lncRNA-6617調控豬肌內前體脂肪細胞分化的篩選與功能研究
2022-07-07 08:23:14李睿霄姬夢婷蔡春波高鵬飛郭曉紅曹果清李步高
畜牧獸醫學報 2022年6期
關鍵詞:研究
孟 珊,楊 陽,李睿霄,姬夢婷,張 娜,路 暢,蔡春波,高鵬飛,郭曉紅,曹果清,李步高
(山西農業大學動物科學學院,太谷 030801)
脂肪組織傳統上被認為是機體內主要的能量儲存器官。隨著分子研究層面的不斷深入,研究發現脂肪組織也是機體最大的內分泌器官之一,通過分泌大量的脂肪細胞因子影響全身新陳代謝。脂肪組織分布在動物機體內的多個部位,如皮下脂肪和內臟脂肪等,其中存在于肌纖維之間和肌纖維束之間的肌內脂肪,是決定動物肉品質的重要因素之一,影響著肉的口感和風味。研究顯示,肌內脂肪細胞來源于肌肉血管基質部分(sromal vascular fraction, SVF)中的肌內前體脂肪細胞,逐步分化形成成熟的脂肪細胞。近年來,隨著功能基因組學研究和轉錄組測序技術的應用,越來越多的調控因子被報道參與脂肪生成的調控。長鏈非編碼RNA(long noncoding RNA, lncRNA)是一類轉錄本長度大于200 nt的RNA分子,通過影響下游關鍵因子的表達在脂肪細胞生成過程中發揮著重要的調控作用。但關于lncRNA調控肌內脂肪生成的研究還較少,其調控網絡尚不夠完善。目前,關于lncRNA調控脂肪細胞生成的研究主要集中于下游的分子機制,lncRNA本身是如何被調控?其調控的分子機制還尚待研究。N6-甲基腺苷(N6-methyladenosine,mA)是一類在真核生物RNA上廣泛存在的表觀遺傳修飾,研究發現,lncRNA本身存在著mA修飾,mA修飾水平的變化影響著lncRNA 功能的發揮,從而參與細胞增殖、纖維化、癌癥等的調控。2014年,首次發現了去甲基化酶通過mA修飾的靶mRNA的選擇性剪切,影響脂肪生成。但是mA修飾是否可以通過介導lncRNA的作用參與成脂分化的調控還需進一步揭示。
馬身豬是山西省地方品種,表現出肉質優良、繁殖力較高、抗逆性強,且背最長肌中肌內脂肪含量極顯著高于大白豬。研究發現,在馬身豬肌肉組織中,脂肪細胞生成的關鍵調控因子423、和表達量顯著升高,說明馬身豬的肌內脂肪細胞生成相較于大白豬顯著增加。因此,本研究通過對肌內脂肪含量高的馬身豬和肌內脂肪含量較低的大白豬背最長肌組織進行lncRNA測序,從而篩選到一個在馬身豬中特異高表達的lncRNA-6617。通過生物信息學軟件分析其生物學特性,比較分析lncRNA-6617在馬身豬和大白豬肌肉組織間的表達差異。利用體外分離培養的豬肌內前體脂肪細胞為模型,干擾lncRNA-6617,探究其對豬肌內前體脂肪細胞成脂分化的影響。為了尋找lncRNA-6617上游作用機制的調控方式,檢測馬身豬和大白豬肌肉組織中mA修飾相關酶的表達差異,為后續分子作用機制的深入研究提供理論基礎。
1 材料與方法
1.1 試驗材料
選取飼養在相同條件下的1、90、180日齡健康馬身公豬和大白公豬各3頭,分別屠宰采集其心、肝、脾、肺、背部和腹部皮下脂肪、背最長肌、腰大肌、股二頭肌等組織,在液氮中速凍,隨后于-80 ℃冰箱保存備用。
1.2 主要試劑與儀器
1.2.1 主要試劑 胎牛血清和Opti-MEM購自美國Gibco公司;胰酶、青鏈霉素、高糖DMEM和胰島素購自美國Sigma公司;Dex、IBMX、4%多聚甲醛和飽和油紅O染液購自北京Solarbio公司;吲哚美辛購自阿拉丁;Trizol Reagen和PARISKit購自美國Life Technologies公司;Prime Script RT reagent Kit with gDNA Eraser和SYBR Premix Ex Taq II等購自北京全式金;Lipofectamine 2000 Reagent購自Invitrogen公司;引物由上海生工合成;siRNA委托上海吉瑪公司設計并合成。
1.2.2 主要儀器 細胞成像系統(EVOS FL Auto,Life Technologies,美國),Nanodrop 2000核酸蛋白測定儀(Thermo,美國),普通PCR儀(Bio-RAD,美國),實時熒光定量PCR儀(Bio-RAD,美國)。
1.3 試驗方法
1.3.1 RNA提取及cDNA文庫的構建和測序 按照Trizol說明書方法提取1、90和180日齡馬身豬和大白豬背最長肌組織RNA,經質量檢測合格后,進行Illumina Hiseq上機測序。
1.3.2 豬lncRNA-6617的篩選及生物信息學分析 通過edgeR和cuffdiff軟件對1日齡馬身豬與大白豬背最長肌lncRNA測序結果分析,選定馬身豬中特異高表達的lncRNA-6617為目標lncRNA。 利用UCSC-Blast和Ensemble-Blast明確lncRNA-6617的基因組定位;通過CPC、CPAT在線網站預測lncRNA-6617的蛋白編碼能力。
1.3.3 lncRNA-6617的鑒定 根據測序得到的lncRNA-6617序列信息,用Primer Premier 6.0軟件進行引物設計,以馬身豬背最長肌組織cDNA為模板,通過RT-PCR對lncRNA進行擴增,隨后用1%瓊脂糖凝膠電泳檢測擴增產物,對目的片段進行膠回收,連接轉化,挑取陽性單菌落進行PCR鑒定,將篩選出的陽性菌送至華大基因測序。
1.3.4 豬lncRNA-6617的核質定位 試驗選用未分化的豬肌內前體脂肪細胞,按照試劑盒說明書進行細胞核和細胞質RNA分離,把RNA反轉錄成cDNA,通過qRT-PCR檢測lncRNA-6617在豬肌內前體脂肪細胞的細胞核和細胞質中的表達量。
1.3.5 豬肌內前體脂肪細胞的成脂分化 馬身豬肌內前體脂肪細胞由實驗室前期分離培養和保存,將傳代細胞接種于6孔板,采用“激素雞尾酒”法誘導成脂。待細胞完全匯合后,更換為誘導培養基(完全培養基+10 μg·mLIns+0.5 mmoL·LIBMX+1 μmoL·LDEX+100 μmoL·LIND),4 d后更換為維持培養基(完全培養基+10 μg·mLIns),每2 d換1次培養基,直至脂滴融合成大脂滴。
1.3.6 細胞轉染及油紅O染色 將肌內前體脂肪細胞接種于6孔板,密度達到60%左右時進行轉染。轉染前更換為新鮮培養基,siRNA試劑按照Lipofectamine 2000說明書步驟轉染,每組設置3個重復孔。轉染48 h后收集細胞,檢測干擾效率。同時收集轉染后誘導分化7 d的細胞,檢測分化中后期成脂關鍵基因的表達。待細胞脂滴明顯形成后,用預冷的PBS清洗兩遍,4%多聚甲醛固定30 min,棄去甲醛,60%異丙醇浸洗1 min,棄去異丙醇。待細胞完全干燥,油紅O染液(飽和油紅O∶蒸餾水=3∶2)染色10 min,PBS洗去多余的油紅O染液,顯微鏡下觀察拍照。
1.3.7 總RNA的提取與實時定量PCR 按照Trizol說明書方法提取RNA。按照全式金反轉錄試劑盒說明書合成第一鏈cDNA,稀釋為20倍。然后以該cDNA作為模板,通過qRT-PCR檢測基因表達情況,-為內參基因,引物序列見表1。反應體系為10 μL:SYBR為5 μL,cDNA為4.4 μL,上、下游引物各為0.3 μL。反應程序:95 ℃預變性3 min;95 ℃變性10 s,60 ℃退火20 s,72 ℃延伸30 s, 40個循環。基因相對表達量根據2法計算。

表1 引物信息
1.3.8 數據統計與分析 試驗均設置3個生物學重復,所有數據結果使用SPSS Statistics 22.0軟件one-way ANOVA和獨立樣本檢驗進行顯著性分析,采用Duncan′s法進行多重比較,<0.05表示差異顯著,<0.01表示差異極顯著。
2 結 果
2.1 lncRNA-6617的篩選
利用edgeR和cuffdiff軟件分析不同發育階段馬身豬和大白豬背最長肌組織中差異表達的lncRNA,結果顯示,1日齡馬身豬與90日齡馬身豬、1日齡馬身豬與180日齡馬身豬、90日齡馬身豬與180日齡馬身豬相比,分別有88、60、18個差異表達的lncRNAs (圖1A、1B),1日齡馬身豬與1日齡大白豬、90日齡馬身豬與90日齡大白豬、180日齡馬身豬與180日齡大白豬相比,分別有87、21、16個差異表達的lncRNAs(圖1C、1D)。
通過比較分析1日齡馬身豬與大白豬背最長肌差異lncRNA的表達量,篩選到在馬身豬中特異高表達的lncRNA-6617(圖2)。
2.2 lncRNA-6617的生物學鑒定
利用測序得到的lncRNA-6617序列放入UCSC和Ensemble數據庫進行比對,發現lncRNA-6617位于豬第18號染色體上,由3基因所在DNA鏈的反義鏈轉錄而來,與3基因內含子3具有278 bp的堿基互補區域(圖3A),提示該lncRNA 為反義型lncRNA。
通過CPAT及CPC在線網站對lncRNA-6617進行蛋白編碼能力預測分析,選擇已報道的非編碼基因1為參考,結果表明,lncRNA-6617序列不具有編碼蛋白質的能力(圖3B)。利用未分化的豬肌內前體脂肪細胞進行核質分離試驗,確定lncRNA-6617 的亞細胞定位,結果顯示,lncRNA-6617主要分布于細胞核(圖3C)。
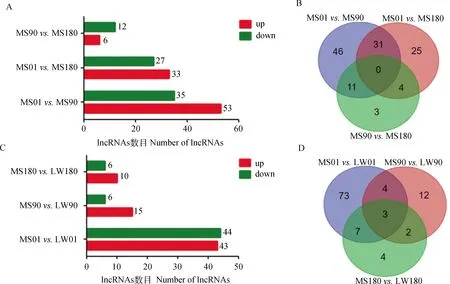
MS. 馬身豬;LW. 大白豬。下同MS. Mashen pig; LW. Large White pig. The same as below圖1 馬身豬和大白豬不同階段差異表達的lncRNA數量Fig.1 The number of differentially expressed lncRNAs at different stages of MS and LW pigs

圖2 1日齡馬身豬和大白豬差異lncRNA熱圖(A)和火山圖(B)Fig.2 The difference lncRNAs heat map (A) and volcano map (B) of 1-day-old MS and LW pigs
為驗證序列的存在,以轉錄組測序獲得的序列為模板設計引物,通過RT-PCR擴增lncRNA-6617。瓊脂糖凝膠電泳結果顯示,擴增出長278 bp的目的條帶(圖3D),條帶清晰且單一,Sanger測序結果表明條帶序列比對一致(圖3E)。

A. 基因組位置;B. 蛋白編碼能力預測:a為CPAT結果,b為CPC結果;C. 亞細胞定位;D. PCR擴增;E. 測序結果與已知序列比對A. Genomic location; B. Protein coding ability prediction: a is the CPAT result, b is the CPC result. C. Subcellular localization; D. PCR amplification; E. Alignment of sequencing results with known sequences圖3 lncRNA-6617的生物學特性Fig.3 Biological characteristics of lncRNA-6617
2.3 lncRNA-6617的表達模式分析
為明確lncRNA-6617組織表達特性,通過qRT-PCR檢測了lncRNA-6617在馬身豬心、肝、脾、肺、背部和腹部皮下脂肪、背最長肌、腰大肌、股二頭肌等9個組織中的表達情況。結果顯示,lncRNA-6617 在各組織中均有表達(圖4A),背部皮下脂肪中表達量最高,其次是腹部皮下脂肪等組織。進一步發現,lncRNA-6617的表達量在肌內脂肪含量高的馬身豬肌肉組織中極顯著高于肌內脂肪含量較低的大白豬(<0.01,圖4B)。
對豬肌內前體脂肪細胞進行成脂誘導分化,分別收集誘導0、1、3、5、7 d的細胞,qRT-PCR檢測和lncRNA-6617在成脂誘導分化中的表達情況。結果表明,隨著成脂分化進程的進行,lncRNA-6617 表達量持續升高,與成脂核心調控因子的趨勢一致(圖4C、4D),相較于未分化階段,該lncRNA主要集中在分化中后期表達。
2.4 干擾lncRNA-6617抑制豬肌內前體脂肪細胞成脂分化
將siRNA和對照組silnc-NC分別轉染至豬肌內前體脂肪細胞中,48 h后收集細胞檢測干擾效率。結果顯示,與silnc-NC組相比,silnc-6617組極顯著降低了lncRNA-6617的表達(<0.01,圖5A)。干擾lncRNA-6617后,對豬肌內前體脂肪細胞進行誘導分化7 d后,分化的成熟脂肪細胞明顯減少(圖5C),成脂關鍵基因、和2的表達均極顯著下調(<0.01,圖5B),表明干擾lncRNA-6617抑制豬肌內前體脂肪細胞分化。
2.5 m6A去甲基化酶FTO調控lncRNA-6617的表達
為了探究lncRNA-6617上游的調控機制,本研究分析了馬身豬和大白豬肌肉組織mA修飾水平的差異,結果顯示,mA去甲基化酶的表達水平在馬身豬背最長肌和股二頭肌中極顯著高于大白豬(<0.01),在馬身豬腰大肌中顯著高于大白豬(<0.05), 上游調節因子217的表達水平在馬身豬各個肌肉組織中均極顯著高于大白豬(<0.01,圖6)。
基于SRAMP數據庫,利用lncRNA-6617序列預測mA修飾位點,發現在lncRNA-6617上存在mA潛在的修飾位點GGACU/A(圖7A)。在豬肌內前體脂肪細胞中干擾,發現干擾極顯著降低了lncRNA-6617的表達(<0.01,圖7B),說明mA修飾可能介導lncRNA-6617的表達。
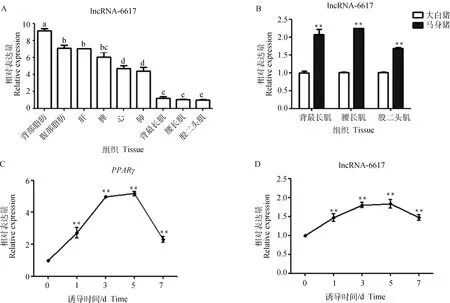
A. 組織表達譜,不同小寫字母表示差異顯著(P<0.05);B. 馬身豬和大白豬肌肉組織間的比較;C、D. 誘導肌內前體脂肪細胞分化過程中PPARγ和lncRNA-6617的時序表達,**. P<0.01,*. P<0.05,下同A. Tissue expression profile, different lowercase letters indicate significant difference(P<0.05); B. Comparison of muscle tissues between Mashen pigs and large White pigs; C, D. Temporal expression during induction of intramuscular preadipocytes differentiation, **. P<0.01, *. P<0.05, the same as below圖4 lncRNA-6617表達模式Fig.4 lncRNA-6617 expression patterns
3 討 論
lncRNA是一類轉錄本長度大于200 nt的RNA分子,通常不編碼蛋白質,而是以RNA的形式在表觀遺傳修飾、轉錄調控以及轉錄后修飾等生物學事件中參與蛋白編碼基因調控。隨著高通量測序的發展,一系列調控脂肪細胞生成的lncRNA 相繼被鑒定,其發揮作用的機制也不斷被揭示。lncRNA通過多種作用方式影響關鍵轉錄因子和等調控脂肪細胞生成。最近研究也發現,lncIMF4和IMFlnc1參與調控豬肌內脂肪細胞的生成。但是,目前在豬肌內脂肪細胞生成的研究中,更多側重于測序數據的分析,對于lncRNA的調控成脂分化功能研究尚不深入。
由于lncRNA具有明顯的時空表達特異性,使得研究肌內脂肪細胞生成需要進一步篩選鑒定特異表達的lncRNA。此前有研究者分別對絨山羊和肉用山羊肌內脂肪細胞成脂分化前后的細胞進行高通量測序,篩選與肌內沉積相關的功能候選基因。李嬡等利用RNA-Seq技術對大白豬和萊蕪豬肌內脂肪中circRNA進行鑒定,以篩選出調控豬脂肪沉積的circRNA,證明高通量測序是篩選功能基因的重要手段。本研究利用本課題組對肌內脂肪含量高的馬身豬和肌內脂肪含量較低的大白豬背最長肌組織lncRNA測序結果,篩選到在馬身豬中特異高表達的lncRNA-6617。進一步通過生物信息學分析和功能鑒定,發現lncRNA-6617在馬身豬肌肉組織中顯著高表達,說明lncRNA-6617可能是影響肌內脂肪沉積差異的一個關鍵因子。同時發現lncRNA-6617在豬肌內前體脂肪細胞成脂分化早期表達量逐漸上升,干擾lncRNA-6617后,顯著抑制肌內脂肪細胞的生成,說明lncRNA-6617是調控豬肌內脂肪細胞生成的關鍵因子。lncRNA-6617功能的發現可以進一步在理論上完善肌內脂肪細胞生成的調控網絡,為解析肌內脂肪組織發育機制和遺傳選育篩選靶標的確立提供理論依據。
作為遺傳信息的傳遞載體,RNA上的修飾對其功能的發揮起著至關重要的作用。前期已有研究對金華豬、長白豬及長金豬的脂肪組織與mRNA mA修飾水平進行相關性分析,發現mA修飾酶在肉脂型金華豬脂肪組織中表達量最高。同樣有研究發現,肌內脂肪含量高的金華豬背最長肌中mA修飾水平顯著低于含量較低的長白豬。本研究中,的表達在肌內脂肪含量高的馬身豬肌肉組織中顯著高于肌內脂肪含量較低的大白豬,且與lncRNA-6617的組織表達模式一致,提示馬身豬和大白豬肌內脂肪含量的差異可能與mA修飾相關。研究發現,在豬肌內前體脂肪細胞中,過表達甲基轉移酶3顯著抑制了脂肪細胞的生成。mA修飾作為RNA表觀修飾方式之一,同樣影響著lncRNA功能的發揮。于是本研究在肌內前體脂肪細胞中干擾,發現lncRNA-6617 的表達顯著下調,說明mA修飾的去甲基化酶可能介導lncRNA-6617功能的發揮,但其具體作用機制還需進一步研究。
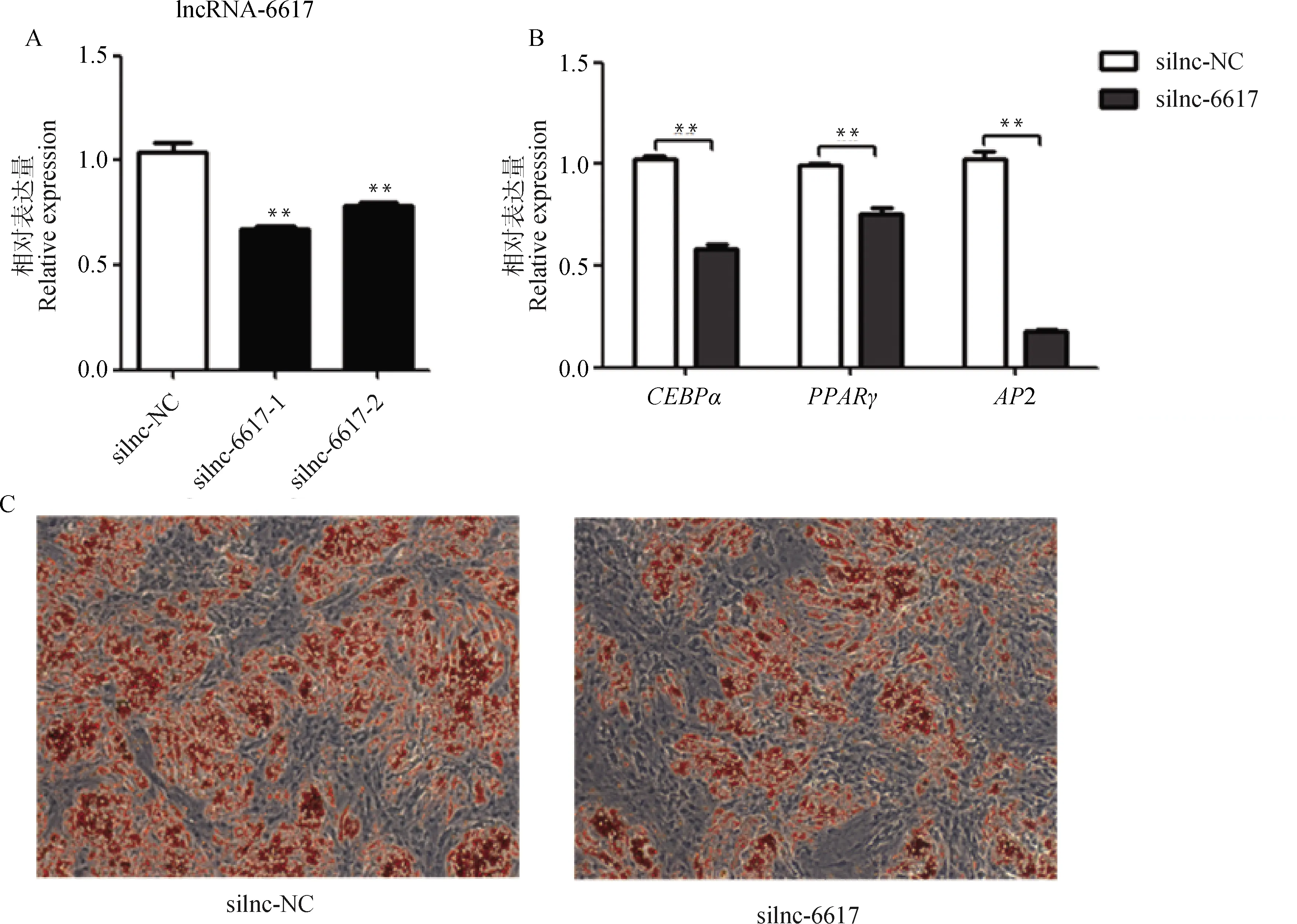
A. 干擾效率;B. 干擾lncRNA-6617后分化7 d成脂相關基因的表達變化;C. 油紅O染色(100×)A. Interference efficiency; B. The expression changes of adipogenesis-related genes after interfering with lncRNA-6617 on day 7 of diferentiation; C. Oil Red O staining (100×)圖5 干擾lncRNA-6617抑制豬肌內前體脂肪細胞成脂分化Fig.5 Interfering with lncRNA-6617 inhibits adipogenic differentiation of pig intramuscular preadipocytes

圖6 馬身豬和大白豬肌肉組織中m6A修飾相關酶的表達Fig.6 The expression of m6A modification related enzymes in muscle tissues of MS and LW pigs
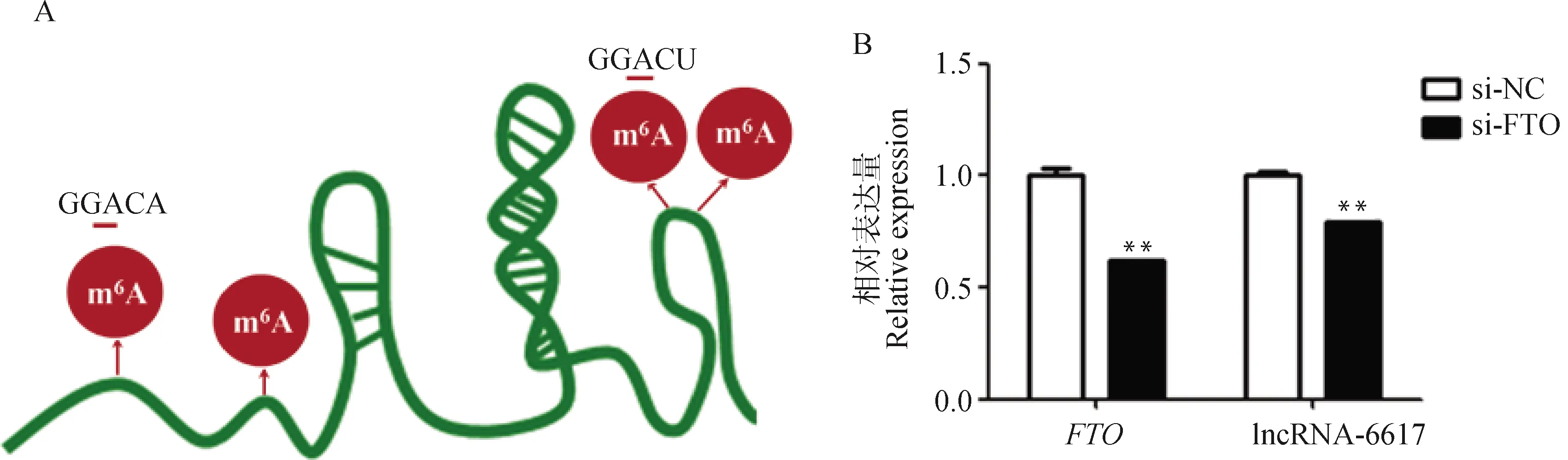
A. lncRNA-6617上m6A修飾位點預測;B. FTO干擾效率以及干擾FTO后lncRNA-6617的表達變化A. Prediction of m6A modification site on lncRNA-6617; B. The expression changes of FTO and lncRNA-6617 after interfering with FTO圖7 FTO可能調控lncRNA-6617的表達Fig.7 FTO may regulate the expression of lncRNA-6617
4 結 論
本研究鑒定了在馬身豬肌肉組織中特異高表達的lncRNA-6617;干擾lncRNA-6617后,顯著抑制豬肌內前體脂肪細胞分化;lncRNA-6617的表達也可能受到上游mA去甲基化酶的調控;這些結果說明,mA修飾可能介導lncRNA-6617參與肌內脂肪細胞生成的調控,為后續深入研究豬肌內前體脂肪細胞生成的分子機制提供了理論基礎。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
