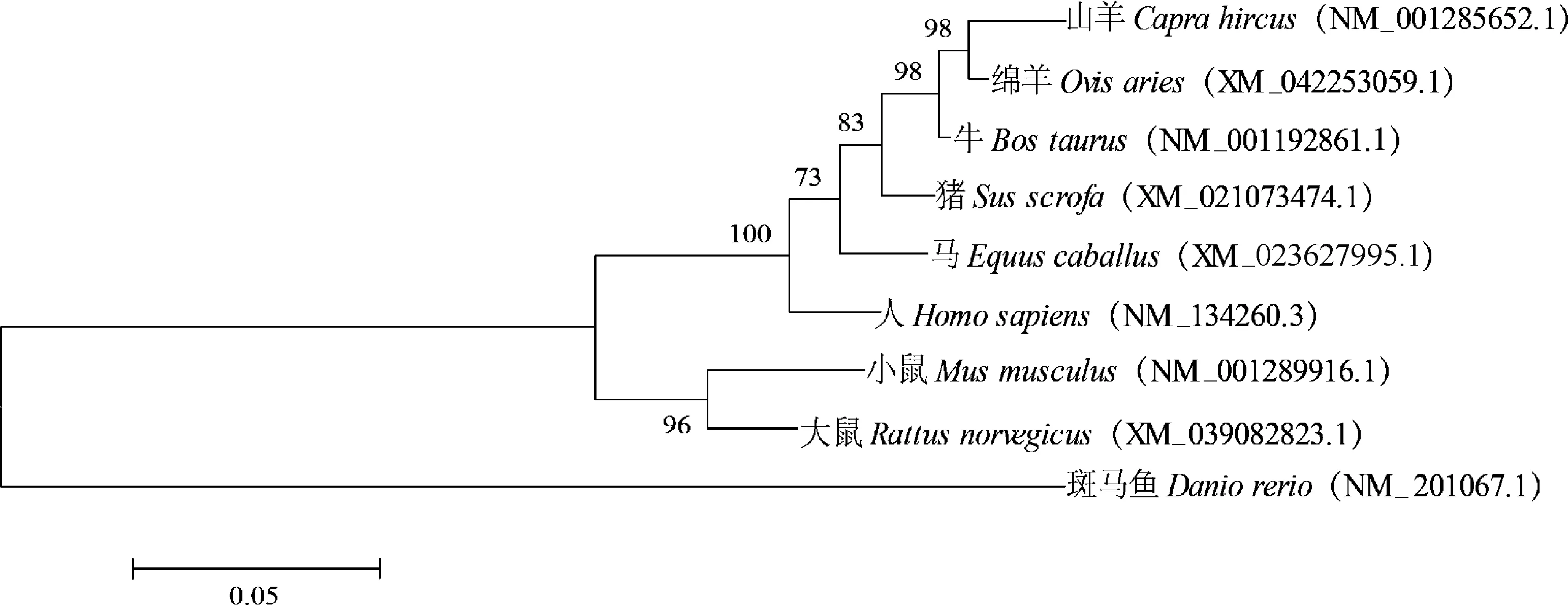
圖4 RORα基因的系統進化樹Fig.4 Phylogenetic tree of RORα gene

圖5 RORα互作蛋白預測Fig.5 Prediction of proteins interacting with RORα protein

表4 山羊RORα蛋白氨基酸組成
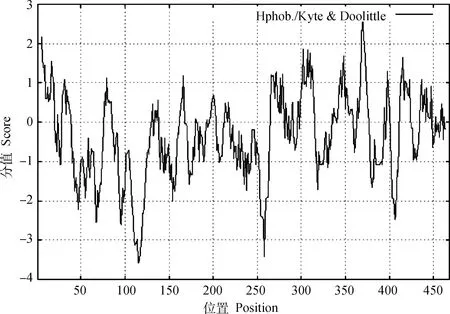
圖6 山羊RORα蛋白親/疏水性預測Fig.6 Hydrophobicity/hydrophilicity prediction of goat RORα protein

h表示α-螺旋,e表示延伸鏈,t表示β-轉角,c表示無規則卷曲h stands for alpha helix, e stands for extended strand, t stands for beta turn, c stands for random coil圖7 山羊RORα蛋白二級結構分析Fig.7 Analysis of the goat RORα protein secondary structure
2.3.6 山羊RORα蛋白三級結構預測分析 通過在線軟件SWISS-MODEL對山羊(圖8A)、小鼠(圖8B)和人(圖8C)的RORα蛋白三級結構進行預測,結果顯示,3個物種的RORα蛋白均具有α-螺旋、β-轉角和無規則卷曲等復雜結構。使用pyMOL軟件對3個物種的蛋白模型的結構進行比對,結果顯示山羊和小鼠模型的原子位置均方根偏差(root-mean-square deviation,RMSD)為0.068,山羊和人的RMSD為0.077,山羊RORα蛋白三級結構模型與小鼠和人的差異均極小。
2.4 pGL4.10-BMAL1-Promoter-Luc及pGL4.10-NR1D1-Promoter-Luc載體的鑒定
以山羊肝基因組DNA為模板,利用合成的1-Promoter及1D1-Promoter引物進行PCR擴增。結果如圖9A、9C所示,1-Promoter(862 bp)及1D1-Promoter(1 657 bp)PCR擴增產物條帶單一,與目的片段大小一致,無非特異性條帶。將克隆得到的目的條帶分別與pGL4.10空載體進行重組連接,構建pGL4.10-BMAL1-Pro-moter-和pGL4.10-NR1D1-Promoter-重組質粒。對重組質粒分別進行PCR及酶切鑒定,結果顯示pGL4.10-BMAL1-Promoter-報告質粒經I單酶切后得到862 和4 242 bp兩個片段(圖9B),且以pGL4.10-BMAL1-Promoter-為模板PCR擴增出大小約862 bp的片段。pGL4.10-NR1D1-Promoter-報告質粒經I單酶切后得到1 657和4 242 bp兩個片段(圖9D),且以pGL4.10- NR1D1-Promoter-為模板PCR擴增出大小約1 657 bp 的片段。同時,兩個重組質粒的測序結果也均與NCBI數據庫中已知序列完全一致。上述結果表明,含有山羊1基因啟動子片段的pGL4.10-BMAL1-Promoter-重組質粒和含有山羊1D1基因啟動子片段的pGL4.10-NR1D1-Promoter-重組質粒構建成功。

A.山羊RORα蛋白三級結構預測;B.小鼠RORα蛋白三級結構預測;C.人RORα蛋白三級結構預測A. Prediction of goat RORα protein tertiary structure; B. Prediction of mouse RORα protein tertiary structure; C. Prediction of human RORα protein tertiary structure圖8 山羊、小鼠和人RORα蛋白的三級結構預測Fig.8 Prediction of the tertiary structure of RORα protein in goat, mouse and human
2.5 山羊RORα蛋白成功上調山羊BMAL1和NR1D1啟動子的轉錄活性
使用pcDNA3.1-3HA-gRORα質粒及含有RORE元件的pGL4.10-BMAL1-Promoter-和pGL4.10-NR1D1-Promoter-重組質粒分別進行雙熒光素酶報告試驗以探究山羊RORα蛋白的功能。如圖10A所示,本研究中擴增獲得的1-Promoter片段位于山羊1基因的-684 ~ +178位置(山羊1基因的轉錄起始位點為+1),包含3個RORE元件;擴增獲得的1D1-Promoter位于山羊1D1基因的-1 421~+236位置(山羊1D1基因的轉錄起始位點為+1), 也包含3個RORE元件。雙熒光素酶報告試驗結果表明,與共轉染pcDNA3.1-Puro-N-3HA空質粒的對照組相比,共轉染pcDNA3.1-3HA-gRORα質粒時1-Promoter(圖10B)和1D1-Promoter(圖10C)調控下的熒光素酶活性均極顯著升高(<0.01)。上述結果表明,山羊RORα蛋白可以顯著上調山羊生物鐘基因1和1D1啟動子的轉錄活性,這與小鼠等哺乳動物生物鐘轉錄調控機制相符。
3 討 論
晝夜節律是地球上的生命體對地球自轉產生的晝夜更替現象所做出的主動適應性變化。多項研究表明,晝夜節律紊亂與睡眠障礙、代謝失調、腫瘤等多種疾病的發生密切相關。生物體內參與生物鐘發生和維持的基因被稱為生物鐘基因,這些基因與鐘控基因共同調控哺乳動物生理、生化、行為等方面的晝夜節律性變化。RORα作為生物鐘系統的重要組分,在維持生物鐘的正常運轉過程中發揮重要作用。然而,目前有關生物鐘分子調控機制的研究大多是以小鼠為模型,關于山羊生物鐘基因及山羊生物鐘轉錄調控機制方面的研究相對較少。本課題組前期成功構建了山羊生物鐘基因1、和1D1的真核表達載體,發現1與1D1基因mRNA節律性表達于山羊睪丸間質細胞中,并證明了1基因通過調節山羊睪丸間質細胞中類固醇合成相關基因的轉錄來調控睪酮產生。趙艷紅等前期雖已克隆了內蒙古絨山羊α基因的CDS區,但只是部分序列(417 bp),目前尚未有克隆山羊α基因全長CDS序列(1 407 bp)的研究報道。基于此,本研究克隆了山羊α基因CDS區全長片段,構建了其真核表達載體,并對山羊RORα進行了生物信息學分析,初步探究了其在山羊生物鐘系統中的作用機制,為后續進一步深入解析山羊生物鐘系統的轉錄調控機制奠定基礎。

A. BMAL1-Promoter PCR擴增結果:M. DNA相對分子質量標準;1. BMAL1-Promoter目的片段。B. pGL4.10-BMAL1-Promoter-Luc載體酶切及PCR鑒定結果:M. DNA相對分子質量標準;1. 空載體單酶切產物;2. pGL4.10-BMAL1-Promoter-Luc載體單酶切產物;3. BMAL1-Promoter質粒PCR目的條帶。C. NR1D1-Promoter PCR擴增結果:M.DNA相對分子質量標準;1.NR1D1-Promoter目的片段。D. pGL4.10-NR1D1-Promoter-Luc載體酶切及PCR鑒定結果:M.DNA相對分子質量標準;1. 空載體單酶切產物;2. pGL4.10-NR1D1-Promoter-Luc載體單酶切產物;3. NR1D1-Promoter質粒PCR目的條帶A. PCR amplification result of BMAL1-Promoter: M. 5 000 bp DNA Marker; 1. The target band of BMAL1-Promoter.B. Restriction digestion and PCR result of pGL4.10-BMAL1-Promoter-Luc vector: M. 5 000 bp DNA Marker; 1. Single digested product of empty vector; 2. Single digestion product of pGL4.10-BMAL1-Promoter-Luc vector; 3. The target band of BMAL1-Promoter. C. PCR amplification result of NR1D1-Promoter: M. 5 000 bp DNA Marker; 1. The target band of NR1D1-Promoter.D. Restriction digestion and PCR result of pGL4.10-NR1D1-Promoter-Luc vector: M. 5 000 bp DNA Marker; 1. Single digested product of empty vector; 2. Single digestion product of pGL4.10-NR1D1-Promoter-Luc vector; 3. The target band of NR1D1-Promoter圖9 pGL4.10-BMAL1-Promoter-Luc和pGL4.10-NR1D1-Promoter-Luc質粒鑒定結果Fig.9 Results of identification of pGL4.10-BMAL1-Promoter-Luc and pGL4.10-NR1D1-Promoter-Luc plasmids
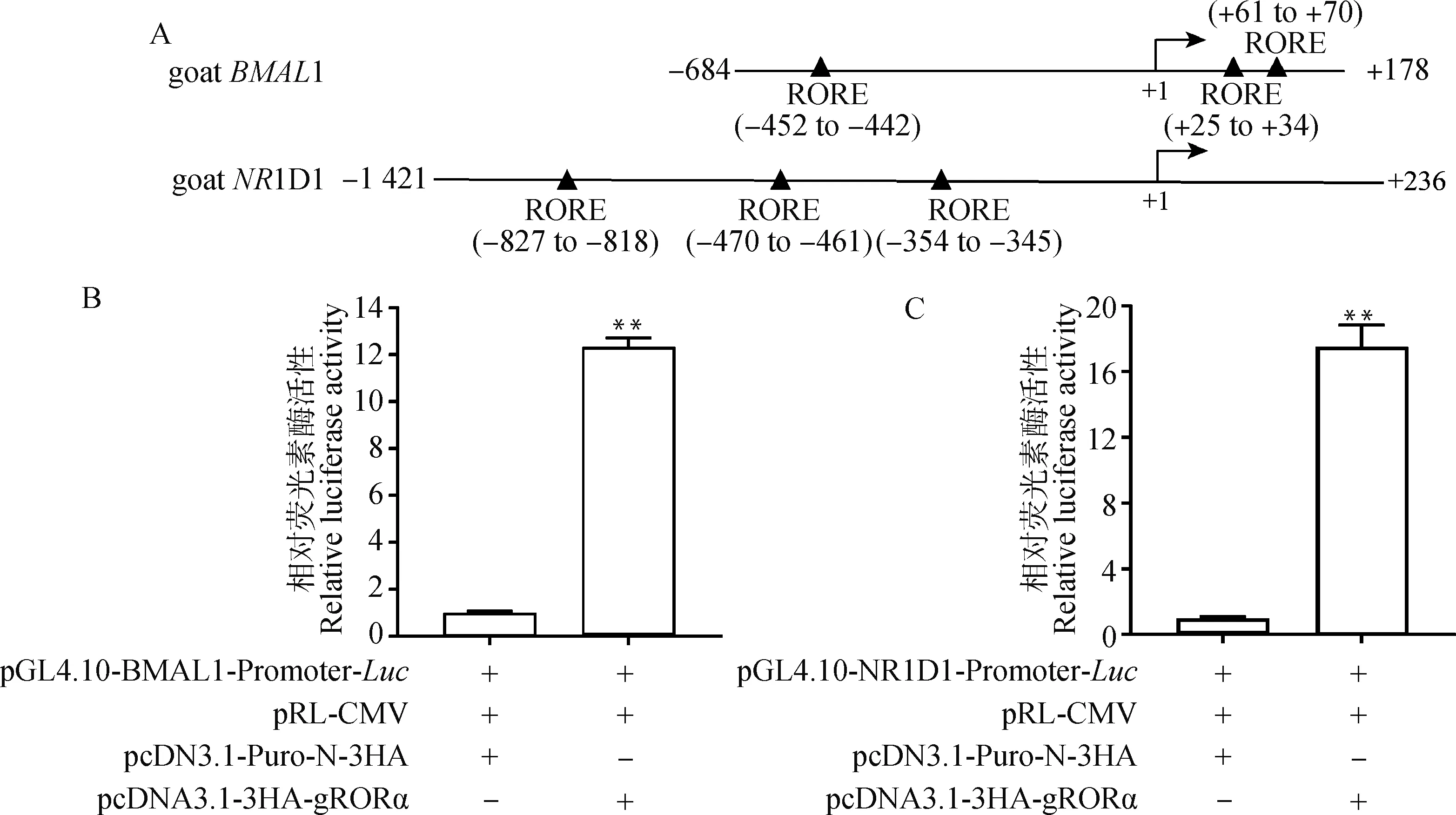
A.山羊BMAL1和NR1D1基因啟動子區域RORE元件相對于基因轉錄起始位點(+1)的位置示意圖;B、C. 雙熒光素酶報告試驗結果。**表示差異極顯著(P<0.01)A. Schematic diagram of the position of the RORE response element in the promoter region of goat BMAL1 and NR1D1 genes relative to the transcription start site (+1) of the gene; B, C. Dual-luciferase reporter assay results. ** means the difference is extremely significant (P<0.01)圖10 RORα蛋白對山羊BMAL1和NR1D1啟動子轉錄活性的影響Fig.10 Effects of RORα protein on the transcriptional activities of goat BMAL1 and NR1D1 promoters
自α基因在小鼠中被克隆鑒定以來,人們對其生理功能的認識不斷增加。Chen等發現,α基因在小鼠小腦發育中起著關鍵作用,α基因缺陷小鼠表現出共濟失調和嚴重的小腦萎縮,其特征是浦肯野細胞明顯減少,小腦顆粒細胞丟失。Dzhagalov等研究表明,RORα在胸腺生成和淋巴細胞發育過程中也發揮重要作用,α缺失小鼠的脾和胸腺都明顯較小,且成熟T和B淋巴細胞均顯著減少。此外,還有大量證據表明,RORα在視網膜、大腦以及T細胞等發育過程中有著重要的調控作用。同時,許多研究發現RORα可以參與調控小鼠和人等哺乳動物的生殖系統發育以及睪酮合成。Sayed等在α基因敲除小鼠睪丸中觀察到生精不足、睪丸間質細胞數量減少以及精子形態發生變化等現象。隨后,該研究又發現α基因突變的老年小鼠會出現生精小管萎縮以及精子頂體帽缺失和空泡化的現象。Yang等研究發現,褪黑素可通過RORα通路來提高雄性哺乳動物睪丸間質細胞的睪酮合成能力。另外,在綿羊的研究發現,褪黑素可通過RORα在體內和體外水平上提高青春期前卵丘細胞抗氧化能力。同時,在山羊精原干細胞體外分化培養系統中,褪黑素可通過RORα通路增加睪丸間質細胞的睪酮分泌。總之,上述研究結果表明RORα在哺乳動物的多個生物學過程中都扮演著重要角色。
哺乳動物的生物鐘系統由位于SCN的中樞生物鐘和分布于機體大部分組織器官的外周生物鐘組成。肝是機體中最為主要的外周生物鐘器官,其在機體營養代謝中發揮重要作用,由相對均質的細胞類型組成,且易于獲取。另外,生物鐘基因α作為一種重要的核受體,與肝脂質代謝調節密切相關,且在小鼠肝中高表達。據此推測,α基因也高表達于山羊肝中,所以本研究選取山羊肝組織來提取cDNA,進一步克隆α基因的CDS區全長序列。另外,本研究還對山羊α基因序列進行了生物信息學分析,同源性分析結果顯示山羊α基因的CDS區序列與綿羊和牛等晝行性反芻動物的α基因相似性極高,說明此基因在反芻動物具有較高的保守性和同源性。另外,分析系統進化樹結果發現,進化樹的形態與分類學基本保持一致。
此外,本研究結果還表明山羊RORα蛋白屬于一種親水性的非跨膜蛋白。通過將山羊、小鼠和人的RORα蛋白三維結構比對后發現,三者間存在高度的相似性,均含有4個主要功能區組成的核受體結構域:N-末端(A/B)結構域、高度保守的DNA結合域(DNA binding domain,DBD)、鉸鏈域和C-末端配體結合域。由于小鼠RORα蛋白的DBD已被證明可通過與特定的DNA反應元件(RORE元件)結合來調控基因轉錄,進而發揮其在生物鐘系統及各項生理過程中的調控作用。蛋白質結構決定其功能,所以推測山羊RORα蛋白功能與小鼠之間存在保守性,其可能也在山羊生物鐘系統中發揮重要調控作用。本研究使用了雙熒光素酶報告試驗探究驗證山羊RORα蛋白的功能,結果顯示山羊RORα蛋白可以顯著上調山羊1和1D1基因啟動子的轉錄活性。該結果進一步證實了上述猜想,證實山羊RORα蛋白確實在山羊生物鐘系統中發揮重要調控作用。另外,結合小鼠和人RORα在調控機體生理功能方面已有的研究證據推測,RORα可能在山羊的生長發育過程中也發揮重要作用,但具體功能有待于進一步的研究。
4 結 論
本研究克隆了山羊α基因的全長CDS區片段,構建了其真核表達載體,從mRNA水平和蛋白水平驗證了α在HEK293T細胞中的過表達效果,并對該基因進行了生物信息學分析。結果表明,RORα蛋白在山羊與小鼠等哺乳動物間存在高度保守性,其可以正向調控山羊1和1D1基因的啟動子活性,為進一步探究山羊核受體RORα的功能及山羊生物鐘的轉錄調控機制提供了前期基礎和關鍵材料。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
